
燕山山脉不同居群豆茶决明叶片特征与光合特性
2016-04-05 00:34:51魏贤星许艳梅杜一鸣徐兴友
河北科技师范学院学报 2016年4期
常 旭,魏贤星,陈 健,许艳梅,杜一鸣,徐兴友*
(1 河北科技师范学院生命科技学院,河北秦皇岛,066600; 2 河北省青龙满族自治县农牧局;3北京林业大学园林学院)
燕山山脉不同居群豆茶决明叶片特征与光合特性
常 旭1,魏贤星1,陈 健2,许艳梅2,杜一鸣3,徐兴友1*
(1 河北科技师范学院生命科技学院,河北秦皇岛,066600; 2 河北省青龙满族自治县农牧局;3北京林业大学园林学院)
以燕山山脉5个居群豆茶决明为研究材料,系统比较了同一生境条件下各居群的叶片特征和光合特性。结果表明:承德兴隆居群叶形态参数值最大,而唐山迁西居群叶形态参数值最小。各居群不同生长时期叶绿素含量都呈逐渐下降趋势,植株处于营养生长期的叶绿素含量达到最大值,结荚期各居群叶绿素含量最低。豆茶决明的净光合速率日变化呈双峰曲线,都是先升高后降低,再升高,最后再降低,中间有明显的午休现象;各居群日平均净光合速率由高到低顺序为秦皇岛青龙居群,承德宽城居群,唐山遵化居群,承德兴隆居群,唐山迁西居群。豆茶决明各居群的蒸腾速率曲线变化及气孔导度变化与净光合速率曲线变化趋势相一致,而胞间CO2摩尔分数日变化趋势与净光合速率变化趋势相反。
豆茶决明;居群;光合生理特性;燕山地区
豆茶决明(Cassianomame(Sieb.)Kitag)为豆科云实亚科决明属一年生草本植物,是一种药食兼备的野生植物[1~3],在燕山山脉有着比较广泛的分布[4~6]。近年来,决明属植物活性成分和其药用价值都有实际应用和一定程度的开发,有关豆茶决明的研究主要偏重于组分及其药理作用[7~13],但光合特性研究还未见报道。不同居群豆茶决明的生长特性以及其对生态环境的适应性存在差异,豆茶决明的生长发育受不同生育期功能叶的光合特性、生理代谢和光合产物代谢的影响[27],为从整体上保持叶片的光合性能,增加光合物质的积累,提高豆茶决明产量,做进一步的药理研究,笔者以燕山山脉5个居群豆茶决明为研究材料,系统比较了同一生境条件下各居群光合特性差异,旨在为豆茶决明种质资源引种和合理开发提供理论依据[15~24]。
1 材料与方法
1.1 试验材料
供试豆茶决明种子是在2013年10月分别采自燕山山麓的青龙、宽城、兴隆、迁西、遵化等5个不同地区(表1)。自然风干后,选择饱满的种子,于2014年5月初将各居群播种在河北科技师范学院生命科技试验站,每个居群面积为5 m×1.2 m,株行距15 cm×20 cm,正常田间管理。

表1 豆茶决明采样点概况
1.2 试验试剂、仪器
丙酮:AR500ml(分析纯,天津试剂三厂);手持式GPS定位仪(型号explorist310,上海达赛导航设备有限公司);SF2000三按键电子显数游标卡尺(批号111N-103-10G,桂林广陆数字测控股份有限公司);便携式光合作用测定仪(型号GFS-3000,上海泽泉科技股份有限公司)。
