
恩施天然红椿种群结构及空间分布格局
2016-04-28 03:42冷艳芝苏长江宋丛文程德华操英南付翠林湖北生态工程职业技术学院湖北武汉4000湖北省七姊妹山国家级自然保护区管理局湖北恩施445500湘潭市第一中学湖南湘潭400
浙江农林大学学报 2016年1期
汪 洋,冷艳芝,苏长江,宋丛文,程德华,操英南,张 敏,付翠林(.湖北生态工程职业技术学院,湖北武汉4000;.湖北省七姊妹山国家级自然保护区管理局,湖北恩施445500;.湘潭市第一中学,湖南湘潭400)
恩施天然红椿种群结构及空间分布格局
汪洋1,冷艳芝1,苏长江2,宋丛文1,程德华3,操英南1,张敏1,付翠林1
(1.湖北生态工程职业技术学院,湖北武汉430200;2.湖北省七姊妹山国家级自然保护区管理局,湖北恩施445500;3.湘潭市第一中学,湖南湘潭411100)
摘要:红椿Toona ciliata是国家Ⅱ级重点保护野生植物,以红椿为优势种的天然种群十分少见。对湖北省恩施州4个不同红椿天然种群设立样方调查。编制了红椿特定时间生命表,绘制存活曲线,对种群龄级数量进行动态预测,并对空间分布格局进行分析,以研究不同种群空间结构和分布格局的成因。结果显示:4个种群的人为干扰强度:T4>T1>T2>T3;4个种群结构动态指数V(pi)分别为23.8%,34.0%,27.8%和32.3%,均为增长型。由于不同外界干扰强度,不同种群死亡率(qx)出现在龄级上推迟的现象;最高进入x龄级个体的生命期望(ex)值T3>T4>T1>T2。存活曲线不符合Deevey曲线,模拟符合三次函数曲线。25,50和100 m2取样面积上,方差/均值比率法的t检验、Morisita指数I(δ)参数的F检验、负二项参数K值均表明:T1和T4种群为聚集分布,T2和T3种群为泊松分布。聚集强度T4>T1>T2>T3;拥挤程度:T1>T4>T3>T2。研究表明:红椿种群在自然状态下处于增长状态,潜在干扰影响时,仍表现为稳定种群。红椿天然种群以泊松分布为特征,聚集分布主要来源于人为干扰。适当人为干扰,可以促进红椿种群更新、物种保护和生态效益最大化。图2表5参25
关键词:森林生态学;红椿;种群;空间结构;分布格局;天然种群
种群是生态学各个层次中最重要的一个层次,其中种群年龄结构和空间分布是其核心研究内容[1-2]。种群年龄结构反映了种群在特定时间世代重叠的特征,是分析探索种群动态的有效方法,被广泛应用于种群动态研究中[3]。植物空间分布格局的研究对于确定种群特征、种群间相互关系以及种群与环境之间的关系具有非常重要的作用,是植物群落空间结构的基本组成要素[4]。通过对森林中优势种群的结构和空间分布格局的研究,对阐明森林生态系统的形成与维持、群落的稳定性与演替规律、种群的生态特征和更新具有极为重要的意义[2,5]。红椿Toona ciliata,又名红楝子,楝科Meliaceae香椿属Toona,落叶或半常绿大乔木,强阳性树种,是中国热带、亚热带地区的珍贵速生用材树种。国家Ⅱ级重点保护野生植物。分布地理坐标为24°2l′~32°42′N,100°16′~119°40′E[6-7]。红椿天然林多呈零星分布,以红椿为优势种的天然种群十分少见。过度开发以及天然更新较慢,其数量不断减少[6]。鉴于红椿的优良品种特性和濒危的现实,越来越多的国内外学者对其进行广泛的研究,包含种质资源、遗传结构、植物生理、育种、栽培、造林、医药和化学成分、资源保护等方面,而生态学相关研究极少见报道。本研究从种群径级结构动态、空间分布格局等方面对红椿种群的基本特征进行了研究分析,旨在为红椿种群生态学研究和红椿种群保护与发展提供理论依据和科学参考。
