
重金属(镉/铬/铅)胁迫对柳杉幼苗生长的影响及其富集特征
2016-04-17 12:22:12赵毅辉徐圆圆蒋维昕
贵州农业科学 2016年2期
赵毅辉,徐圆圆,蒋维昕,谭 玲,杨 梅*
(1.广西大学林学院,广西南宁530004;2.广西林业科学与工程高校重点实验室,广西南宁530004)
重金属(镉/铬/铅)胁迫对柳杉幼苗生长的影响及其富集特征
赵毅辉1,2,徐圆圆1,蒋维昕1,谭 玲1,杨 梅1,2*
(1.广西大学林学院,广西南宁530004;2.广西林业科学与工程高校重点实验室,广西南宁530004)
为寻求土壤重金属污染的生态修复新树种,采用砂土培养法,研究重金属镉(Cd)、铬(Cr)和铅(Pb)胁迫对柳杉幼苗生长的影响及其富集特征。结果表明:随着重金属胁迫浓度的升高,幼苗苗高及地径生长受抑程度、叶片电导率和丙二醛(MDA)含量也随之变大,尤其是对地径的抑制作用较为明显,300mg/L Pb胁迫下柳杉幼苗生长的增长率较CK低27.28%;柳杉叶片的总叶绿素含量则随重金属胁迫浓度的升高而降低,150mg/L Cr胁迫下柳杉叶片的叶绿素含量最低,为8.66mg/L,较CK差异显著;柳杉根系对重金属Cd、Cr和Pb的吸收作用明显,其富集系数最高,分别达17.71、10.45和6.87,其中Cd和Pb在柳杉植株中的吸收富集量依次为根>茎>叶,Cr依次为根>叶>茎;Cr不同胁迫浓度的转运系数均高于Cd和Pb,100mg/L Cr的幼苗转运系数达1.23,能有效吸收转运土壤中的重金属。柳杉对重金属污染具有一定的抗性,可将其作为低浓度重金属污染土壤的生态修复树种。
柳杉;重金属;抗性生理;生长抑制
工农业生产产生的重金属物质通过生物循环系统参与到环境中,当其被植物吸收后大量积累于植物的根、茎、叶中,严重时危害到植物的生长发育、生态环境及人类健康[1],镉(Cd)、铬(Cr)和铅(Pb)是目前环境中主要的重金属污染物,在Cd、Cr、Pb污染区域生长的香椿幼苗、桑树、樟树、栾树、香根草、水稻、玉米、油菜和小麦的细胞膜质、丙二醛(MDA)积累量和光合作用等均会受到严重影响[2-12],进而抑制作物的根系、茎、叶、芽的生长甚至死亡,其在作物中富集表现出显著毒害作用的同时,也通过食物链影响到人类的健康,治理重金属污染已刻不容缓。然而,采用物理、化学手段进行重金属污染修复常伴随着土壤结构的破坏、肥力及生物活性降低[13],寻求生态修复缓解重金属污染可行之有效地防止物理化学方法引起的土壤二次危害。Tlustos等[14-16]曾利用杨树(Populus)、柳树(Salix)、银杏(Ginkgo)和马尾松(Pinus massoniana)进行重金属污染土壤的生物修复研究,在含重金属土壤中,并非所有含量状态下的植物生长均表现为受毒害抑制。一定浓度的Cd可促进观光木和山白兰的生长,植物通过排斥和积累2种途径来减轻或消除其对自身的伤害[17],在高浓度的重金属土壤环境中,木本植物根系可从土壤中吸收、浓集和沉淀去除重金属污染物或通过迁移分配到植物的地上部分,玫瑰属植物通过产生新的重金属,芦苇、芥菜分别积累Cd等重金属于地下部分及叶片表皮毛中来对抗重金属的毒害[1820],利用植物的吸收富集作用治理土壤中的重金属污染具有良好的效果[21-23]。