1.3 试验方法
1.3.1 豆茶决明叶片大小测定 在每个居群中选取主茎上从上向下第6和第7片生长状况良好、无虫蛀、同一朝向的30个叶片,用分辨率0.01 mm显数游标卡尺测量小叶与复叶的长、宽、对数,计算复叶面积,3次重复。
1.3.2 叶绿素含量的测定 在2014年7月中旬(开花前期)、8月中旬(进入开花期)、9月中旬(结荚期)等3个生长阶段,采取各居群一定量健康功能叶,用体积分数为0.80的丙酮提取叶片叶绿素,采用752-型紫外可见分光光度法计测量各居群叶中叶绿素的质量分数[25]。
1.3.3 各气体交换参数的测定 于2014年8月中旬晴朗无云的某天,采用GFS-3000光合仪,在各豆茶决明居群中分别选取生长健康、光照比较均一的豆茶决明3株,每株上选择主茎中部位置、朝东南方向、具有正常功能并长势一致的叶片进行测定,取平均值,测定参数包括净光合速率(A)、蒸腾速率(E)、胞间CO2摩尔分数(Ci)、气孔导度(Gs)及光合有效辐射(PAR)等。测定植物光响应曲线时,光强梯度设置为1 800,1 500,1 200,900,600,300 μmol·(m2·s)-1共6个梯度。
1.3.4 数据分析 利用Excel 2003和SPSS 17.0对数据进行分析,分析结果以平均值和标准差的形式呈现。
2 结果与分析
2.1 不同居群豆茶决明叶片的形态特征
承德兴隆居群复叶及小叶长的值最大,唐山迁西居群复叶长及小叶长的值最小,承德宽城居群复叶及小叶宽值最大,唐山迁西居群复叶及小叶宽值最小,承德兴隆居群小叶对数最多,叶面积也是最大的,唐山迁西居群叶面积最小;唐山迁西居群复叶和叶面积大小与其他居群间具有极显著差异;承德兴隆豆茶决明的小叶最长,极显著长于承德宽城、秦皇岛青龙和唐山迁西的小叶,而唐山迁西豆茶决明的小叶最短;唐山迁西、遵化豆茶决明的小叶最窄,极显著于承德宽城、兴隆和秦皇岛青龙,而承德宽城的小叶最宽;承德兴隆豆茶决明的小叶对数最多,极显著于唐山迁西的小叶对数,唐山迁西的小叶对数最少;唐山迁西豆茶决明的复叶最短,极显著短于承德宽城、兴隆、秦皇岛青龙和唐山遵化的复叶,而唐山遵化的豆茶决明的复叶最长;秦皇岛青龙豆茶决明的复叶最宽,极显著宽于承德宽城、兴隆、唐山遵化和迁西的复叶,而唐山迁西的豆茶决明的复叶最窄;承德兴隆豆茶决明的叶面积最大,极显著大于唐山迁西的叶面积,而唐山迁西的豆茶决明的叶面积最小(表2,表3)。

表2 不同居群豆茶决明小叶形态参数
注:大写字母表示0.01水平上差异极显著,小写字母表示0.05水平上差异 显著。表3同。
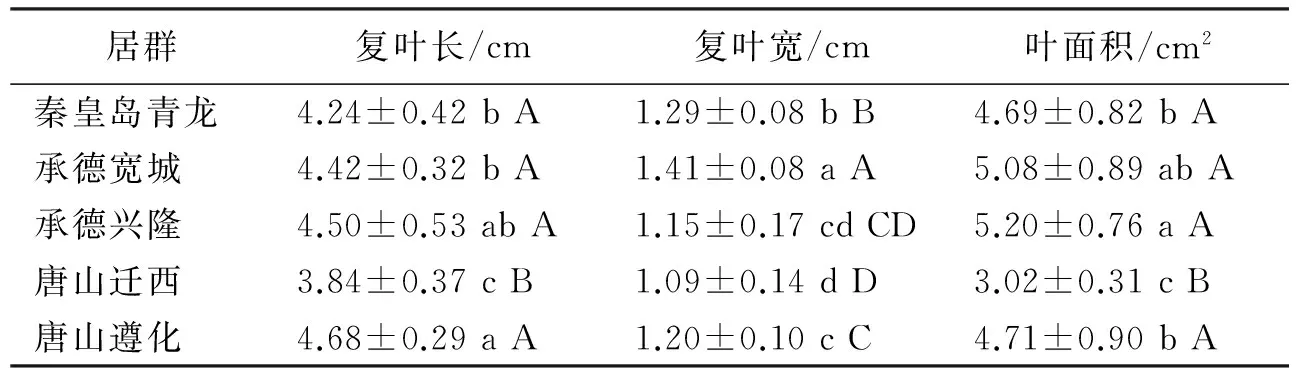
表3 不同居群豆茶决明复叶的形态参数
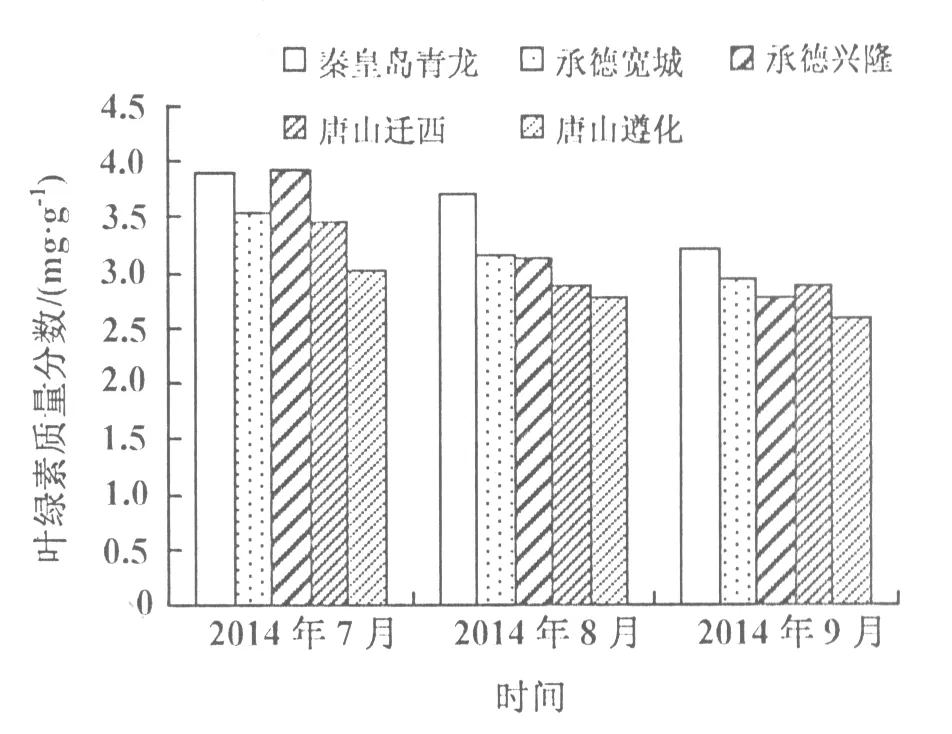
图1 不同居群豆茶决明叶片叶绿素质量分数的变化
2.2 不同居群豆茶决明叶片叶绿素质量分数的变化
