
早春类短命植物生物量研究(Ⅲ)——顶冰花生物量特征及模型构建
2016-04-27 06:17郑金萍范春楠李金玉
北华大学学报(自然科学版) 2016年2期
郑金萍,范春楠,程 岩,刘 彬,李金玉
(1.北华大学林学院,吉林 吉林 132013;2.吉林省松花江三湖自然保护区管理局,吉林 吉林 132013)
早春类短命植物生物量研究(Ⅲ)
——顶冰花生物量特征及模型构建
郑金萍1,范春楠1,程岩2,刘彬1,李金玉1
(1.北华大学林学院,吉林 吉林132013;2.吉林省松花江三湖自然保护区管理局,吉林 吉林132013)
摘要:以早春类短命植物顶冰花为研究对象,采用全收获法对其生物量进行统计.结果表明:顶冰花的单株生物量平均为0.316 2 g,地下部分较地上部分平均高出37.92%.随着叶长的增大,叶生物量、果生物量、总生物量、地上生物量与地下生物量均显著增加,而花生物量显著减小.有性生殖生长与植株个体大小的关系密切,占总生物量的5.47%~19.42%,平均为11.92%;占营养器官的比例平均为14.41%,表现出随叶长的增大呈先增加后下降再增加的变化趋势.根冠比介于0.93~1.86,平均为1.33,随叶长的增大呈单峰型变化.整体来看,顶冰花的生物量有随生长发育由地上生长转为地下生长,再由地下生长转为地上为主的变化和从营养生长逐渐转向有性生殖的趋势.以叶长为自变量拟合顶冰花地上、地下和全株的4种生物量估测模型,除地下生物量的指数模型外,拟合和验证结果均达到相关建模的精度要求.
关键词:早春类短命植物;顶冰花;生物量;模型构建
【引用格式】郑金萍,范春楠,程岩,等.早春类短命植物生物量研究(Ⅲ):顶冰花生物量特征及模型构建[J].北华大学学报(自然科学版),2016,17(2):165-170.
生物量不仅可以应用于分析植被的生产能力,还可为评价植被的能量利用率、养分循环及多途径利用等提供数据基础[1].近年来,生物量的研究已经与全球气候变化及碳循环等紧密联系[2-6].草本层生物量与乔木层相比,所占比例仅为0.03%~2.87%[7-8],因而在森林生态系统的研究中常被忽略;但事实上,草本植物层片却提供了大约4%的净初级生产量[9].早春类短命植物作为温带森林生态系统独特的草本植物类群[10],可提高森林生态系统的光能利用率和物质循环、能量流动速度,是森林生态系统早春时期生产者的重要组成部分;并具有抗寒能力强,对光照敏感,含水率高,分布广泛等特点.然而有关早春类短命植物的生物量状况、分配特点及其在森林生态系统中占有的地位等研究,还鲜见报道[11-13].
顶冰花(Gagealutea)为百合科(Liliaceae)顶冰花属(Gagea)多年生早春类短命草本植物,鳞茎卵球形,基生叶1枚,花3~5朵,排成伞形花序着生于花梗顶端,蒴果卵圆形至倒卵形,花期4~5月,果期6月;广泛分布于我国东北各省,生于林下、灌丛或草地,在日本、朝鲜、俄罗斯和欧洲也有分布[14-15].顶冰花的药用价值始载于《新华本草纲要》,性苦、味平,入心经,民间常用本品治疗心脏病;但据肖淑宁[16]的记载,顶冰花食用数株即可中毒,4 g以上即可致死;然而现代医药在其药理与药效等方面的研究还未见报道.本文采用全收获法测定顶冰花植株生物量特点,分析生物量的分配特征,并以叶长级为自变量构建生物量的预估模型.研究结果可为明确顶冰花的生长策略,估测其资源储量和在森林生态系统中的地位,以及资源开发与利用等提供重要的理论支撑.
1研究方法
本研究采样地点位于吉林市磨盘山的次生落叶阔叶林内,研究地位置、气候与植被状况等详见文献[17-18].取样方法:通过踏查选择顶冰花集中分布区,随机选取生长健康植株,测量后采用全收获法采集植株样本.由于顶冰花的基生叶仅1枚,下弯,自然生长株高测量困难,且难以表征其生长状况,因此本研究以叶长作为易测生长指标,并将采集样本按不同叶长级(以2 cm为步长)进行分装,标记后带回.顶冰花样本的室内处理以每个植株个体为单位,按不同器官(地下、叶、花、果)进行分拣、包装、编号后,置于干燥箱内,80 ℃烘干至恒重,称取其器官生物量.研究于2014年4月末至5月初进行,共计采集顶冰花植株样本154株.