1 研究区概况
4个研究样地(T1,T2,T3,T4)位于湖北省恩施州。样地T1和T2分别位于咸丰县黄金洞乡横石梁和村木田,星斗山国家级自然保护区最南端。样地T3位于恩施市盛家坝乡的马鹿河,属星斗山保护区的东部边沿。样地T4位于宣恩县七姊妹山国家级保护区的肖家湾。4个样地内红椿均为优势种。
T1与T2样地属亚热带大陆性季风气候。该区年平均气温为14.9℃,年平均降水1 400.0 mm以上,土壤为泥质页岩发育成的以扁砂土为主的质地疏松的山地黄壤,水热资源丰富,群山连绵起伏,植被生长繁茂[8-9]。样地T1主要乔木包含重阳木Bischofia polycarpa,枫杨Pterocarya stenoptera,丝栗栲Castanopsis fargesii等13种;灌木层及藤本主要有苎麻Urtica fissa,葎叶蛇葡萄Ampelopsis humulifolia,蔓赤车Pellionia scabra等13种;主要草本层为山麦冬Liriope spicata,过路黄Lysimachia christinae和天名精Carpesium abrotanoides等43种。
样地T2主要乔木有湖北木姜子Litsea hupehana,红麸杨Rhus punjabensis,栓叶安息香(红皮树)Styrax suberifolius等13个种;主要灌木有南一笼鸡Paragutzlaffia henryi,棠叶悬钩子Rubus malifolius和臭牡丹Clerodendrum bungei等16种;草本主要有楼梯草Elatostema umbellatum,南一笼鸡Paragutzlaffia henryi,疏叶卷柏Selaginella remotifolia和金荞麦Fagopyrum dibotrys等22种。
样地T3山地气候明显。该区年平均气温14.9℃,年降水量达1 300.0 mm以上[8-9]。5-9月间的降水量占全年的71.0%,年内季节分配不均匀。土壤为质地疏松的山地黄壤[8],土壤自然含水率较高,有机质、全氮、有效氮、全磷、速效钾相对较高。乔木丰富度最高(S=33)。群落乔木主要有绒毛钓樟Lindera floribunda,黑壳楠Lindera megaphylla,竹叶楠Arundina graminifolia等。灌木有蒙桑Morus mongolica,棠叶悬钩子Rubus malifolius,乌泡子Rubus parkeri等27种。草本主要有楼梯草Elatostema involucratum,赤爮Thladiantha dubia,高梁泡Rubus lambertianus等38种。
样地T4属于海拔800~1 200 m的亚高山地带,阳光充足,年平均气温为13.7℃,年降水量1 635.3 mm[10]。红椿群落位于山坡,土壤为山地黄棕壤。由于人为干扰严重,物种丰富度最低。主要乔木包含棕榈Trachycarpus fortunei和樱桃Cerasus pseudocerasus等。灌木藤本有八角枫Alangium platanifolium,柔毛绣球Hydrangea villosa,中国旌节花Stachyurus chinensis等11种。草本蕨类丰富,有大叶水龙骨Goniophlebium niponicum,卵心叶虎耳草Saxifraga aculeate和管茎凤仙花Impatiens tubulosa等36种。