各居群叶绿素含量都呈逐渐下降趋势,变化较为一致(图1)。不同居群豆茶决明在7月份植株处于营养生长期的叶绿素含量达到最大值,到9月份结荚期各居群叶绿素含量最低。7月中旬秦皇岛青龙居群叶绿素总质量分数最高,为3.90 mg/g,而唐山遵化居群叶绿素总质量分数最低,为3.01 mg/g;9月中旬各居群叶绿素总质量分数高低的变化趋势基本保持不变,秦皇岛青龙居群叶绿素总质量分数仍最高,为3.22 mg/g,唐山遵化居群叶绿素总质量分数最低,为2.59 mg/g。
2.3 不同居群豆茶决明叶片气体交换参数的日变化
2.3.1 各居群净光合速率变化 5个豆茶决明居群净光合速率日变化趋势较为一致,都是先升高,后降低一定值,再升高,最后下降,呈现双峰型,有明显的午休现象(图2)。各居群在上午10:00都有个最高峰,即净光合速率最大,此时秦皇岛青龙、承德宽城、承德兴隆、唐山迁西及唐山遵化居群的净光合速率值分别是10.86,6.90,7.62,7.09,8.76 μmol·(m2·s)-1。在下午14∶00出现第二次小高峰,各居群此时的净光合速率值分别是7.24,5.99,4.37,2.27,3.57 μmol·(m2·s)-1。秦皇岛青龙居群净光合速率值全天高于其他居群,唐山居群的净光合速率一直处于较低状态;在同一环境条件下,各居群相比较,日平均净光合速率由高到低的顺序为秦皇岛青龙居群,承德宽城居群,唐山遵化居群,承德兴隆居群,唐山迁西居群。
2.3.2 不同居群蒸腾速率日变化 各豆茶决明居群蒸腾速率曲线呈单峰值型变化,先上升后下降,在上午10:00前后,各居群蒸腾速率均出现最大值,分别为5.23,4.81,3.32,4.32,3.97 mmol·(m2·s)-1;各居群蒸腾速率日变化平均值由大到小的顺序为秦皇岛青龙居群,唐山遵化居群,承德宽城居群,承德兴隆居群,唐山迁西居群(图3)。在10:00~14:00之间,秦皇岛青龙居群的蒸腾速率高于其他居群,秦皇岛青龙居群可以很好地利用有效蒸腾作用来适应高温环境。

图2 不同居群豆茶决明净光合速率日变化 图3 不同居群豆茶决明蒸腾速率日变化
2.3.3 不同居群胞间CO2摩尔分数日变化 胞间CO2摩尔分数 (Ci)是外界CO2进入植物叶肉细胞过程中,所受各种驱动力和阻力以及叶片内部光合作用和呼吸作用的最终平衡结果[24]。5个居群豆茶决明胞间CO2摩尔分数(Ci)的日变化趋势较为一致,日变化曲线也呈双峰型,各居群胞间CO2摩尔分数日变化均是呈现先降低后又逐渐上升,再下降后快速上升的走向,这与净光合速率日变化曲线正好相反(图4)。5个豆茶决明居群胞间CO2摩尔分数变化范围在200~700 μmol·mol-1之间,各居群在12:00前后出现一个小波峰,在16:00以后5个豆茶决明居群胞间CO2摩尔分数均快速上升,在18:00时,5个居群胞间CO2摩尔分数达到最大值;在各居群胞间CO2摩尔分数相比较之下,承德宽城居群胞间CO2摩尔分数日变化平均值最低,为277.72 μmol·mol-1,而唐山遵化居群胞间CO2摩尔分数日变化平均值最高,为338.92 μmol·mol-1。