柳杉(Cryptomeria fortunei)是我国的特有树种,其树冠高大、树干通直、木材纹理直、材质轻软,应用广泛,有学者从抗旱、抗寒及生长条件方面进行研究[24-26],而作为兼具景观功能和生态功能于一体的良好的绿化和环保树种,却鲜见柳杉在吸收富集重金属方面的研究报道。为此,研究柳杉在3种重金属(Cd/Cr/Pb)胁迫下的生长生理变化,分析其对重金属的富集转运特性,以期为其作为土壤重金属污染的生态修复树种提供理论依据。
1材料与方法
1.1试验材料
1.1.1柳杉幼苗1年生柳杉实生幼苗150株,广西国有六万林场。其播种期一致,未受环境污染,生长一致。
1.1.2供试基质 原土与沙子的混合物(森林原土∶沙子=9∶1),其Pb含量为11.26mg/kg,Cr含量为15.22mg/kg,Cd含量为0.33mg/kg,Pb 与Cr含量未超国家土壤环境质量Ⅰ级标准(清洁等级),Cd含量未超Ⅱ级标准[27]。
1.1.3仪器DDS-11A型电导率仪,上海康仪仪器有限公司;500-164-20型数显千分尺(Mitutoyo日本三丰和得力8208钢卷尺等。
1.2试验时间及地点
试验于2012-2013年在广西大学林学院研究教学基地(22.50°N,108.17°E)进行,该地区属南亚热带季风气候。将1年生柳杉实生幼苗移栽至等量基质的育苗钵中,共150钵。适应性培养10d后采用土壤浇灌法进行为期60d的重金属浇灌试验,重金属溶液胁迫处理时间为2012年11月11日至2013年1月31日,试验期间每天傍晚补充土壤水分,每隔10d停止浇水1d后进行重金属溶液浇灌,每次等量处理至盆栽土壤全湿润,且栽植盆底部刚好滴水为止,于2013年1月31日最后一次重金属浇灌处理10d后采样测定。
1.3试验设计
采用单因素随机区组设计,依据调查的南宁市土壤重金属含量[28],分别以Cr(NO3)3·9H2O、CdCl2·2.5H2O和Pb(AC)2·3H2O为重金属胁迫试剂设置3种重金属(镉/铬/铅)试验处理。重金属铅(以Pb2+计)设置100mg/L、200mg/L和300mg/L 3个浓度处理(即处理1~3,记为Pb-100,Pb-200,Pb-300),重金属镉(以Cd2+计)设20mg/L、40mg/L和60mg/L 3个浓度水平处理(即处理1~3,记为Cd-20,Cd-40,Cd-60),重金属铬(以Cr3+计)设置50mg/L、100mg/L和150mg/L(即处理1~3,记为Cr-50,Cr-100,Cr-150),3种重金属处理均以清水浇灌为对照(CK,即无重金属处理)。
1.4指标测定与计算
1)测定。苗高及地径,用数显千分尺测定地径(精确至0.01mm),得力8208钢卷尺测定苗高(精确至0.01cm);叶绿素含量,参照文献[29]的方法,经95%丙酮-乙醇提取,采用分光光度法测定;细胞质膜透性,参照文献[30]的方法,采用DDS-Ⅱ型电导率仪的测定;丙二醛(MDA)含量,参照文献[31]的方法,采用硫代巴比妥酸法测定;柳杉植株各器官、土壤中重金属含量,参照文献[32-33]的方法,采用ICP-AES法测定。
2)计算。参照文献[34-35]的方法计算生物富集系数和转运系数。
生物富集系数(BCF):BCF=Cp/Cs生物转运系数(BTF):BTF=Csl/Cr
式中,Cp为植物地上某部分重金属含量,Cs为对应土壤中重金属含量,Csl为地上部分的重金属含量,Cr地下部分的重金属含量。
1.5数据处理
采用Microsoft Excel 2003、SPSS19.0软件进行数据的处理,同时进行显著性检验(LSD法)。
2结果与分析
2.1重金属(镉/铬/铅)对柳杉幼苗生长的影响