顶冰花生物量模型的构建选取4种常用的一元线性方程(幂函数y=axb、指数y=aebx、直线y=ax+b和对数方程y=a+blnx),以叶长级为自变量,建立其地上生物量、地下生物量和全株生物量回归模型.其中,建模样本121株,预留33株样本用于验证所构建模型的精度(在每个叶长级内随机选取样本各3株),模型的具体验证方法详见文献[17-18].
2结果与分析
2.1生长状况及生物量分配
以2 cm叶长为步长,对154株顶冰花植株样本的生长状况及生物量分配进行统计,结果见图1和表1.
由图1可以看出:顶冰花的生长发育与叶长之间的关系密切,当叶长级为16 cm时即存在24%的开花个体,但随着叶长的增大开花个体比例整体下降,当叶长为30 cm以上时开花植株消失;处于营养生长阶段的植株比例也随叶长的增大而下降,16 cm叶长级为76%,在32 cm叶长级下降为0;结果植株个体的比例变化与二者相反,在18 cm叶长级开始出现,在34 cm叶长级处达到100%;花果并存个体比例整体随叶长级的变化呈单峰曲线,在22 cm叶长级时最高,达30%.经相关分析发现:营养生长植株和开花植株个体数量所占比例均与叶长呈极显著的负相关关系(P=-0.909和-0.830,Sig.=0.000和0.002);花果共存的植株个体比例也与叶长呈负相关,但相关关系并不显著;而结果植株个体数量所占比例则与叶长呈极显著的正相关关系(P=0.910,Sig.=0.000).随着叶长级的增大,顶冰花处于营养生长和开花阶段的植株个体显著减少,处于结果阶段的个体数量显著增多.
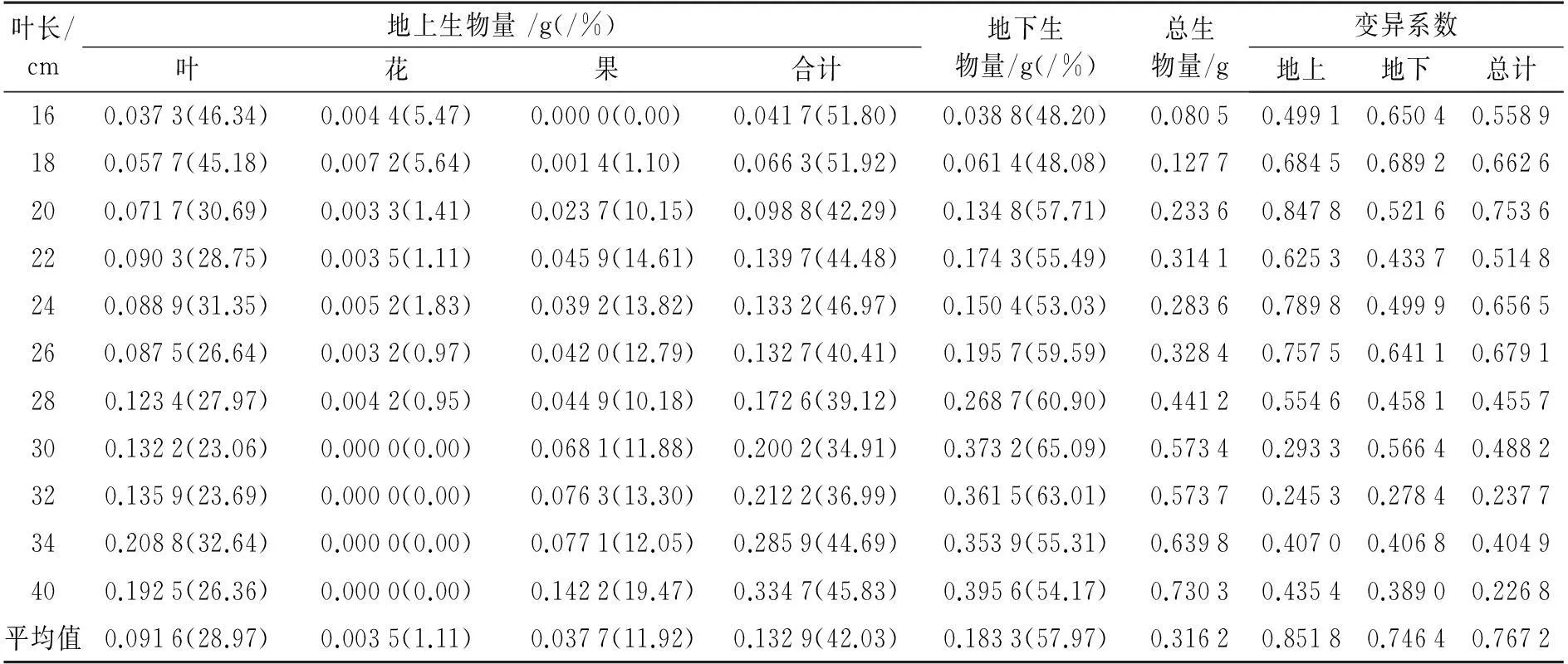
表1 顶冰花的生物量特征
注:括号内数值为各器官生物量占总生物量的比例
顶冰花平均单株生物量为0.316 2 g,地下生物量(0.183 3 g)略高于地上生物量(0.132 9 g),平均高出37.92%.不同叶长级植株的叶生物量、果生物量、总生物量、地上生物量与地下生物量均表现出随叶长的增加而增大,正相关关系均达到了极显著水平(P>0.9,Sig.<0.001),而花生物量则表现出随叶长的增加而显著减小(P=-0.817,Sig.=0.002);但受自身年龄、光照、土壤以及立地条件等因素影响其生物量在不同叶长级内表现出较大的变幅.其中,地下生物量在各叶长级内的变异系数介于24.53%~84.7%,以叶长级18~28 cm范围的变异相对较大,均在50%以上;地上生物量的变异较地下生物量的变异略小,介于27.84%~68.92%,整体随叶长的增加而减小,且负相关关系显著(P=-0.715,Sig.=0.013);总生物量的变异系数介于22.68%~75.36%,也表现出整体随叶长的增加而减小的趋势,且负相关达极显著水平(P=-0.821,Sig.=0.002).