2 研究方法
2.1样地选择与设置
香椿属植物都有种群规模小的特点。经过2 a的调查,确定了4处较为完整、有代表性的种群研究样地。按不同生境和现存规模,取样以覆盖全部红椿存活植株为标准。样地设置分别为:T1,15 m× 30 m;T2,20 m×20 m;T3,20 m×20 m;T4,10 m×40 m。4个样地总面积共1 650 m2。在各样地内采用5 m×5 m格子设置样方,共66个。对样方内红椿进行每木调查:对胸径≤2.5 cm的植株,测定地径、高度;对大于胸径≥2.5 cm的活立木,记录胸径、冠幅、枝下高等指标。样方内所有植物种类进行调查并统计记录。同时记录海拔高度、坡度等生境指标(表1)。样地人为干扰强度大小:T4>T1>T2>T3。
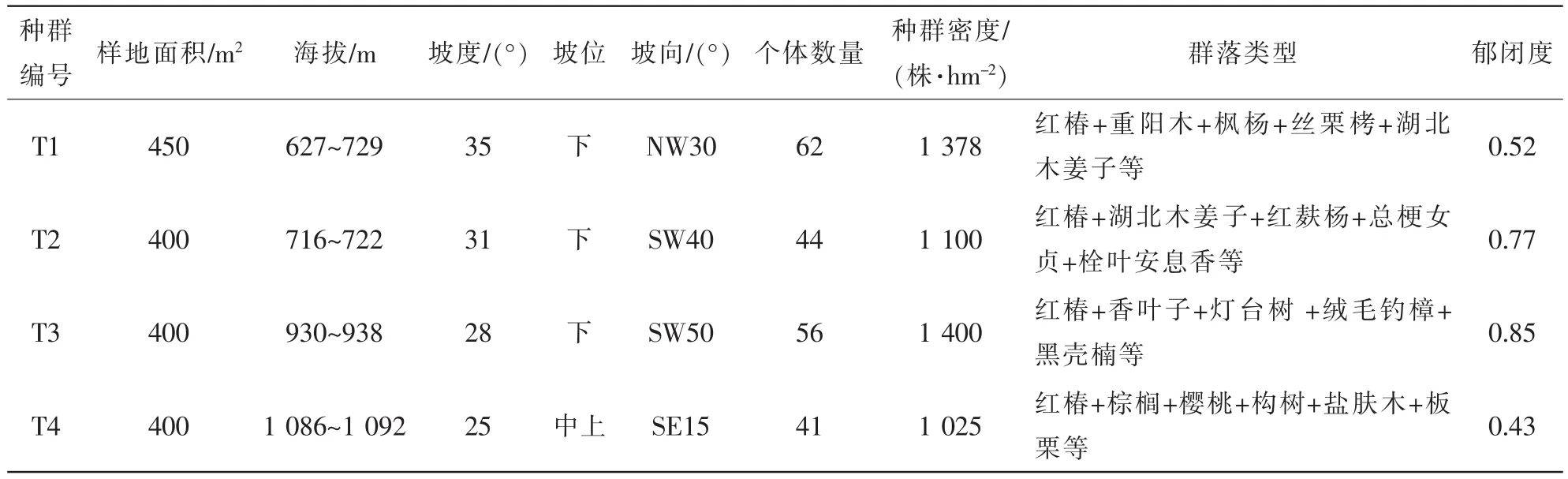
表1 不同红椿群落样地的环境特征Table 1 Environmental features of 4 Toona ciliata populations
2.2种群结构特征分析
2.2.1径级的划分龙汉利等[11]对红椿胸径与年龄的相关性进行拟合,得出:D=2.036 6 ln A0.784 1,相关系数R2=0.759 5,说明年龄(A)与胸径(D)之间相关性较高。本研究参照戴其生等[12]红椿用材林的解析数据,通过红椿平均胸径与连年生长过程分析,采用“空间替代时间法”,即将林木依胸径大小分级,以立木级结构代替种群年龄结构分析种群动态[13-14],将该种群划分为8个径级(平均4 a对应1个径级),从第2径级开始,以胸径5 cm为步长增加1级,即幼苗级Ⅰ(DBH<2.5 cm),幼树级Ⅱ(2.5 cm≤DBH<7.5 cm),Ⅲ(7.5 cm≤DBH<12.5 cm),Ⅳ(12.5 cm≤DBH<17.5 cm);中树级Ⅴ(17.5 cm≤DBH<22.5 cm),Ⅵ(22.5 cm≤DBH<27.5 cm);大树级Ⅶ(27.5 cm≤DBH<32.5 cm),Ⅷ(DBH≥32.5 cm)。然后将第Ⅰ径级对应第I龄级,第Ⅱ径级对应第Ⅱ龄级。如此类推,按龄级标准统计各级的植株数量。
2.2.2结构特征及动态以龄级为横坐标,个体数为纵坐标,分别绘制红椿种群龄级结构图。采用陈晓德的数学模型,推导种群年龄结构的动态指数,对红椿种群动态(V)进行定量描述[15-16],Vn,Vpi,V′pi取正、负、零值的意义分别反映种群或相邻年龄级个体数量的增长、衰退、稳定的动态关系。
2.2.3静态生命表及其存活曲线编制红椿种群静态生命表[13,17]。特定时间内生命表包含:x为单位时间内龄级;ax为在x龄级内红椿现有个体数;lx为在x龄级开始时标准化存活个体数(一般转换为1 000);dx为从x到x+1龄级间隔期内标准化死亡个体数;qx为从x到x+1龄级间隔期间死亡率;Lx为从x到x+1龄级间隔期间还存活的个体数;Tx为从x龄级到超过x龄级的个体总数;ex为进入x龄级个体的生命期望或平均期望寿命;Kx为亏损率(损失度)。以上各项相互关联,通过实测值ax或dx求得,其关系为:lx=(ax/a0)×1 000;dx= lx+lx+1;dx= lx-lx+1;qx=(dx/ lx)×100%;Lx=(lx+ lx+1)/2;Tx=Lx;ex=Tx/lx。以龄级为横坐标,以生命表中标准化存活数的自然对数ln(lx)为纵坐标作图,绘制4个红椿种群存活曲线。
