
4种化学小分子联合诱导鼠胚胎成纤维细胞转化为施旺细胞效果观察
2016-04-08 23:47王永辉杨琼徐辉明高维强
山东医药 2016年18期
王永辉,杨琼,徐辉明,高维强
(1上海交通大学医学院附属仁济医院,上海200217;2上海交通大学Med-X临床干细胞研究中心)
4种化学小分子联合诱导鼠胚胎成纤维细胞转化为施旺细胞效果观察
王永辉1,2,杨琼1,2,徐辉明1,2,高维强1,2
(1上海交通大学医学院附属仁济医院,上海200217;2上海交通大学Med-X临床干细胞研究中心)
目的观察转化生长因子-β抑制剂SB431542、骨形态发生蛋白抑制剂Noggin、腺苷酸环化酶抑制剂FSK、 组蛋白去乙酰化酶抑制剂丁酸钠联合诱导鼠胚胎成纤维细胞(MEF)转化为施旺细胞的效果。方法将孕13.5 d雌鼠处死后取胎鼠分离MEF细胞,分为观察组和对照组;观察组置于含10 ng/mL碱性成纤维细胞生长因子、20 ng/mL表皮细胞生长因子、 20 ng/mL胶质细胞源性神经营养因子、 5 μmol/L 全反式维甲酸、10 μmol/L SB431542、200 ng/mL Noggin、10 μmol/L FSK、0.2 mmol/L丁酸钠的neurobasal培养基中诱导培养,隔天换液,共诱导3周,对照组不处理。在倒置相差显微镜下观察并记录两组转化细胞的形态,采用Q-PCR法检测两组施旺细胞标志物S100β、人蛋白脂质蛋白抗体(PLP)mRNA。结果诱导培养3周后可见部分MEF细胞变成梭形,胞体较小,两侧突起纤长,呈旋涡状排列。观察组S100β、PLP mRNA相对表达量分别为34.100±5.657、18.980±2.031,对照组分别为0.985±0.015、0.995±0.005(P均<0.05)。结论SB431542、Noggin、Forskolin和丁酸钠联合可诱导鼠胚胎MEF转化为施旺细胞。
大鼠;胚胎;成纤维细胞;施旺细胞;转化生长因子-β;骨形态发生蛋白;腺苷酸环化酶;组蛋白去乙酰化酶
脊髓损伤是脊柱损伤最严重的并发症,可导致损伤节段以下肢体严重功能障碍,目前临床尚无有效治疗方法。施旺细胞是周围神经主要的支持细胞,具有神经传导、神经保护、神经再生及神经营养作用[1,2]。最新研究发现,施旺细胞可促进损伤脊髓神经细胞修复[2~4],但具有再生困难等缺点。自体细胞直接诱导分化为施旺细胞是目前的研究热点[5]。S100β和人蛋白脂蛋白抗体(PLP)是施旺细胞的特异表达基因。2014年12月~2015年12月,我们观察了转化生长因子-β(TGF-β)抑制剂SB431542、骨形态发生蛋白(BMP)抑制剂Noggin、腺苷酸环化酶(cAMP)抑制剂FSK、 组蛋白去乙酰化酶抑制剂丁酸钠联合诱导鼠胚胎成纤维细胞(MEF)转化为施旺细胞的情况,旨在为脊髓损伤的细胞治疗提供参考。现报告如下。
1 材料与方法
1.1材料孕13.5 d的C57B1/6的雌性大鼠5只,体质量25~35 g,购自上海斯莱克公司;兔抗S100β单克隆抗体及兔抗PLP多克隆抗体均购自艾美捷公司,TAKARA逆转录试剂盒及核酸凝胶染料(SBYR Green)、碱性成纤维细胞生长因子(bFGF)、表皮细胞生长因子(EGF)、胶质细胞源性神经营养因子(GDNF),全反式维甲酸(RA)、SB431542、Noggin、FSK、丁酸钠等材料购自R&D system和Sigma公司,其余仪器购自Applied Biosystems公司, 10 % 胎牛血清(FBS)和1%青霉素-链霉素(P-S)购自Life technology公司。
1.2MEF细胞的分离将孕13.5 d雌鼠处死后取胎鼠,根据文献[6]方法分离MEF,去除胎鼠的脊髓以避免残留的神经干细胞及神经元的干扰。用含有10 % FBS和1% P-S的DMEM培养基在37 ℃、5% CO2培养箱中培养至第2代备用。
1.3细胞诱导培养取适量MEF,当细胞融合至80%~90%时传代至多聚赖氨酸预处理的培养皿和圆玻片上,第2天时吸出MEF培养基,PBS冲洗1次,置于含10 ng/mL bFGF、20 ng/mL EGF、 20 ng/mL GDNF、 5 μmol/L RA、10 μmol/L SB431542、200 ng/mL Noggin、10 μmmol/mL FSK、0.2 mmol/L丁酸钠的neurobasal培养基中,37 ℃、5% CO2环境下诱导培养,隔天换液,共诱导3周。