2.3.4 不同居群气孔导度日变化 5个豆茶决明居群的气孔导度与净光合速率日变化趋势是一致的(图5),在上午10:00及下午14:00前后出现2个峰值,14:00之后随着光强、气温的下降而迅速降低;气孔导度日均值由大到小的顺序为秦皇岛青龙居群,唐山遵化居群,承德宽城居群,承德兴隆居群,唐山迁西居群。光合速率和蒸腾速率均与气孔导度有一定关系,在10:00~14:00 秦皇岛青龙居群的气孔导度明显高于其它居群,该居群通过有效的调控气孔开闭,提高自身的蒸腾作用和光合作用。

图4 不同居群豆茶决明胞间CO2摩尔分数日变化 图5 不同居群豆茶决明气孔导度日变化
2.4 不同居群豆茶决明叶片的光响应
2.4.1 净光合速率对光强的响应 5个豆茶决明居群的净光合速率均随着光照强度的增加而慢慢增大(图6),在光强300~900 μmol·(m2·s)-1范围内,各居群净光合速率都缓慢变大,在光强900~1 800 μmol·(m2·s)-1,秦皇岛青龙居群的净光合速率随光强增加而迅速增加,其他居群的净光合速率增加相对较缓慢;在光强300~1 800 μmol·(m2·s)-1之间,秦皇岛青龙居群的净光合速率始终高于其他居群,而承德兴隆居群的净光合速率始终保持最低水平,这也说明秦皇岛青龙居群适应光强环境的能力更强,唐山遵化居群更适应低光强环境。
2.4.2 蒸腾速率对光照强度变化的响应 承德兴隆居群及唐山遵化居群豆茶决明的蒸腾速率随光强的增加呈先降低后升高的变化趋势,而其他3个居群的蒸腾速率则随光强的增加而升高(图7)。光照强度在300~1 800 μmol·(m2·s)-1范围内,5个豆茶决明居群的蒸腾速率由高到低的顺序为秦皇岛青龙居群,承德宽城居群,唐山迁西居群,承德兴隆居群,唐山遵化居群。

图6 豆茶决明净光合速率对光强的响应 图7 豆茶决明蒸腾速率对光照强度变化的响应
2.4.3 气孔导度Gs对光照强度变化的响应 秦皇岛青龙居群豆茶决明随光照强度的增加,气孔导度呈现先缓慢上升,后快速升高的变化趋势;其他居群的气孔导度亦均随光强的增加均呈现缓慢、平稳上升的特点(图8)。在光强300~900 μmol·(m2·s)-1范围内,5个豆茶决明居群的气孔导度都处于很低的水平;在光强高于900 μmol·(m2·s)-1时秦皇岛青龙居群的气孔导度快速升高,且明显高于其他居群;到1 800 μmol·(m2·s)-1时,5个居群的气孔导度均达到最大值,此时秦皇岛青龙居群气孔导度为30.40 mmol·(m2·s)-1,承德宽城为15.44 mmol·(m2·s)-1,承德兴隆居群为11.46 mmol·(m2·s)-1,唐山迁西居群为8.82 mmol·(m2·s)-1,唐山遵化为4.54 mmol·(m2·s)-1。另外,5个居群的气孔导度对光强的响应变化趋势均与其蒸腾速率、净光合速率的变化趋势相一致,说明气孔导度和蒸腾速率、净光合速率有很强的正相关性和一致性,气孔因素是影响蒸腾速率的最主要因素之一,也能很好反映光合作用的强弱。