2.2.4种群空间分布格局分析为准确研究红椿种群分布格局,设定不同格子大小进行研究:5 m×5 m,5 m×10 m,10 m×10 m。采用多个分布格局数学模型进行测度,以避免不同模型的片面性,即扩散系数(C)的t检验和负二项参数(K),Cassie指标(Ca),Lloyd平均拥挤度(m*),聚块性指数IPA(m*/m),David &Moore的丛生指标(I),Morisita扩散型指数(Iδ)的F检验,进行分布格局与聚集强度的判定[17-18]。
3 结果与分析
3.1年龄结构及动态
从红椿龄级结构图1可以看出,4个不同样地的红椿种群均属于增长型种群。种群均在第Ⅰ龄级占有最大百分比,分别达到30.65%,31.82%,28.57%和36.59%。种群T1存活数量在第Ⅴ龄级最低,占4.84%;种群T2的存活数最低在第Ⅶ龄级和第Ⅷ龄级,均为4.55%;种群T3最低存活数在第Ⅳ龄级,为3.57%;种群T4的最低值在第Ⅴ龄级,只有2.44%。4个样地同样经历了较高数量的幼苗存活期和损失期,中龄级植株的高损失期,高龄级时的种群稳定期。

图1 不同种群红椿年龄结构图Figure 1 Age structure of Toona ciliata populations
种群相邻级间个体数量的变化动态结果(表2)显示,4个不同种群动态指数变化差异较大,但增长性较强。4个种群在不同龄级间均出现2~3次负增长动态,但总体变化指数Vpi均大于0,V′pi指数也大于0,说明4个种群均为增长型种群。4个种群动态增长指数为T2>T4>T3>T1,说明不同种群增长能力的大小;干扰条件下的动态指数为T4>T2>T3>T1;干扰敏感指数为T4>T2=T3>T1,表明了种群对应外界风险能力指数的大小。实地调查发现,4样地红椿结实情况良好。丰富的生殖个体维系了种群整体的相对稳定。
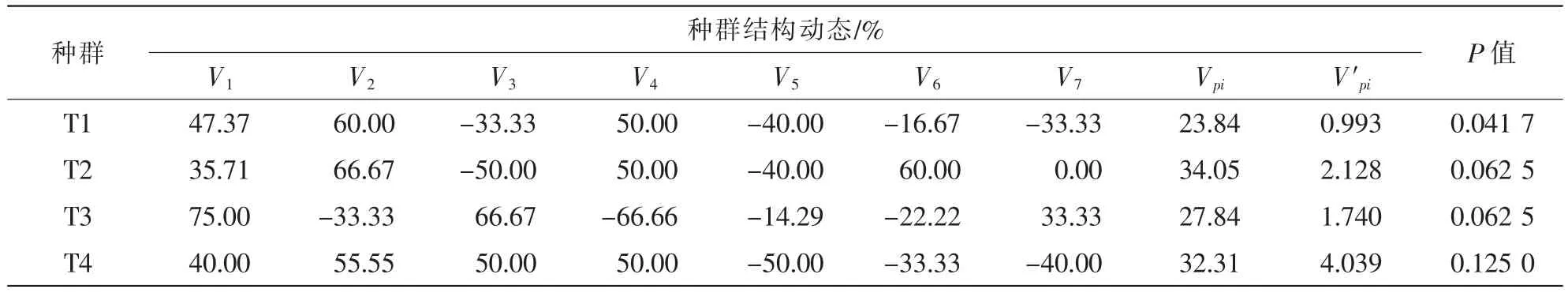
表2 不同生境红椿种群结构动态分析Table 2 Dynamic analysis of population structure of Toona ciliata in different habitats
3.2种群生命表分析
根据径级年龄结构分级数据,编制出红椿种群的特定时间生命表。表3显示:4个不同年龄结构的种群都表现出幼苗期植株数量最高,中间龄级数量最少或波动,高龄级种群趋于稳态。种群T1幼苗最多,高龄级存活数量也最多。实际调查中,T1种群位于自然村落旁,人为干扰因素促进了种群的结构变化。T1种群人为干扰仅低于T4,其qx最高出现在第Ⅴ龄级,低峰在第Ⅵ龄级;T2种群也存在一定人为干扰,qx在第Ⅱ龄级和第Ⅴ龄级;低峰在第Ⅶ龄级,为0;T3种群人为干扰最小,能代表红椿天然种群的典型特征,qx出现2个高点,分别在第Ⅰ龄级和第Ⅳ龄级,说明了幼苗和幼树2个阶段的环境筛选。种群T4的人为干扰最强,在第Ⅴ龄级时,qx=1.000,其种群结构与T1种群存在相似性。期望寿命ex可反映各龄级内个体的平均生存能力[19-20]。ex值高表明生理活动旺盛,生命力最强,生存质量高。ex值的大小分别为:T3>T4>T1>T2。T1,T2与T4表现出了同样的qx龄级对应现象,明显与T3不同。显然这与3个种群受到的不同程度强干扰有关。干扰改变了第Ⅱ龄级的环境筛选结构,使qx向后龄级推迟。
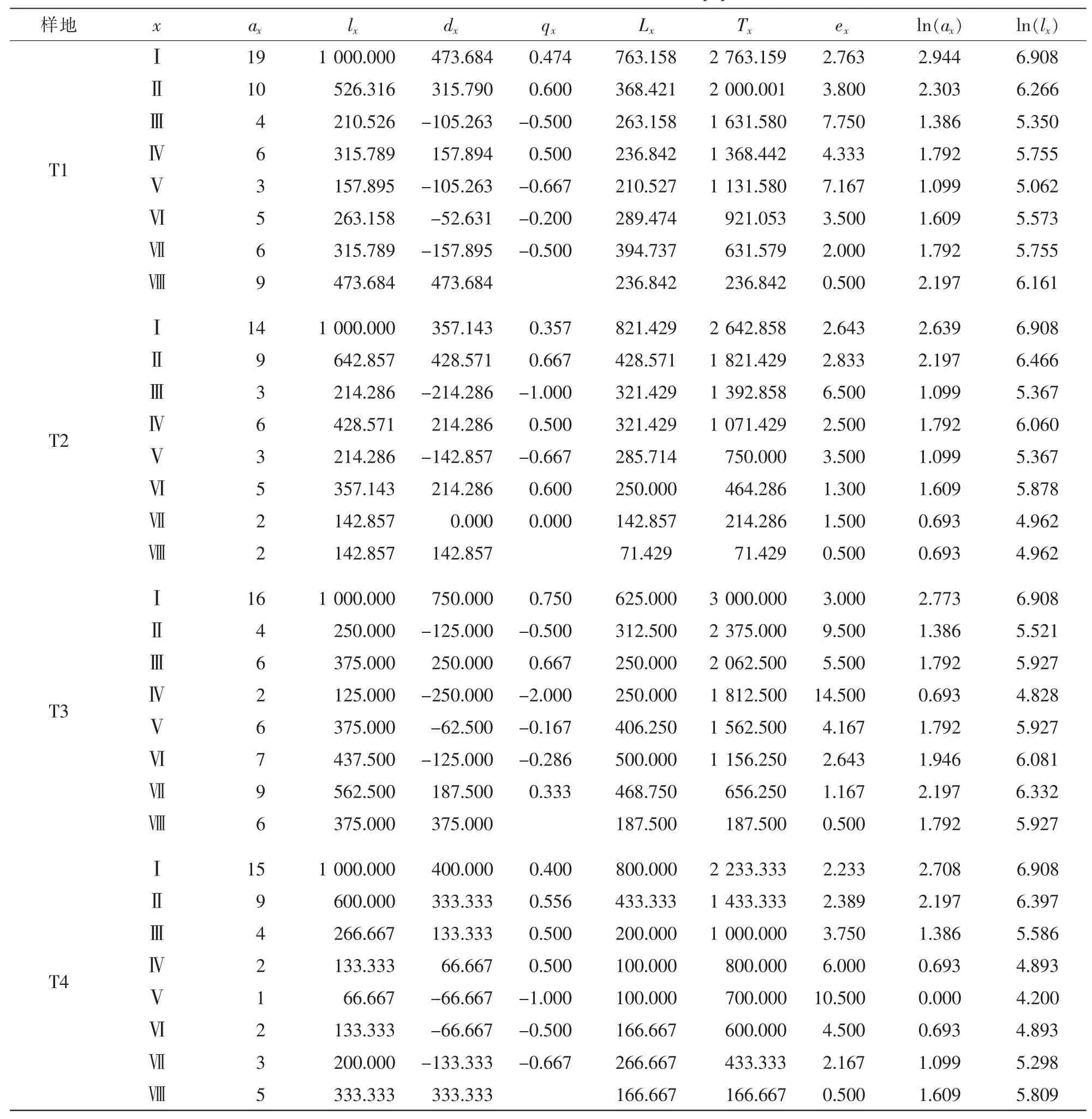