2.1.1叶片形态 从表1可知:随着重金属镉(Cd)、铬(Cr)和铅(Pb)胁迫浓度的升高,柳杉幼苗叶片失绿黄化症越严重,当重金属Cd、Cr和Pb胁迫浓度分别达40mg/L、100mg/L和200mg/L时,柳杉叶片开始变黄,而后叶片出现较大面积斑点至幼苗接近枯萎死亡。Cd、Cr低浓度(20mg/L,50mg/L)和CK柳杉幼苗正常生长,随着Cd和Cr铬浓度的增加,柳杉幼苗叶片黄化斑点随之加大;Pb浓度为100mg/L时,柳杉幼苗叶片开始出现胁迫黄化现象,Pb浓度为300mg/L时,植株黄化症状最明显,受害最严重。

表1 重金属(镉/铬/铅)胁迫柳杉的叶片形态Table 1 Leaf morphology of C.fortunei seedlings under Cd,Cr and Pb stress
2.1.2苗高 从图1看出:柳杉幼苗的苗高增长率与重金属镉(Cd)、铬(Cr)和铅(Pb)胁迫浓度成反比。其中,Cr和Pb不同胁迫浓度下,柳杉的苗高生长均受到抑制,其增长率均小于CK(2.85%);Cd不同胁迫浓度,柳杉幼苗苗高的相对增长率最小为0.50cm,低于CK(0.38cm),Cd胁迫浓度为20mg/L时,苗高增长率比CK高0.20%,随着土壤中Cd浓度增加其苗高相对增长率对应降低至Cd胁迫浓度60mg/L时的1.41%,表明低浓度的Cd能促进柳杉幼苗的苗高增长。Cr和Pb不同胁迫浓度下,柳杉幼苗苗高与CK的相对增长率最大差值分别为1.14%和1.07%,幼苗苗高的相对增长率随着Cr和Pb浓度的增加而降低。其中,Pb胁迫下,苗高受抑程度较小,其相对增长率随着Pb胁迫浓度的增加呈平缓下降趋势,Cr胁迫浓度为50~100mg/L时,柳杉幼苗苗高的增长量最大。
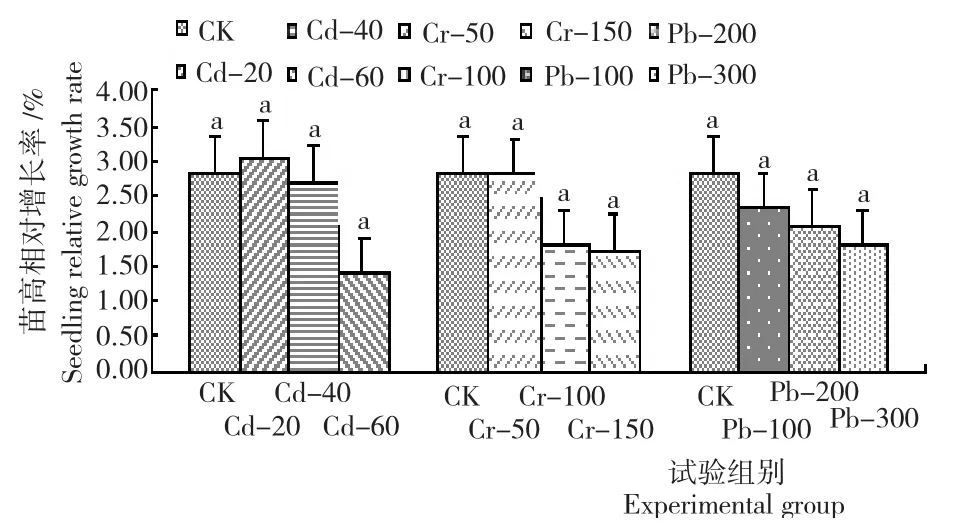
图1 重金属(镉/铬/铅)胁迫柳杉幼苗的苗高生长情况Fig.1 Height of C.fortunei seedlings under Cd,Cr and Pb stress
2.1.3地径生长 从图2看出:柳杉幼苗地径的相对增长率也与重金属镉(Cd)、铬(Cr)和铅(Pb)的胁迫浓度成反比。柳杉幼苗地径增长率均小于CK (1.65mm),相对增长34.55%。除Pb胁迫浓度为100mg/L和200mg/L外,其余处理差异均不显著;Cd胁迫浓度为20mg/L、40mg/L和60mg/L时,柳杉幼苗地径的相对增长率下降幅度随着Cd胁迫浓度增加而增大,其相对增长率分别为22.46%、10.97%和8.59%,且3种重金属胁迫对柳杉地径相对增长率的影响较苗高大。不同浓度Cr胁迫,柳杉幼苗地径的相对增长率波动小于2.89%,地径的最大增长量差值为0.98mm;Cr胁迫浓度为150mg/L时,幼苗地径的相对增长率较CK降低22.34%。不同浓度Pb胁迫,柳杉幼苗地径最大增长率较CK相差27.28%,最小为15.30%;Pb胁迫浓度为300mg/L时,柳杉幼苗的地径生长受抑制最严重。

图2 重金属(镉/铬/铅)胁迫柳杉幼苗的地径生长情况Fig.2 Ground diameter of C.fortunei seedlings under Cd,Cr and Pb stress
2.2重金属(镉/铬/铅)对柳杉叶片生理的影响