2.4.4 胞间CO2摩尔分数对光照强度的响应 5个豆茶决明居群胞间CO2摩尔分数光响应曲线,在整体上均呈随光强的增加一直下降的特点(图9)。在600~1 800 μmol·(m2·s)-1区间,5个居群的胞间CO2摩尔分数快速下降,这与其净光合速率对光响应的变化趋势大体呈相反的变化规律,说明净光合速率大时,有更多的CO2用于光合,从而使胞间CO2摩尔分数降低,表现为净光合速率与胞间CO2摩尔分数的负相关性。

图8 豆茶决明气孔导度Gs对光照强度变化的响应 图9 豆茶决明胞间CO2摩尔分数对光照强度的响应
3 结论与讨论
光合作用是源物质积累的生理基础,光合速率是光合作用的重要体现[14]。叶片形态会影响植物的光合能力,但植物光合作用不只受叶片大小单因素的影响,还受叶片叶绿素质量分数、叶蒸腾速率、叶气孔导度等多因素共同的作用;叶绿素质量分数是与光合性能密切相关的重要生理特性,气孔导度通常是影响光合速率的初始因素[26,27]。通过对5个不同居群豆茶决明叶片的形态、叶绿素含量及各光合参数分析,发现叶片的形态大小与净光合速率未有明显的正相关,但叶绿素含量与光合能力大小有一定关系,植物的光合作用受叶片大小、叶绿素含量及气孔等多因素的影响。承德兴隆居群叶形态参数值最大,唐山迁西居群叶参数值最小;各居群不同生长时期叶绿素质量分数都呈逐渐下降趋势,变化较为一致。植株处于营养生长期的叶绿素质量分数达到最大值,结荚期各居群叶绿素质量分数最低,秦皇岛青龙居群叶绿素总质量分数最高,为3.22 mg/g,唐山遵化居群叶绿素总质量分数最低,为2.59 mg/g。
通过对5个豆茶决明居群的各气体交换参数分析,发现5个居群的净光合速率曲线呈双峰型变化,都是先升高后降低,再升高,最后降低,中间有明显的午休现象;5个居群在10:00 时出现净光合速率最大值,在14:00时均出现一个小峰值。秦皇岛青龙居群的最大净光合速率明显高于其他居群,在高光强条件下,净光合速率也显著高于其他居群,各居群日平均净光合速率由高到低的顺序为秦皇岛青龙居群,承德宽城居群,唐山遵化居群,承德兴隆居群,唐山迁西居群。豆茶决明居群的蒸腾速率曲线变化及气孔导度曲线与净光合速率曲线变化趋势相一致,豆茶决明居群的气孔导度与蒸腾速率存在着紧密的相关性,两者的变化均表现为在较高光强下由大到小的顺序为秦皇岛青龙居群,承德宽城居群,唐山遵化居群,承德兴隆居群,唐山迁西居群。在各居群胞间CO2摩尔分数日变化过程中,胞间CO2摩尔分数随光强的增加先下降后上升,下降后再上升,其变化趋势均与其净光合速率、气孔导度的变化趋势相反。
本次研究是首次探究豆茶决明的光合特性、相关参数变化,对豆茶决明的种质的归类、栽培、品种选育等有非常重要的意义。植物光合作用日变化进程,反映了一天中植物进行物质积累与生理代谢的持续能力[29]。5个豆茶决明居群的净光合速率日变化均呈双峰型,在中午高温、高光强辐射条件下,由于叶片气孔开张程度受到影响,净光合速率均呈下降趋势,能够吸收较强的光能,促进有机物的积累,为果实后熟阶段的油脂转化提供营养。相比而言,秦皇岛青龙居群的光合特性更强,能够通过调节自身的蒸腾速率和气孔的张开程度、提高自身的光合能力来适应高温、强光的环境。