表3 不同样地红椿种群静态生命表Table 3 Static life table in different Toona ciliata populations
按照DEEVEY的划分,种群存活曲线一般有3种基本类型:I型是凸型曲线,Ⅱ型是直线,Ⅲ型是凹型曲线[21-22],分别表示不同的动态意义。存活曲线图2显示4个种群的存活曲线均不符合Deevey型曲线特征。红椿天然种群的特殊生理特征,生活环境和不同干扰因子,使得存活数量在不同龄级出现波动。通过SPSS软件用不同的回归方程进行存活曲线的拟合(表4)。
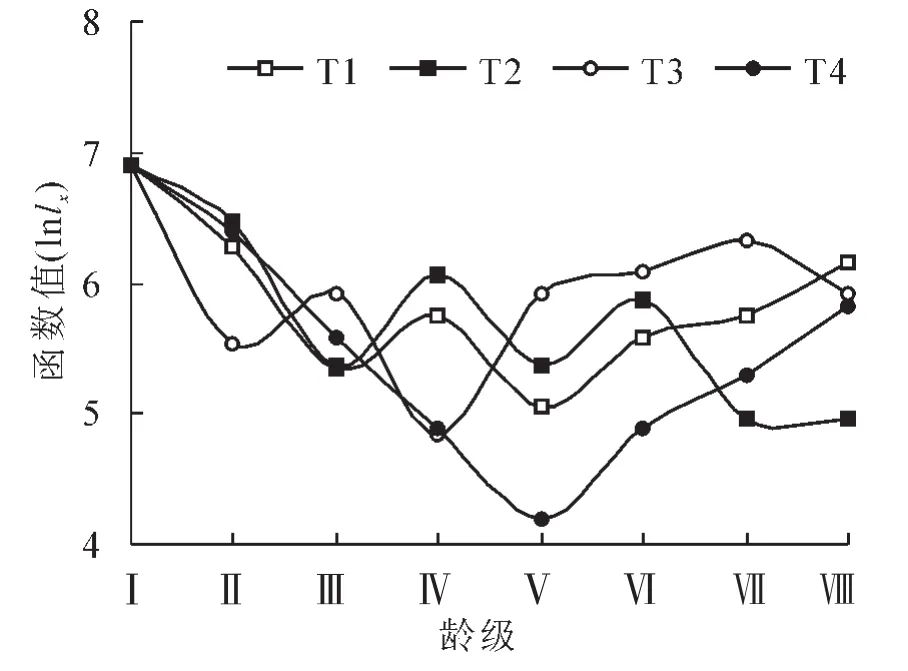
图2 不同种群红椿种群存活曲线Figure 2 Sruvival curves of different Toona ciliata populations
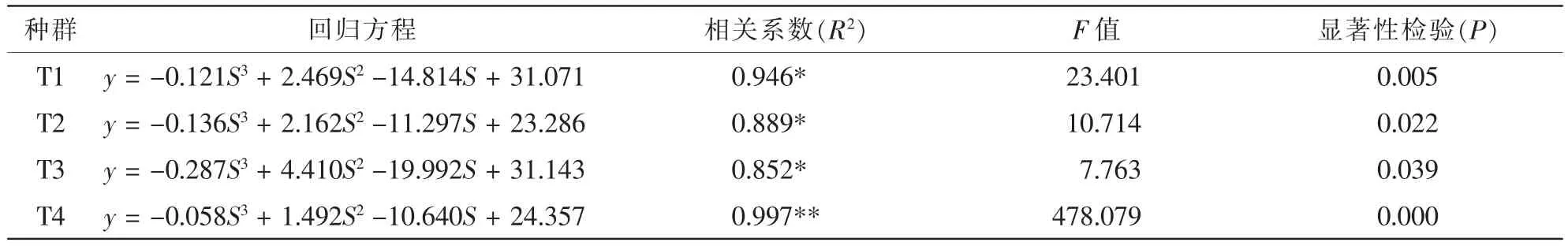
表4 不同红椿种群存活曲线回归方程拟合Table 4 Cubic curvilinear equation fitting for survival curves of 4 Toona ciliata populations
表4表明:T1,T2和T3等3个种群的显著性检验表现为极显著,T4种群为显著。4个不同种群存活曲线都符合表4模型。T3种群相关系数最小:R2>0.852,最小F值大于7.763。因此,三次曲线回归方程比较适合4个红椿种群的发展趋势。通过模型的建立,可以预测出不同径级的个体存活数量趋势。4个存活曲线反应出T1,T2和T4的曲线特征与T3不同。
3.3空间格局分析
种群在一定的空间内都会呈现出特有的分布格局,它能够反映出种内种间关系、环境适应机制、空间异质性等[2]。表5中,T1种群无法在100 m2取样。在25和50 m2取样时,对应自由度为17和8;T2,T3和T4种群在25,50,100 m2尺度对应自由度分别为15,7和3。由表5可见:4个红椿种群不同取样大小,扩散系数法的t检验结果表明:T1种群在25和50 m2尺度时为聚集分布;T2种群在25,50和100 m2为泊松分布;T3种群25和50 m2尺度下为泊松分布,100 m2尺度时趋近均匀分布;T4种群在25,50和100 m2尺度时为聚集分布。聚集程度负二项参数K值愈小,聚集程度越高,当K值趋于无穷大时(一般为8以上),则逼近泊松分布[23]。K>0时,为聚集分布,T2和T3种群K大于8,为泊松分布。T1和T4种群小于8,为集群分布。对F0的检验,F0≤F0.05为随机分布,F0>F0.05为集聚分布。Morisita指数Iδ结果经F0检验,与扩散系数t检验完全一致。
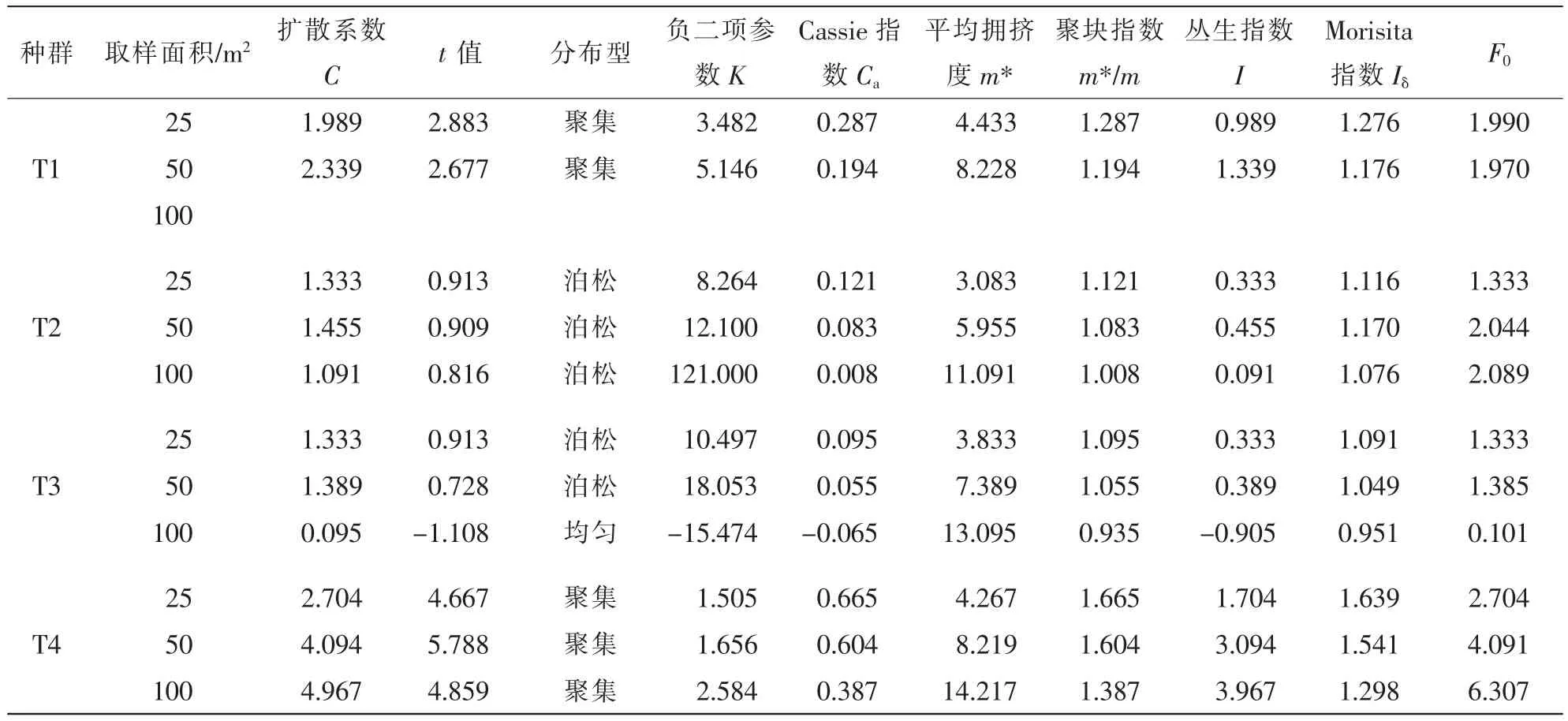
表5 不同取样尺度红椿种群分布格局Table 5 Distribution pattern of Toona ciliata populations in different sample sizes
聚集强度T4>T1>T2>T3。平均拥挤指数m*表示生物个体在1个样方中的平均邻居数,反映了样方内生物个体的拥挤程度,数值越大聚集强度越大,表示种群内个体受其他个体的拥挤效应越大[24]。4个不同种群的拥挤程度分别表明:T1>T4>T3>T2。聚集强度与拥挤程度进一步说明了T1和T4种群的聚集分布特性及T2和T3种群的泊松分布特性。