2.2.1细胞膜透性 从表2可知:柳杉幼苗叶片电导率随着镉(Cd)和铬(Cr)胁迫浓度的升高而升高,溶液浓度越高,叶片细胞膜受伤害越大。Cd和Cr胁迫浓度为20mg/L和50mg/L时,柳杉幼苗叶片电导率均低于CK。Cr胁迫柳杉幼苗叶片的电导率差异不显著。Cd胁迫则影响幼苗叶片的电解质渗漏,叶片质膜的结构和功能或丧失,导致质膜选择透过性减弱,细胞组织液的电导率逐渐增大,且随胁迫浓度的增加和处理时间的延长而加剧;Cd胁迫浓度为40mg/L时,柳杉叶片电导率达17.23%,显著高于CK;Cd胁迫浓度为60mg/L时亦显著高于CK。铅(Pb)胁迫浓度低于200mg/L时,柳杉叶片电导率差异不显著,且均低于18.17%;当Pb胁迫浓度达300mg/kg时,其电导率较CK提高7.28%,差异极显著。表明,高浓度Pb会致使柳杉叶片质膜结构受到损害,显著降低膜的选择透性。
2.2.2丙二醛(MDA)含量MDA是细胞膜发生膜脂过氧化作用的主要产物,其积累是活性氧对细胞膜毒害作用的具体表现。当重金属镉(Cd)、铬(Cr)和铅(Pb)胁迫浓度达最大时,柳杉MDA含量分别达85.74μmol/mg、58.84μmol/mg和82.26μmol/mg,较CK(35.61μmol/mg)差异极显著。最低Cd浓度胁迫柳杉叶片MDA含量与CK差异不显著;最低Cr浓度胁迫MDA含量为对照的1.32倍,差异不显著,随着浓度的继续升高其影响愈加显著;不同Pb浓度胁迫对柳杉叶片的MDA含量的影响均达极显著差异。

表2 重金属(镉/铬/铅)胁迫柳杉叶片的生理响应Table 2 Leaf physiological response of C.fortunei seedlings under Cd,Cr and Pb stress

表3 盆栽土壤及柳杉根、茎、叶中重金属(镉/铬/铅)的富集特征Table 3 Cd,Cr and Pb enrichment characteristics in root,stem and leaf of C.fortune and soil
2.2.3叶绿素含量 从表2可知,镉(Cd),随着Cd胁迫浓度的增加,柳杉叶片的叶绿素a呈降低→升高→降低的变化趋势,而叶绿素b和总叶绿素含量呈升高→降低的变化趋势。低浓度(20mg/L)Cd胁迫与CK相比差异不显著,表现为促进柳杉叶片叶绿素b的合成和抑制叶绿素a的合成;当Cd胁迫浓度达60mg/L时,叶绿素含量均下降,叶绿素a和总叶绿素含量下降显著。铬(Cr)胁迫,柳杉叶片叶绿素含量与Cr胁迫浓度呈反比,Cr胁迫浓度为50mg/L时,叶绿素b和总叶绿素含量分别为4.85mg/L和11.91mg/L,均高于CK;Cr胁迫浓度为150mg/L时,叶绿素b和总叶绿素含量较CK分别下降32.8%和20.6%,差异显著。说明,高浓度铬抑制叶绿素的合成,且对于叶绿素b的合成抑制程度高于叶绿素a。铅(Pb)胁迫,柳杉叶片叶绿素a和总叶绿素随浓度升高而逐渐降低,Pb胁迫浓度为100mg/L时,叶绿素b含量达最大值3.67 mg/L,其变化不显著;Pb胁迫浓度达300mg/L时,叶绿素含量降幅较大,叶片的失绿症状明显,高浓度Pb抑制柳杉叶绿素的合成,对叶绿素a的抑制程度大于叶绿素b。
2.3柳杉幼苗对重金属(镉/铬/铅)的富集作用
2.3.1镉(Cd) 从表3看出:镉在土壤及植株器官中的含量均随着胁迫浓度的升高而升高,柳杉幼苗各器官的富集系数及转运系数均在Cd胁迫浓度为40mg/L时达最大,其中根系中的Cd富集量最大,其富集系数最高,达17.71;而茎及叶的富集系数最高分别为1.51和0.18。在相同Cd胁迫浓度下,柳杉叶片的富集系数均低于1.0,并且随着重金属胁迫浓度的升高,Cd胁迫浓度为60mg/L时,柳杉根系对Cd的吸收富集量增加至176.10mg/kg,其富集系数达15.79,而转化系数仅0.04。