[1] 河北植物志编辑委员会.河北植物志:第一卷[M].石家庄:河北科学技术出版社,1986:693-696.
[2] 中国科学院中国植物志编辑委员会.中国植物志:第三十九卷[M].北京:科学出版社,1998:128.
[3] 国家药典委员会.中国药典(2005年版一部及补充检验方法)[M].北京:化学工业出版社,2005.
[4] 尹秀玲,温静,张斌,等.秦皇岛市野生药用植物资源的调查研究[J].农家之友,2008(10):19-23.
[5] 韩宝强.河北省生物柴油能源植物种质资源研究[D].秦皇岛:河北科技师范学院,2014:12-70.
[6] 徐兴友.老岭自然保护区主要野生药用被子植物资源研究[J].河北职业技术师范学院学报,2000,14(2):55-58.
[7] 徐义龙.决明子化学成分及炮制对其影响研究[D].北京:中国中医科学院中药研究所,2014:1-47.
[8] 李婷,冯占民,杨巡纭,等.决明属植物的化学成分及药理作用研究进展[J].林业化学与工业,2012,32(6):107-118.
[9] 张玥,张秋华,杨楚枫,等.豆决明中木犀草苷的提取及含量测定[J].中国中医药现代远程教育,2015,13(2):142-144.
[10] 赵东利,李东霞,王仁军.豆茶决明(Cassianomame)的核型分析[J].大连大学学报,1998,19(4):53-55.
[11] 杨楚枫,杨洋,张玥,等.豆茶决明对高脂诱导C57BL/6J小鼠肥胖和胰岛素抵抗的影响[J].中国实验方剂学杂志,2015,21(10):141-145.
[12] 王芳生,丁艳丽,邓旭明,等.从豆茶决明中分离蒽醌酮的结构鉴定[J].中国药学杂志,2000,35(s1):37-38.
[13] Kitanaka S,Takido M.Demethyltorosa flavones C and D fromCassianomame[J].Phytochemistr,1992,31(8):2 927-2 929.
[14] 左振朋,孙庆泉,董鲁浩,等.爆、甜、糯玉米生育后期叶片光合特性的比较[J].作物学报,2009,35(10):1 930-1 935.
[15] 王康才,王立会,汤兴利,等.江苏地区3个居群细柱五加叶片显微结构及光合特性研究[J].安徽农业大学学报,2011,38(5):651-655.
[16] 左应梅,张金渝,杨维泽,等.珠子参不同居群的光合特性及保护酶活性比较[J].广东农业科学,2014(23):16-19,25.[17] 赵相勇,吴佳海,杨菲.喀斯特山区混播草地草本植物的光合特性[J].贵州农业科学,2011,39(3):46-49.
[18] 程建峰,沈允钢.试析光合作用的研究动向[J].植物学报,2011,46(6):694-704.
[19] 肖亚雯,王康才,杨磊,等.不同居群半夏光合及荧光动力学研究[J].中国中药杂志,2009,34(11):1 339-1 342.