4 讨论与结论
红椿幼苗有一定的耐阴性,幼苗到幼树阶段,需要足够的光照条件。强阳性特征,决定了红椿种群必须占据群落上层才能进一步发展。群落上层环境复杂,其他乔木、大型藤本一旦进入群落上层,较差的光照条件必然影响红椿幼树的生长,产生高死亡率,导致种群在第Ⅲ龄级到Ⅴ龄级的数量稀少,种群出现负增长。4个不同天然种群动态结构分析说明:红椿天然种群基本处于增长型向稳定型过渡阶段。
种群的龄级与存活数量曲线表明:T2和T3种群出现2次波动现象,可能与种群的2次“自疏”现象有关。波动在幼苗到幼树期间最为明显,此间种内种间竞争激烈。而T1和T4种群成活曲线显示第Ⅴ级的高死亡率qx和高期望寿命ex,则可能缘于“自疏”和外界干扰的共同作用。
种群空间分布格局是生物群落中各种内外因素相互作用最直接的反应结果。这不仅与物种的生物学特性和种群间的竞争排斥有关,而且与物种的生境(包括土壤、地形、地貌等)有密切的联系[25]。植物种群分布格局并不是一成不变的,而是随时间变化表现出动态变化的过程[25]。
恩施天然红椿种群的空间格局主要符合聚集分布和随机分布。这2种分布格局表现出与外部干扰有相关效应。4个种群的人为干扰强度为:T4>T1>T2>T3。T1和T4为集聚分布,T2和T3为泊松分布。T1种群位于自然村落旁,种群处于村民住宅和河流之间的狭长落叶阔叶林群落之间,群落结构较完整。虽然人为干扰较小,但种群的发展区域受限,且红椿群落被几条小路分为多段。T4种群位于七姊妹山保护区的缓冲区,红椿群落一面依山,另一面被开垦的农田隔断,形成沿山凹地段延展的受干扰群落。T2种群干扰较小,种群处于恢复期。T3种群人为干扰最小,能代表红椿天然种群的特征。
可以得出结论:红椿天然种群以泊松分布为特征,聚集分布主要来源于人为干扰。另一方面,种群聚集分布或在某一斑块上形成优势有利于增加存活机会、抵抗外来种的侵入和定居,发挥群体效应,从而维持种群的稳定和续存[25]。根据红椿种群结构和分布格局特性,采用适当人为干扰,采伐成熟林,制造林窗,加强林隙和林窗的幼苗抚育等管理措施,可以促进幼苗向幼树转化和种群更新、生长与保护,提高林木质量,使生态效益最大化。
5 参考文献
[1]李博,杨持,林鹏.生态学[M].北京:高等教育出版社,2000.
[2]张育新,马克明,祁建,等.北京东灵山海拔梯度上辽东栎种群结构和空间分布[J].生态学报,2009,29(6):2789-2796.ZHANG Yuxin,MA Keming,QI Jian,et al.Size structure and spatial pattern of Quercus liaotungensis population along elevation gradient in Dongling Mountain,Beijing[J].Acta Ecol Sin,2009,29(6):2789-2796.
[3]胡尔查,王晓江,刘永洪,等.乌拉山自然保护区油松种群结构与分布格局研究[J].林业科学研究,2011,24(3):363-369.HU ercha,WANG Xiaojiang,LIU Yonghong,et al.Population structure and distribution pattern of Pinus tabulaeformis in Wulashan Nature Reserve of Inner Mongolia[J].For Res,2011,24(3):363-369.