1.4细胞特性鉴定
1.4.1细胞形态观察取适量细胞,用PBS洗3次;4%多聚甲醛固定15 min,3%牛血清白蛋白封闭 1 h,再用一抗4 ℃孵育过夜。PBS洗3~5次,然后用相应的二抗室温避光孵育1 h。PBS洗3~5次,DAPI孵育3 min。封片后在倒置相差显微镜下观察,记录转化细胞的形态。
1.4.2细胞S100β、PLP mRNA检测 采用Q-PCR法。取适量诱导3周的MEF细胞(观察组)、方法1.2中分离的MEF细胞(对照组),PBS冲洗1次,提取总RNA,反转录生成1 μg cDNA,所有操作严格按照使用说明书进行。利用ABI PRISM 7900荧光实时定量PCR仪对扩增数据进行收集,并利用ABI公司的SDS2.3软件进行数据分析。以GAPDH为内参对照,采用ΔCt法计算两组细胞S100β、PLP mRNA相对表达量,实验重复3次。
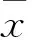
2 结果
诱导培养3周后,可见部分MEF细胞变成梭形,胞体较小,两侧突起纤长,呈旋涡状排列。观察组S100β、PLP mRNA相对表达量分别为34.100±5.657、18.980±2.031,对照组分别为0.985±0.015、0.995±0.005,观察组S100β、PLD mRNA相对表达量均高于对照组,P均<0.05。
3 讨论
神经细胞再生困难,多能干细胞[包括胚胎干细胞和诱导型多能干细胞(iPS)]分化、体细胞转分化是目前研究热点。施旺细胞主要分布在周围神经系统中神经元的突起周围,细胞形状不规则;其是周围神经纤维的鞘细胞,可反复包裹周围神经纤维的轴突形成髓鞘。研究发现,施旺细胞外表面有一层基膜,在周围神经再生中起重要作用[7, 8]。研究发现,脂肪干细胞[9]、间充质干细胞[10]和iPS[11]均可诱导分化为施旺细胞,但分化效率低,且iPS具有高致瘤性[12]。Vierbunchon等[6]通过病毒载体表达转录因子Ascl1、Brn2和Myt1l成功将MEF转分化为神经细胞,但病毒载体可导致基因组整合,染色体不稳定,最终导致细胞癌变。既往研究发现,化学小分子可将人体细胞直接转分化为神经元[16]和神经胶质细胞[5]。Thoma等[5]使用多种化学小分子成功将成纤维细胞转化为神经嵴细胞, 然后再进一步分化为施旺细胞。我们前期研究成功将MEF转分化为GABA能神经元[12,13]。
在施旺细胞的发育过程中,TGF-β会导致施旺细胞凋亡[17]。有实验证明,在神经元与施旺细胞共培养的体系中,TGF-β1可抑制树突与施旺细胞之间的信号传导,最终抑制施旺细胞增殖,抑制髓鞘的形成[18]。因此,本研究采用TGF-β抑制剂SB431542来抑制施旺细胞凋亡、促进施旺细胞增殖及髓鞘形成。而Noggin可通过抑制BMP,促进神经干细胞的增殖[19,20]。cAMP在抑制施旺细胞的增殖和分化成髓鞘细胞的过程中均起重要作用[21],而FSK可通过抑制其来降低cAMP水平;丁酸钠是将成纤维细胞转分化为施旺细胞诱导培养基中非常重要的成分[5]。另外,我们的培养体系中还添加了生长因子RA,原因是RA在神经系统形成、神经元的特化以及轴突生长过程中均起重要作用[22]。既往研究发现施旺细胞表面存在RA受体,提示RA在病理情况下和轴突的生长及再生发挥重要作用。此外,bFGF、GDNF和EGF均可促进神经系统髓鞘形成[23]。
综上所述,SB431542、Noggin、FSK和丁酸钠联合可诱导鼠胚胎MEF转化为施旺细胞,但其具体机制尚有待于进一步研究。
[1] Jessen KR, Mirsky R. The origin and development of glial cells in peripheral nerves[J]. Nat Rev Neuro Sci, 2005,6(9):671-682.