[20] 呼和牧仁,周梅,翟洪波,等.影响树木光合作用因素的研究进展[J].内蒙古农业大学学报,2009,30(2):287-291.
[21] 贺安娜,滕攀.9个不同居群虎耳草光合特性及有效成分含量分析[J].怀化学院学报,2014,33(11):6-9.
[22] Quick W P,Chaves M M,Wendler R,et al.The effect of water stress on photosynthetic carbon metabolism in four species grown under field conditions[J].Plant Cell and Environment,1992,15(1):25-35.
[23] 任丽花,王义祥,翁伯琦,等.土壤水分胁迫对圆叶决明叶片含水量和光合特性的影响[J].厦门大学学报(自然科学版),2005,44(增刊):28-31.
[24] 姚春娟,郭圣茂,陈兰兰,等.二种决明属植物叶绿素荧光特性的比较研究[J].北方园艺,2016(02):151-153.
[25] 刘永军,郭守华,杨晓玲.植物生理生化实验[M].北京:中国农业科技出版社,2002.
[26] 王艳霞,李双双,李华兴.垃圾填埋场周边桉树光合生理特性研究[J].生态环境学报,2012,21(5):919-923.
[27] 徐俊华,荒漠草原豆科与禾本科牧草光合特性及产量研究[J].广东农业科学,2014(13):17-24.
(责任编辑:朱宝昌,陈于和)
On Photosynthetic Physiological Characteristics of DifferentCassianomamePopulations in Yanshan Mountain
CHANG Xu1, WEI Xianxing1, CHEN Jian2, XU Yanmei2, DU Yiming3, XU Xingyou1
(1 College of Life Science & Technology, Hebei Normal University of Science & Technology, Qinhuangdao Hebei, 066600; 2 Agriculture and Pasture Bureau, Hebei Qinglong Manchu Autonomous County; 3 School of Landscape Architecture, Beijing Forestry University; China)
FiveCassianomamepopulations in Yanshan mountain were used to study the differences of leaf morphology and photosynthetic physiological characteristics. The results showed that the maximum leaf morphological parameters belonged to Chengdexinglong population, and the minimum to Tangshanqianxi population. The chlorophyll contents of all populations displayed a gradual downward trend, while the total chlorophyll contents might reach the peak in the vegetative stage and in contrast might reach the bottom in the pod stage. The diurnal variation of the net photosynthetic rate ofCassianomameexhibited a double-peak curve, i.e. increased first and then reduced and increased again and reduced. The midday depression in diurnal variation was obviously observed. The daily average net photosynthetic rate of differentCassianomamepopulations ranged from Qinhuangdaoqinglong population, Chengdekuancheng population, Tangshanzunhua population, Chengdexinglong population to Tangshanqianxi population. The diurnal variation of transpiration rate and stomatal transmitting degree approached with the diurnal variation of net photosynthetic rate but the diurnal variation of intercellular CO2concentration tended to be opposite.
Cassianomame; population;photosynthetic physiological characteristics;Yanshan area
10.3969/J.ISSN.1672-7983.2016.04.008
河北省研究生创新资助项目。*通讯作者,男,教授,博士,硕士研究生导师。主要研究方向:野生植物资源的保护与利用。E-mail: xuxingyou@126.com。
2016-10-11; 修改稿收到日期: 2016-12-07
S567.21+9
A
1672-7983(2016)04-0046-06
常旭(1988-),女,硕士研究生,主要研究方向:植物资源开发与利用。
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26 08:55:24
浙江中医药大学学报(2021年6期)2021-07-12 03:05:14
节水灌溉(2021年5期)2021-05-26 06:49:38
中国农业科技导报(2020年12期)2020-03-15 10:50:08
草地学报(2018年5期)2018-11-07 02:25:00
新课程·下旬(2017年7期)2017-08-14 11:12:50
西北林学院学报(2015年5期)2015-01-03 05:12:20
赤峰学院学报·自然科学版(2012年19期)2012-10-14 02:53:58
植物营养与肥料学报(2011年2期)2011-10-26 03:52:46
植物营养与肥料学报(2011年2期)2011-10-26 03:52:30