[4]周国英,陈桂琛,魏国良,等.青海湖地区芨芨草群落主要种群分布格局研究[J].西北植物学报,2006,26(3):579-584.ZHOU Guoying,CHEN Guichen,WEI Guoliang.et al.Distribution patterns of major populations in Achnatherum splendens communities of Qinghai Lake area[J].Acta Bot Boreali-Occident Sin,2006,26(3):579-584.
[5]徐学红,于明坚,胡正华,等.浙江古田山自然保护区甜槠种群结构与动态[J].生态学报,2005,25(3):645 -653.XU Xuehong,YU mingjian,HU Zhenghua,et al.The structure and dynamics of Castanopsis eyrei population in Gutian Mountain Natural Reserve in Zhejiang,East China[J].Acta Ecol Sin,2005,25(3):645-653.
[6]吴际友,程勇,王旭军,等.红椿无性系嫩枝扦插繁殖试验[J].湖南林业科技,2011,38(4):5-8.WU Jiyou,CHENG Yong,WANG Xujun,et al.Softwood cutting propagation of Toona ciliata clones[J].J Hunan For Sci Technol,2011,38(4):5-8.
[7]汪洋,冉勇军,冷艳芝,等.红椿和毛红椿遗传与生理生态相关研究[J].安徽农业科学,2014,42(20):6910 -6912.WANG Yang,RAN Yongjun,LENG Yanzhi,et al.Research progress of rare commercial tree species Toona ciliata Roem[J].J Anhui Agric Sci,2014,42(20):6910-6912.
[8]陈绍林,张志华,廖于实,等.星斗山自然保护区秃杉原生种群生境现状及保护对策[J].安徽农业科学,2008,36(11):4624-4625.CHEN Shaolin,ZHANG Zhihua,LIAO Yushi,et al.Current status and protection countermeasures of Taiwania flousiana Gaussen habitat in Xingdoushan Nature Reserve[J].J Anhui Agric Sci,2008,36(11):4624-4625.
[9]刘毅,孙云逸,陈世明,等.湖北星斗山国家级自然保护区生物多样性分析与保护[J].湖北林业科技,2007(6):42-45.LIU Yi,SUN yunyi,CHEN Shiming,et al.Analysis and protection on biological diversity in Xingdoushan State-class Reserve of Hubei[J].J Hubei For Sci Technol,2007(6):42-45.
[10]方元平,刘胜祥,汪正祥,等.七姊妹山自然保护区野生保护植物优先保护定量研究[J].西北植物学报,2007,27(2):348-355.FANG Yuanping,LIU Shengxiang,WANG Zhengxiang,et al.Quantitative assessment of priority for conservation of the national protected plants in Qizimeishan Mountain Nature Preserve[J].Acta Bot Boreali-Occident Sin,2007,27(2):0348-0355.
[11]龙汉利,冯毅,向青,等.四川盆周山地红椿生长特性研究[J].四川林业科技,2011,32(3):37-41.LONG Hanli,FENG Yi,XIANG Qing,et al.Study of the growth characteristics of Toona ciliata trees in mountainous areas around the Sichuan Basin[J].J Sichuan For Sci Technol,2011,32(3):37-41.
[12]戴其生,张梅林,徐玉伟,等.红楝子人工造林试验初报[J].安徽林业科技,1997,34(1):33-34.DAI Qisheng,ZHANG Meilin,XU Yuwei,et al.Plantation test of Toona ciliata Roem[J].J Anhui For Sci Technol,1997,34(1):33-34.
[13]江洪.云杉种群生态学[M].北京:中国林业出版社,1992:8-26.
[14]吕海英,王孝安,李进,等.珍稀植物银砂槐中国分布区的种群结构与动态分析[J].西北植物学报,2014,34(1):177-183.LÜ Haiying,WANG Xiao’an,LI Jin,et al.Structure and dynamics of China rare plant Ammodendron bifolium(Pall.)Yakoal natural population[J].Acta Bot Boreali-Occident Sin,2014,34(1):177-183.
[15]陈晓德.植物种群与群落结构动态量化分析方法研究[J].生态学报,1998,18(2):214-217.CHEN Xiaode.A study on the method of quantitative analysis for plant population and community structural dynamics[J].Acta Ecol Sin,1998,18(2):214-217.
[16]LEAK W B.Age distribution in virgin red spruce and northern hard woods[J].Ecology,1975,56(6):1451-1454.
[17]洪伟,王新功,吴承祯,等.濒危植物南方红豆杉种群生命表及谱分析[J].应用生态学报,2004,15(6):1109-1112.HONG Wei,WANG Xingong,WU Chengzhen,et al.Life table and spectral analysis of endangered plant Taxus chinensis var.mairei population[J].Chin J Appl Ecol,2004,15(6):1109-1112.
[18]兰国玉,雷瑞德.植物种群空间分布格局研究方法概述[J].西北林学院学报,2003,18(2):17-21.LAN Guoyu,LEI Ruide.Brief introduction of spatial methods to distribution patterns of population[J].J Northwest For Coll,2003,18(2):17-21.
[19]刘任涛,毕润成,闫桂琴,等.山西稀有濒危植物山核桃种群动态与谱分析[J].武汉植物学研究,2007,25(3):255-260.LIU Rentao,BI Runcheng,YAN Guiqin,et al.Population dynamics and spectral analysis of rare-endangered plant Juglans mandshurica in Shanxi Province[J].J Wuhan Bot Res,2007,25(3):255-260.
[20]申仕康,马海英,王跃华,等.濒危植物猪血木(Euryodendron excelsum H.T.Chang)自然种群结构及动态[J].生态学报,2008,28(5):2404-2412.SHEN Shikang,MA Haiying,WANG Yuehua,et al.The structure and dynamics of natural population of the endan-gered plant Euryodendron excebum H.T.Chang[J].Acta Ecol Sin,2008,28(5):2404-2412.
[21]万志兵,程国源,丁强强,等.绩溪光皮桦次生林的种群动态[J].东北林业大学学报,2014,42(1):27-30.WAN Zhibing,CHENG Guoyuan,DING Qiangqiang,et al.Population structure dynamics of Betula luminifera secondary forest in Jixi County of Anhui Province,China[J].J Northeast For Univ,2014,42(1):27-30.