[2] Jessen KR, Mirsky R, Lloyd AC. Schwann Cells: Development and Role in Nerve Repair[J]. Cold Spr Har Persp Bio, 2015,7(7):020487.
[3] Zhang SQ, Wu MF,Liu JB, et al. Transplantation of human telomerase reverse transcriptase gene-transfected Schwann cells for repairing spinal cord injury[J]. Neural Rege Res, 2015,10(12):2040-2047.
[4] Namgung U. The role of Schwann cell-axon interaction in peripheral nerve regeneration[J]. Cells Tis Org, 2014,200(1):6-12.
[5] Thoma E, Merkl C, Heckel T, et al. Chemical conversion of human fibroblasts into functional schwann cells[J]. Stem Cell Res, 2014,3(4):539-547.
[6] Vierbunchon T, Ostermeier A, Pang ZP, et al. Direct conversion of fibroblasts to functional neurons by defined factors[J]. Nature, 2010,463(7284):1035-1041.
[7] Faroni A, Smith RJP, Lu L, et al. Human schwannlike cells derived from adiposederived mesenchymal stem cells rapidly dedifferentiate in the absence of stimulating medium[J]. Europ J Neuro Sci, 2016,43(3):417-430.
[8] Gao S, Zheng Y, Cai Q, et al. Different methods for inducing adipose-derived stem cells to differentiate into Schwannlike cells[J]. Arch Med Sci AMS, 2015,11(4):886-892.
[9] Zhu H, Yang A, Du J, et al. Basic fibroblast growth factor is a key factor that induces bone marrow mesenchymal stem cells towards cells with Schwann cell phenotype[J]. Neur Lett, 2013,559(5):82-87.
国家自然科学基金资助项目(31571399)。
高维强(E-mail: gao.weiqiang@sjtu.edu.cn)
10.3969/j.issn.1002-266X.2016.18.010
R329.2
A
1002-266X(2016)18-0032-03
猜你喜欢
昆明医科大学学报(2021年8期)2021-08-13
云南医药(2021年3期)2021-07-21
医学新知(2019年4期)2020-01-02
中国医药生物技术(2015年4期)2015-12-26
中国现代医学杂志(2015年26期)2015-12-23
化学反应工程与工艺(2015年3期)2015-04-16
中国当代医药(2015年33期)2015-03-01
西南国防医药(2015年11期)2015-02-28
疑难病杂志(2014年12期)2014-04-16
中国中医药现代远程教育(2014年17期)2014-03-01