不同叶长级的顶冰花变异系数总生物量、地下和地上部分分别为76.72%,85.18%和74.64%,均普遍高于同等叶长级的生物量变异.经方差分析得出:不同叶长级的顶冰花全株生物量(F=25.169,Sig.=0.000)、地上生物量(F=16.870,Sig.=0.000)和地下生物量(F=19.577,Sig.=0.000)均存在极显著差异.
顶冰花的地上生物量分配比例随叶长的增大呈先减小后增加的变化,地下生物量与之相反.根冠比介于0.93~1.86,平均为1.33,整体呈单峰型变化,在叶长级为30 cm时达到峰值,随后下降.有性生殖器官和营养器官生物量均随叶长的增加而增大,其线性关系均可用直线方程拟合,且相关系数r均在0.95以上;但随着叶长的增加花、果生物量比例的降低和增高变化明显,合计比例介于5.47%~19.42%,平均为11.92%;有性生殖器官占营养器官的比例平均为14.41%,随叶长的增加呈先增加后下降再增加的变化趋势,由叶长16 cm的5.78%增至22 cm的18.67%,随后降至28 cm的12.52%,在40 cm叶长级达最大值24.18%(图2).可见,顶冰花随叶长的增加(生长发育)生物量分配由地上生长逐渐转为地下生长,而后再次由地下转向地上(叶长30 cm以上)(图2).结合图1顶冰花的生长状况可以看出,顶冰花在叶长16~28 cm处于花期和花果并存期,在30 cm以上时进入果期,主要原因应为营养生长向生殖生长的转换.
2.2生物量模型构建
通过对顶冰花生物量与叶长间的相关分析可知,顶冰花的总生物量、地上生物量和地下生物量均与叶长存在极显著的正相关关系,这为构建生物量模型提供了可能.选用4种常见的回归方程(线性y=ax+b,指数y=aebx,对数y=a+blnx,幂函数y=axb),以叶长为自变量,拟合得到顶冰花地上、地下和全株生物量的估测模型.拟合的相关系数及参数见表2.

表2 顶冰花生物量估测模型
从各方程的拟合结果可以看出:除以指数方程拟合的地下生物量r值略低于0.90外,其余方程均在0.92以上,拟合效果较好.通过相关系数比较各方程的拟合结果可知,整体以直线方程最佳;其次为对数方程和幂函数;指数方程的拟合效果相对较差.各生物量拟合结果整体以总生物量和地上生物量拟合结果较好,而地下生物量相对较差.