2.3.2铬(Cr)从表3看出:随着Cr胁迫浓度的升高,柳杉幼苗的根系、茎、盆栽土的铬含量也随之升高,不同胁迫浓度Cr富集量较CK均达极显著差异。其中,Cr胁迫浓度为50mg/L和100mg/L时,柳杉幼苗根系及叶片对Cr的富集系数均达8.20以上;Cr胁迫浓度为150mg/L时,柳杉叶片对Cr的吸收富集量为225.80mg/kg。经对盆栽土与幼苗根系、茎、叶片中铬含量的相关性分析,盆栽土与柳杉根系中的铬含量相关系数为0.98,柳杉根系对低浓度的铬具有较强的富集作用,其转运系数也较大,Cr胁迫浓度为100mg/L时,其转运系数为1.23。
2.3.3铅(Pb) 从表3可知:柳杉盆栽土及其根、叶中的铅含量较CK差异极显著,Pb在土壤及植株器官中的含量均随着胁迫浓度的升高而升高。Pb胁迫浓度为300mg/L时,柳杉各器官对Pb的吸收量最高,又以根系中的Pb富集量最大,其富集系数达6.87,而茎及叶的富集系数最高仅0.23和0.10。根系对Pb的富集量较大而转运系数较低,表明根是柳杉幼苗吸收土壤中Pb的主要器官。
3结论与讨论
1)当土壤重金属胁迫浓度提高时,Cd、Cr和Pb对柳杉幼苗苗高、地径生长的抑制作用越明显。水稻、月季、薤白在受到Cr、Pb胁迫时的作用也相同[3638],而Cd胁迫对植株苗高和地径生长的影响则因镉浓度、胁迫时间及植物基因型等因素的不同而有所差异。在对土沉香、美洲黑杨、香樟、龙葵、芥菜和青菜研究中发现,重金属的胁迫浓度越高,胁迫时间越长,对植物细胞膜的伤害越大[39-43];高浓度的Cd、Cr、Pb胁迫柳杉幼苗叶片的MDA含量及细胞膜透性升高,低浓度Cd、Cr、Pb胁迫柳杉叶片膜透性、MDA及叶绿素a含量较CK差异不显著;柳杉叶片叶绿素含量随Cr、Pb胁迫浓度的升高呈下降趋势,而Cd在低浓度范围内则促进叶绿素的生物合成,并且叶绿素a比叶绿素b对Cd反应更敏感[4347],当胁迫浓度继续升高时,重金属影响叶绿体中的类囊体排列规则,破坏其膜系统,导致叶绿体膨大而破坏其结构[48-50]。本研究表明,Cd胁迫浓度范围内叶绿素a/b及总叶绿素含量呈先上升后降低的趋势,在20mg/L Cd、50mg/L Cr、100mg/L Pb等低浓度胁迫下,柳杉幼苗的生理代谢稳定而维持植株的正常生长;日本楤木叶片对Cd2+胁迫也具有相同的响应特性[51]。因此,作物能够承受一定强度的逆境胁迫。
2)植物组织对重金属的转移和运输具有屏蔽作用,能阻止重金属从地下部向地上部分转移,大花萱草对铜镉的富集等试验表明,植株根系的重金属累积量大于茎叶[52-56]。本研究中,3种重金属在柳杉根系中的累积量最大,但其转运系数均较低,未达富集植物标准[57],这也是由于不同器官的外部形态和内部结构的不一致,吸收不同重金属的生理生化机制各异而导致的重金属元素的累积量差异性较大[5859]。柳杉具有庞大的地上生物量,在单位重金属富集量小的情况下也能吸收大量重金属,同时,重金属Cd、Cr、Pb在柳杉根系的最低富集系数也分别达10.61、8.23和3.62。说明,该树种具有富集重金属的作用,而且向地上部分传导的量较少,低浓度重金属胁迫不会影响柳杉幼苗的正常生理代谢及生长,因而在轻污染区域内,可尝试将柳杉作为重金属治理生态修复树种。
[1]梁 芳,郭晋平.植物重金属毒害作用机理研究进展[J].山西农业科学,2007,35(11):59-61.