[22]赵志刚,郭俊杰,曾杰,等.广西大明山格木种群的空间分布格局与数量动态特征[J].林业科学,2014,50(10):1-7.ZHAO Zhigang,GUO Junjie,ZENG Jie,et al.Spatial patterns and quantitative dynamics of Erythrophleum fordii populations at Damingshan Mountain,Guangxi[J].Sci Silv Sin,2014,50(10):1-7.
[23]徐坤,谢应忠,李世忠.宁南黄土丘陵区退化草地群落主要植物种群空间分布格局对比研究[J].西北农业学报,2006,15(5):123-l27.XU Kun,XIE Yingzhong,LI Shizhong.A community on spatial patterns of prime plant population of degenerative steppe in loess hilly region of Northern Ningxia[J].Acta Agric Boreali-Occident Sin,2006,15(5):123-l27.
[24]王新功,洪伟,吴承祯,等.武夷山米储林群落优势种群空间格局的研究[J].河南农业大学学报,2003,37(1):44-48.WANG Xingong,HONG Wei,WU Chengzhen,et al.Study on the distribution pattern of dominant population Castanopsis carlessii forest in Wuyishan Mountains[J].J Henan Agric Univ,2003,37(1):44-48.
[25]韩路,王海珍,彭杰,等.塔里木河上游天然胡杨林种群空间分布格局与动态研究[J].西北植物学报,2007,27(8):1668-1673.HAN Lu,WANG Haizhen,PENG Jie,et al.Spatial distribution patterns and dynamics of major population in Populus euphratica forest in upper reaches of Tarim River[J].Acta Bot Boreali-Occident Sin,2007,27(8):1668-1673.
Spatial structure and distribution pattern of natural Toona ciliata populations in the Enshi Region
WANG Yang1,LENG Yanzhi1,SU Changjiang2,SONG Congwen1,CHENG Dehua3,CAO Yingnan1,ZHANG Min1,FU Cuilin1
(1.Hubei Vocational College of Ecological Engineering,Wuhan 430200,Hubei,China;2.Management Office,Qizimeishan National Nature Reserve,Enshi 445500,Hubei,China;3.Xiangtan No.1 High School,Xiangtan 411100,Hunan,China)
Abstract:Toona ciliata is a key protected tree species at the second national level,its natural population acts as a dominant species in its existing community,is rarely seen in subtropical regions of China.This study was to research the structure of age class,the quantity dynamic conditions,the trend of development and the correlation between the distribution pattern and the structure of age class,the quantity dynamic condition of different Toona ciliata populations.Four sample plots were selected in the Enshi Region of Hubei.The sample plots,entirely covering 4 populations,were set up,each in 5 m×5 m grid,by using contiguous grid quadrate method,with T1 10 m×30 m,T2 20 m×20 m,T3 20 m×20 m and T4 10 m×40 m in size respectively.Based on field survey data,a specific life table was established,survival curve was determined,a dynamic estimation of the survival number for different age classes was made,and spatial distribution patterns of different Toona ciliata populations were analyzed to determine the causes of spatial structures and distribution patterns in different sampling plots.Analysis included Deevey curves,regression,ratio method of variance &mean valuebook=18,ebook=21with t test,Morisita indices(I(δ))with F test and negative binomial parameter(K),patchiness index(m*/m),and mean crowding index(m*).Results of the human disturbance intensity for the four populations was T4>T1>T2>T3 with dynamic indices(V(pi))of T1 = 23.8%,T2 = 34.1%,T3 = 27.8%,and T4 = 32.3% showing growth form.Also,the qxvalues of different populations lagged in age-class with external disturbance intensities,and the life expectancy peak values(ex)varied as follows:T3>T4>T1>T2.Survival curves did not agree with any Deevey curves,but 4 cubic functions represented a regression of the four survival curves:y=-0.121S3+2.469S2-14.814S+31.071,y=-0.136S3+2.162S2-11.297S + 23.286,y=-0.287S3+4.410S2-19.992S +31.143,y=-0.058S3+1.492S2-10.64S + 24.357;With 25 m2,50 m2,and 100 m2as sample sizes,a ratio of variance and mean value to t test,I(δ)with F test,and K all demonstrated that T1 and T4 populations had clumped distributions,but T2 and T3 had Poisson distributions.The m*/m varied with a sequence of T4>T1>T2>T3;whereas,m* was T1>T4>T3>T2.This research demonstrated that in a natural state,the Toona ciliata population was in growth form with a stable population susceptible to potential external disturbances and having a Poisson distribution,but with human disturbance a clumped distribution emerged that with proper human objective regulations,population regeneration,species protection,and maximum ecological effect could be enhanced.[Ch,2 fig.5 tab.25 ref.]
Key words:forest ecology;Toona ciliata;population;spatial structure;distribution pattern;natural population
作者简介:汪洋,副教授,从事生态学研究。E-mail:13296698026@163.com。通信作者:宋丛文,教授,博士,从事植物遗传学研究。E-mail:songcongwen@126.com
项目基金:湖北省科技厅公益性研究项目(40 2012DBA40001)
收稿日期:2015-03-01;修回日期:2015-05-27
doi:10.11833/j.issn.2095-0756.2016.01.003
中图分类号:S718.5;S792.33
文献标志码:A
文章编号:2095-0756(2016)01-0017-09
猜你喜欢
海洋通报(2022年6期)2023-01-07
今日农业(2022年15期)2022-09-20
湖南电力(2021年1期)2021-04-13
新农业(2020年18期)2021-01-07
阿来研究(2020年1期)2020-10-28
——评《多中心城市空间结构:概念、案例与优化策略》
浙江工业大学学报(社会科学版)(2020年2期)2020-07-17
红土地(2018年7期)2018-09-26
生物学教学(2018年3期)2018-08-08
中学生物学(2018年8期)2018-03-01
环球人文地理·评论版(2016年8期)2017-01-19