2.3生物量模型验证
为检验顶冰花地上、地下和全株生物量估测模型在实际应用中的估测效果,将预留样本的实测数据代入表2的拟合方程,获取估测值,以总相对误差(RS)、平均相对误差(EE)、平均相对误差绝对值(RMA)、预估精度(P)进行预测效果比较,结果见表3.
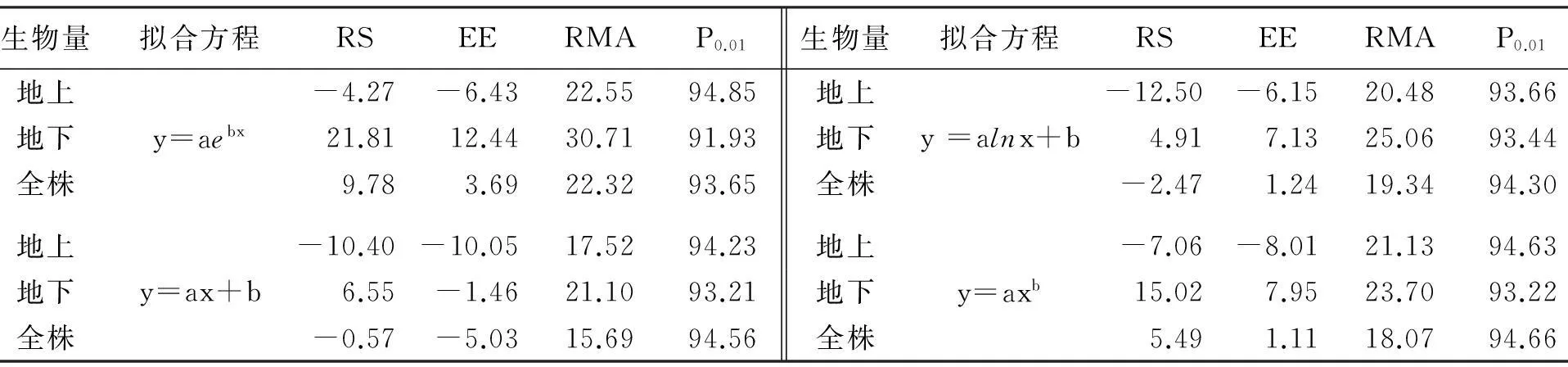
表3 模型验证
由表3可知:以叶长为自变量构建的顶冰花地上生物量、地下生物量和全株生物量模型,验证结果P值均达到90%以上,整体以直线和幂函数略高,对数方程次之,指数方程相对较差;仍表现出总生物量和地上生物量的精度略高于地下生物量.从RS和EE的结果来看,除以指数方程构建的总生物量模型预估结果高于实测值外,其余方程的预测值均低于总生物量和地上生物量的实测值;而4种方程构建的地下生物量预估测均高于其实测值.但从RS,EE和RMA的误差结果来看,除以指数方程拟合的地下生物量模型的RMA值略高于30%外,其余方程的误差结果均满足了相关建模精度要求(RS,EE和RMA<30%),均可以用于顶冰花生物量的估测[18-20].
3结论与讨论
生物量直接反映了生态系统中绿色植物的物质生产量,是生态系统获取能量能力的体现,对生态系统结构的形成及其功能具有十分重要的影响[21].生物量分配研究是分析植物物质分配的基础,根冠比不仅能够反映光合产物在地上和地下器官的不同投资分配,而且对地下生物量的估算也具有非常重要的价值[22-23].经统计可知:顶冰花随叶长级的增大,处于营养生长和开花阶段的植株个体显著减少,处于结果阶段的个体数量显著增多,即有性生殖与植株个体大小关系密切.单株生物量为0.316 2 g,地下生物量平均高于地上37.92%.叶生物量、果生物量、总生物量、地上生物量与地下生物量均表现出随叶长的增加而增大的变化,但在同等叶长间和不同叶长间均表现出较大的变异.顶冰花的根冠比平均为1.33,随叶长的增大呈单峰型变化,在叶长30 cm达到峰值;有性生殖器官占营养器官的比例介于5.78%~24.18%,平均为14.41%,随叶长的增加呈先增加后下降再增加的变化趋势.