[2]王利宝,朱宁华,鄂建华.Pb、Zn等重金属对樟树、栾树幼苗生长的影响[J].中南林业科技大学学报,2010,30(2):44-47.
[3]颜新培,龚 昕,唐汇清,等.镉超标土壤桑树修复研究进展[J].广东蚕业,2014,48(2):20-24.
[4]李文誉,李德明.盐碱及重金属对植物生长发育的影响[J].北方园艺,2010,12(8):221-224.
[5]曹 莹,黄瑞冬,曹志强.铅胁迫对玉米生理生化特性的影响[J].玉米科学,2005,13(3):61-64.
[6]江海东,周 琴,李 娜.Cd对油菜幼苗生长发育及生理特性的影响[J].中国油料作物学报,2006,28 (1):9-43.
[7]孔祥生,易现峰.植物生理学实验技术[M].北京:中国农业出版社,2008:248-250,257.
[8]杨树华,曲仲湘,王焕校.铅在水稻中的前一和富集以及对其生长的影响[J].生态学报,1986,6(4):312-322.
[9]林 杰,柯金炼,魏道军.铅胁迫对香根草叶片膜系统及抗氧化系统的影响[J].福建农业学报,2009,24 (3):254-257.
[10]徐美燕.镉铬胁迫以及硒对水稻幼苗生理生化影响的研究[D].广西:广西师范大学,2004.
[11]李晓晶.香椿对重金属的抗性研究[D].天津:天津理工大学,2012.
[12]Macfarlane G R,Burchett M D.Toxicity growth and accumulation relationships of copper lead and zinc in the grey mangrove Avicennia marina(Forsk)Vierh [J].Marine Environ Res,2002,54:65-84.
[13]Dermont G,Bergeron M,Mercier G,et al.Soil washing for metal removal:a review of physical/chemical technologies and field applications[J].Journal of Hazardous Materials,2008,152(1):1-31.
[14]Tlustos P,Pavl kov D,Sz kov J,et al.Exploitation of fast growing trees in metal remediation[M]//Mackova M,Dowling D,Macek T.Phytoremediation rhizoremediation.Germany:Springer,2006:83-102.
[15]曹福亮,郁万文,朱宇林.银杏幼苗修复Pb和Cd重金属污染土壤特性[J].林业科学,2012,48(4):8-13.
[16]陈文德,李贤伟,彭培好.马尾松林对土壤重金属的修复进程[J].林业科学,2008,44(5):6-10.
[17]谢安德.观光木和山白兰苗木的耐盐性和耐镉性研究[D].南宁:广西大学,2011.
[18]Rascio N.Metal accumulation by some plants growing on zinc-mine deposits[J].Oikos,1977,29:250-253.
[19]何勇强,陶勤南.Distribution of Ca in sogbean andquality of soybean seed under Cd strese[J].环境科学学报,2000,20(4):500-512.
[20]王志香,周光益,吴仲民,等.植物重金属毒害及其抗性机理研究进展[J].河南林业科技,2007,27(2):92-99.
[21]冯秀珍.闽北5种阔叶树抗镉胁迫能力的比较研究[J].安徽林业科技,2013,39(1):22-26.
[22]黄会一,张春兴,张有标.重金属镉﹑铅对木本植物光合作用影响的初步研究[J].生态学杂志,1986,5(2):6-9.
[23]Ali M B,Vajpayee P,Tripathi R D,et al.Phytoremediation of lead,nickel,and copper by Salixacmophylla Boiss:role of antioxidant enzymes and antioxidant substances[J].Bulletin of Environment Contamination and Toxicology,2003,70:462-469.
[24]代 莉,谢双喜.水分胁迫对日本柳杉种子萌芽的影响[J].贵州林业科技,2003,31(4):12-23.
[25]马定文,章定青.日本落叶松、日本柳杉雪折倒伏成因及预防措施浅析[J].湖北生态工程职业技术学院学报,2011,9(4):31-43.
[26]韦鹏练,李永忠,符韵林.柳杉木材干燥特性研究[J].林业实用技术,2012(2):25-27.
[27]周国华,秦绪文,董岩翔.土壤环境质量标准的制定原则与方法[J].地质通报,2005,24(8):721-727.
[28]刘宝庆.南宁市城区土壤重金属污染状况研究[D].南宁:广西大学,2004.
[29]杨敏文.快速测定植物叶片叶绿素含量方法的探讨[J].光谱实验室,2002,19(4):478-481.