相关研究表明,一次性结实草本植物的生殖分配一般占其净同化通量的20%~40%[24].而本研究获得的顶冰花有性生殖分配最大值仅为11.92%,远低于以往研究结果,这应与顶冰花为应对早春不利环境影响还存在克隆繁殖方式的生存策略有关.植物在易受干扰的不适生环境中,将光合积累的有限能量主要用于促进有性生殖;而在极端胁迫的生境中,则将通量的积累促进无性生殖[25-26].
生物量可通过收获法直接获得,该方法准确度最高,但对生态系统的破坏性大,而利用生物量模型进行估算已在大尺度生物量的估算中得到广泛应用[27].本研究以叶长级为自变量构建了顶冰花地上生物量、地下生物量和总生物量4种常用一元线性回归模型,并利用总相对误差、平均相对误差、平均相对误差绝对值、预估精度对模型进行了验证.结果表明:除以指数方程拟合的地下生物量模型的RMA值略高于30%外,其余方程的误差均满足相关建模精度要求,均可以用于顶冰花生物量的估测.受顶冰花物候期早和生活周期史紧凑的影响,研究采集样本已进入花、果期,植株个体相对较大,给生物量分配和模型构建的研究带来了一定的局限,在今后的研究中将进一步加以补充和完善,但通过此法可在不破坏资源的情况下较为准确地估算出顶冰花资源储量,进而服务于顶冰花资源的开发与利用等,研究的理论和现实意义均较大.
参考文献:
[1] 杜虎,宋同清,曾馥平,等.桂东不同林龄马尾松人工林的生物量及其分配特征[J].西北植物学报,2013,33(2):394-400.
[2]JacqueR,BemardS,HamldAM.Terrestrialglobalproductivity[M].SanDiegao:AcademicPress,2001.
[3]ZhaoM,ZhouGS.EstimationofbiomassandnetprimaryproductivityofmajorplantedforestsinChinabasedonforestinventorydata[J].ForestEcologyandManagement,2005,207:295-313.
[4]FangJ,ChenA,PengC,et al.ChangesinforestbiomasscarbonstorageinChinabetween1949and1998[J].Science,2001,291(5525):2320-2322.
[5]DepartmentF.Globalforestresourcesassessment2005.Progresstowardssustainableforestmanagement[M].Rome:ElectronicPublishingPolicyandSupportBranchCommunicationDivision,2005.
[6]TomppoE,GschwantnerT,LawrenceM,et al.Nationalforestinventories:pathwaysforcommonreporting[M].NewYork:Springer,2010.
[7] 漆良华,彭镇华,张旭东,等.退化土地植被恢复群落物种多样性与生物量分配格局[J].生态学杂志,2007,26(11):1697-1702.
[8] 张浩,宋同清,王克林,等.桂西地区不同林龄栎类群落的生物量及其分配格局[J].农业现代化研究,2013,34(6):758-762.
[9]CairnsMA,BrownS,HelmerEH,et al.Rootbiomassallocationintheworld’suplandforests[J].Oecologia,1997,111:1-11.
[10] 李晴,宋锦锦,邹春静.2种早春植物物候与小气候关系研究[J].上海环境科学,2010,29(4):148-152.
[11] 周永斌,殷有,于凤英,等.长白山阔叶红松林早春植物的养分吸收与归还[J].生态学杂志,2007,26(6):793-796.
[12] 刘红民.长白山阔叶红松林早春植物生长特征研究[J].吉林林业科技,2015,44(2):7-11.
[13] Sunmonu N,Ida T Y,Kudo G.Photosynthetic compensation by the reproductive structures in the spring ephemeralGagealutea[J].Plant Ecology,2013,214(2):175-188.
[14] 郑金萍,杨晶,郭忠玲,等.东北温带森林早春类短命植物种类及区系分布[J].北华大学学报(自然科学版),2014,15(4):516-521.
[15] 中国科学院中国植物志编辑委员会.中国植物志[M].北京:科学出版社,1980:14,72.
[16] 肖淑宁.顶冰花(漉林)中毒的临床分析[J].白求恩医科大学学报,1989,4(17):83-86.
[17] 范春楠,程岩,郑金萍,等.早春类短命植物生物量研究(Ⅰ):齿瓣延胡索生物量特征及模型构建[J].北华大学学报(自然科学版),2015,16(6):790-795.