[30]阮少宁,林 婷,王成伟,等.铝胁迫对不同杉木无性系质膜透性的影响[J].安徽农学通报,2008,14(19):135-136.
[31]张立军,樊金娟.植物生理学试验教程[M].北京:中国农业大学出版社,2007:259.
[32]吴建之,葛 滢.植物标样多元素测定的五种前处理方法比较[J].光谱学与光谱分析,1999,19(3):369-372.
[33]李清昌,王 微,刘 阁.ICP-AES法连续测定土壤样品中的铜、铅、锌、铁、锰[J].有色矿治,2012,28(2):97-106.
[34]马宏瑞,张 茜.长江南京段近岸沉积物中重金属富集特征与形态分析[J].生态环境学报,2009,18(6):2061-2065.
[35]卢德亮,乔 璐.哈尔滨市区绿地土壤重金属污染特征及植物富集[J].林业科学,2012,48(8):16-24.
[36]徐芬芬.铬胁迫对水稻幼苗生长和生理特性的影响[J].杂交水稻,2012,27(3):76-78.
[37]曾 俊,汪有良,王国良,等.镉铅胁迫对月季生长及生理生化特性影响[J].林业科技开发,2010,24(5):60-63.
[38]李 波,魏成熙,曾貂婵.重金属Pb对薤白生长发育的影响及其重金属积累特征[J].贵州农业科学,2010,38(2):59-61.
[39]Wang Q,Xiong D,Zhao P,et al.Effect of applying an arsenic-resistant and plant growth-promoting rhizobacterium to enhance soilarsenic phytoremediation by Populus deltoides LH05-17[J].Journal of Applied Microbiology,2011,111(5):1065-1074.
[40]王锦文,白 秀,等.复合重金属Pb/Zn对香樟生理特征的影响[J].安徽农业科学,2009,37(21):10253-10254,10257.
[41]任安芝,高玉葆,刘 爽.铬、镉、铅胁迫对青菜叶片几种生理生化指标的影响[J].应用与环境生物学报,2000,6(2):112-116.
[42]万文生,覃 静,陈卫国,等.Cd胁迫对土沉香幼苗生长及生理特性的影响[J].林业科技,2012, 37(1):1-3.
[43]王爱云,钟国锋,徐刚标,等.铬胁迫对芥菜型油菜生理特性和铬富集的影响[J].环境科学,2011,32(6):45-48.
[44]Baszynski.Photosynthetic activities of cadmium-treated tomato plants Physi-ol[J].Plant,1980,48:365-370.
[45]Sheoran I S,Singal H R,Randhir S.Effect of cadmium and nicked on photosynthesis and the enzymes of the photosynthetic carbon reduction cycle in pigeonpea(Cajanus cajan L.)[J].Photosynthesis Research.1990,23:345-351.
[46]Stobart A K.The effect of Cd on the biosynthesis of chlorophyll inleaves of barley[J].Physiol.Plant,1985,63:293-298.
[47]李荣春.镉铅及其复合污染对烟叶生理生化指标的影响[J].云南农业大学学报,1997,12(1):45-50.
[48]Mobin M,Nafees A K.Photosynthetic activity.Pigment composition and antioxidative response of two mustard(Brassica juncea)cultivars differing in photosynthetic capacity subjected to cadmium stress[J].Journal of Plant Physiology,2007,164(5):601-610.
[49]Hattab S,Dridi B,Chouba L,et al.Photosynthesis and growth responses of pea Pisum sativum L.under heavy metals stress[J].Journal of Environmental Sciences,2009,21:1552-1556.
[50]刘延盛,鲁家米,周晓阳.Pb在豌豆幼苗细胞中的超微结构分布与毒性研究[J].应用与环境生物学报,2007,13(5):647-651.
[51]孟 丽,李德生,李海茹,等.重金属Cd2+对日本楤木营养品质和抗氧化酶活性的影响[J].东北林业大学学报,2013,41(11):94-96.
[52]关梦茜,董 然.铜镉单一及复合胁迫下大花萱草对铜镉的富集特征[J].东北林业大学学报,2014,42 (9):138-142.
[53]唐贵才,董 伟.苎麻生长初期对重金属铅镉汞的迁移富集特征[J].贵州农业科学,2015,43(1):171-174.
[54]Smilde K W.Heavy-metal accumulation in crops grown on sewage sludge amended with metal salts [J].Plant and Soil,1981,62:3-12.
[55]Borovka L,kozak J.Heavy Metal Accumulation in Plants Grown in Heavily Polluted Soils[J].Folia Microbiol,1997,42(5):524-526.
[56]李松克,张春林,李克勤,等.多年生黑麦草对黄壤重金属污染的修复[J].贵州农业科学,2014,42(11):147-151.
[57]吴大付,张 莉,任秀娟,等.超富集植物研究新进展[J].河南科技学院学报,2011,39(3):55-59.
[58]Liang Chengfu,Chen Zhengfa,Liu Mingyue.Study processes on heavy metal pollution on vegetables[J].Hunan Agricultural Sciences,2002,16(4):45-48.
[59]何刚,耿晨光,罗 睿.重金属污染的治理及重金属对水生植物的影响[J].贵州农业科学,2008,36(3):147-150.
(责任编辑:杨 林)
Effect of Cd,Cr and Pb Stress on Growth of Cryptomeria fortunei Seedlings and Enrichment Characteristics of Heavy Metals
ZHAO Yihui1,2,XU Yuanyuan1,JIANG Weixin1,TAN Ling1,YANG Mei1,2*
(1.College of Forestry,Guangxi University,Nanning,Guangxi 530004;2.Guangxi Key Laboratory of Forestry Science and Engineering,Nanning,Guangxi 530004,China)
The effect of Cd,Cr and Pb stress on growth of Cryptomeria fortunei seedlings and enrichment characteristics of heavy metals were studied by the sandy soil culture method to seek new tree species for ecological remediation of the soil polluted with heavy metals.Results:The inhibition degree of height and ground diameter,leaf conductivity and MDA content of C.fortunei seedlings increase with increase of heavy metal concentration and heavy metal concentration has the obvious inhibition effect on ground diameter of C.fortunei seedlings.The increase rate of C.fortunei seedlings’growth under 300mg/L Pb stress is 27.28%lower than CK.Total leaf chlorophyll content of C.fortunei seedlings reduces with increase of heavy metal concentration and total leaf chlorophyll content of Cryptomeria fortunei seedlings under 150mg/L Cr stress is the minimum(8.66mg/L),significantly lower than CK.Cd,Cr and Pb enrichment coefficient of C.fortunei seedlings’root system reaches 17.71,10.45and 6.87respectively,higher than C.fortunei seedlings’leaf and stem.Cd and Pb enrichment amount in Cryptomeria fortunei seedlings is root>stem>leaf and Cr enrichment amount in Cryptomeria fortunei seedlings is root>leaf>stem.Cr transfer coefficient of C.fortunei seedlings under different Cr stress is higher than Cd and Pb.Cr transfer coefficient of C.fortunei seedlings under 100mg/L Cr stress is up to 1.23,which indicates that C.fortunei seedlings can absorb and transfer heavy metals in soil effectively.C.fortunei with a certain resistance against the soil polluted with heavy metals can be used as a tree species for ecological remediation of the soil slightly polluted with heavy metals.
Cryptomeria fortunei;heavy metal;resistant physiology;growth inhibition
S791.31;Q945.78
A
1001-3601(2016)02-0091-0166-06
2015-07-22;2016-01-29修回
国家自然科学基金项目“桉树人工林土壤酚酸物质积累机制及其环境行为”(31260176);“十一五”广西林业科技项目“六万林场柳杉大径材经营技术及木材综合利用研究项目”[桂林科字(2009)第23号]
赵毅辉(1989-),男,在读硕士,研究方向:用材林培育理论与技术。E-mail:fjzyh126@126.com
*通讯作者:杨 梅(1970-),女,教授,从事森林培育及林木抗逆性方面的教学与研究。E-mail:fjyangmei@126.com
猜你喜欢
安徽农学通报(2022年6期)2022-04-07 21:30:29
今日农业(2021年10期)2021-11-27 09:45:24
阅读(科学探秘)(2020年8期)2020-11-06 06:22:48
浙江林业(2020年7期)2020-08-06 14:55:32
中国果业信息(2019年1期)2019-01-05 17:41:42
中成药(2018年3期)2018-05-07 13:34:25
生物学教学(2017年9期)2017-08-20 13:22:32
喜剧世界(2016年7期)2016-08-31 06:21:06
中国火炬(2015年12期)2015-07-31 17:38:35
食品工业科技(2014年6期)2014-05-10 06:04:50