[18] 范春楠,程岩,郑金萍,等.早春类短命植物生物量研究(Ⅱ):多被银莲花生物量特征及模型构建[J].北华大学学报(自然科学版),2016,17(1):24-29.
[19] 唐守正,张会儒,胥辉.相容性生物量模型的建立及其估计方法研究[J].林业科学,2000,36(专刊1):19-27.
[20] 范春楠,庞圣江,郑金萍,等.长白山林区14种幼树生物量估测模型[J].北京林业大学学报,2013,35(2):1-9.
[21] 宇万太,于永强.植物地下生物量研究进展[J].应用生态学报,2001,12(6):927-932.
[22] Cao K F,Ohkubo T.Allometry,root/shoot ratio and root architecture in understory saplings of deciduous dicotyledonous trees in central Japan[J].Ecological Research,1998,13:217-227.
[23] Gilmanov T G,PartonW J,Ojima D S.Testing the“CENTURY”ecosystem level model on data sets from eight grassland sites in the former USSR representing a wide climate/soil gradient[J].Ecological Modelling,1997,96:191-210.
[24] 祖元刚,张文辉,阎秀峰,等.濒危植物裂叶沙参保护生物学[M].北京:科学出版社,1999.
[25] 祖元刚,王文杰,杨逢建,等.植物生活史型的多样性及动态分析[J].生态学报,2002,22(11):1811-1818.
[26] 祖元刚,赵则海,杨逢建,等.植物生活史型的划分及其相互转化[J].生态学报,2005,25(9):2347-2353.
[27] Somogyi Z,Cienciala E,Mäkipää R,etal.Indirect methods of large-scale forest biomass estimation[J].European Journal of Forest Research,2007,126(2):197-207.
【责任编辑:郭伟】
On Spring Ephemeral Plants Biomass(Ⅲ)——Biomass Characteristics and Biomass Model Construction ofGagealutea
Zheng Jinping1,Fan Chunnan1,Cheng Yan2,Liu Bin1,Li Jinyu1
(1.ForestryCollegeofBeihuaUniversity,Jilin132013,China;2.AdministrationBureauofSanhuNatureReserveofSonghuaRiverinJilinProvince,Jilin132013,China)
Abstract:We investigated the biomass of Gagea lutea by using the whole harvest method,and analyzed the distribution characteristics of underground biomass and aboveground biomass,meanwhile,constructed biomass model of Gagea lutea.The results showed that:the mean individual biomass of Gagea lutea was 0.316 2 g.Mean biomass of underground was higher than that of aboveground about 37.92%.Leaf biomass,fruit biomass,total biomass,aboveground biomass and underground biomass were increased with leaf length enlarging,but flower biomass was decreased.There was a close relationship between sexual reproduction and individual size.The proportion of sexual reproduction biomass in total biomass was in a range from 5.47% to 19.42%,the mean was 11.92%;the proportion of sexual reproduction biomass in vegetation growth biomass was 14.41%,and showed rise firstly after the fall to rise again with leaf length enlarging.The root-shoot radio was from 0.93 to 1.86,the mean was 1.33,and assumed unimodal type change with the leaf length enlarging.In general,the increase of the Gagea lutea biomass with the leaf length manifested as aboveground growth transformed into underground growth,and the growth of vegetative organ transformed into the growth of reproductive organ.The four biomass estimation models of Gagea lutea with the leaf length including aboveground biomass,underground biomass and total biomass all reached very significant level,except model of underground biomass by index equation.
Key words:spring ephemeral plants;Gagea lutea;biomass;estimating model
中图分类号:Q948;S718.52
文献标志码:A
作者简介:郑金萍(1979-),女,博士,副教授,主要从事森林生态系统结构、功能及植被恢复等研究,E-mail:zhengjinping80@126.com.
基金项目:国家自然科学基金项目(31400387).
收稿日期:2015-09-07
文章编号:1009-4822(2016)02-0165-06
DOI:10.11713/j.issn.1009-4822.2016.02.005
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
农业工程学报(2022年10期)2022-08-22
林业调查规划(2022年3期)2022-06-13
世界竹藤通讯(2022年2期)2022-05-07
今日农业(2020年19期)2020-12-14
安徽农学通报(2020年7期)2020-05-26
学会(2016年12期)2017-01-13
科技创新导报(2016年23期)2016-12-23
中学教学参考·理科版(2016年9期)2016-12-15
亚太教育(2016年31期)2016-12-12
