
白菜型与甘蓝型冬油菜抗寒机理差异的研究
2016-03-18 07:59:05孙万仓刘自刚赵艳宁刘海卿武军艳
华北农学报 2016年1期
马 骊,孙万仓,刘自刚,赵艳宁,杨 刚,刘海卿,武军艳,方 彦,
李学才,刘林波,钱 武,侯献飞
(甘肃省油菜工程技术研究中心,甘肃省作物遗传改良与种质创新重点实验室,
甘肃省干旱生境作物学重点实验室,甘肃农业大学 农学院,甘肃 兰州 730070)
白菜型与甘蓝型冬油菜抗寒机理差异的研究
马骊,孙万仓,刘自刚,赵艳宁,杨刚,刘海卿,武军艳,方彦,
李学才,刘林波,钱武,侯献飞
(甘肃省油菜工程技术研究中心,甘肃省作物遗传改良与种质创新重点实验室,
甘肃省干旱生境作物学重点实验室,甘肃农业大学 农学院,甘肃 兰州730070)
摘要:为探究白菜型与甘蓝型冬油菜抗寒机理差异的原因。以8个抗寒性不同的冬油菜品种为材料,采用大田试验和盆栽试验相结合的方法,待油菜长至5~6片真叶时,大田试验进行植物学形态,干物质积累量的测定,盆栽试验按24 ℃→10 ℃→5 ℃→0 ℃→-5 ℃→-10 ℃各48 h依次降温处理后测定生理生化指标。结果表明,白菜型冬油菜冬前生长点洼陷,幼苗匍匐生长,干物质积累主要集中在地下部分,其中白菜型冬油菜地下部鲜质量与地下部干质量较甘蓝型冬油菜平均增加了236.1%,263.0%,说明抗寒性强的冬油菜能够在营养生长阶段将光合有机产物优先运输到地下部,建立庞大的根系,为安全越冬提供代谢能量。随着温度的变化,不同类型冬油菜的生理生化活性有较大的差异,-5 ℃时陇油7号SOD活性较CK增加了10.7%,0 ℃时陇油7号 CAT、POD 活性较CK分别增加了24.7%,28.6%,而0 ℃时白菜型冬油菜SP含量较甘蓝型冬油菜平均增加了32.3%,-10 ℃时白菜型冬油菜的SS含量较甘蓝型冬油菜平均增加了71.4%,-10 ℃甘蓝型冬油菜MDA含量较白菜型冬油菜平均增加了52.8%,这说明抗寒性强的品种在低温条件下能够保护自身免受损伤,其中CAT、POD、SP是抵抗冷害的保护性物质,SOD、SS是抵御冻害的保护性物质。白菜型冬油菜比甘蓝型冬油菜在形态学及生理水平上都具有明显的优势,形态学上的优势使其有利于抵御极端低温天气,提供维持越冬及冬后返青所需的代谢能量;生理水平上,低温胁迫后保护性酶活性、调节性物质含量增加,能够有效地保护细胞膜结构,MDA含量减少,可以缓解低温对冬油菜叶片的伤害,从而保证高越冬率,为北方白菜型与甘蓝型冬油菜抗寒性研究提供理论依据。
关键词:白菜型;甘蓝型;冬油菜;抗寒性;形态特征;干物质积累;生理生化;隶属函数
中国北方大部分地区一季有余,两季不足,属于一年一熟制区,冬油菜成熟期早,可在5月下旬-6月上中旬成熟收获,收获后可复种玉米、马铃薯、大豆、荞麦、糜子、籽瓜等,可改变一年一熟为一年两熟或两年三熟,具有较高的经济效益[1]。同时,产量和生态效益均高于同类地区胡麻和春油菜。但我国北方地区冬寒春旱,生态条件严酷,对品种抗寒性要求严苛,甘蓝型冬油菜均难以越冬,白菜型冬油菜是在北方地区安全越冬的唯一类型。保证高越冬率是北方地区对冬油菜品种的基本要求,而品种抗寒性鉴定是选择强抗寒品种的基础[2]。对西葫芦、甜瓜、茄子、杧果、水稻、小麦、大白菜等研究表明,品种的抗寒性与生理生化指标间存在一定的关系[3-9]。胡胜武等[10]指出甘蓝型冬油菜叶片组织中的细胞汁液浓度、游离脯氨酸含量与抗寒性密切相关。朱慧霞等[11]研究表明,白菜型冬油菜的抗寒性与叶片中过氧化物酶活性、可溶性蛋白含量、可溶性糖含量关系密切。蒲媛媛等[12]研究指出,抗寒性强的油菜品种在低温下能保持较高的SOD活性、CAT活性以及高的可溶性蛋白含量和较低的MDA含量。王晓军等[13]研究表明,野生红三叶由于根系存在优势(根系发达、根冠比较大),因此抗寒性优于进口红三叶。然而关于白菜型与甘蓝型冬油菜抗寒机理差异研究的报道较少。因此,本研究选用8个抗寒性不同的冬油菜品种,分析比较其植物学形态、干物质积累量、生理生化指标间的差异,利用隶属函数法及相关性分析对其进行抗寒性综合评价,以探究白菜型与甘蓝型冬油菜抗寒机理差异的原因,为北方白菜型与甘蓝型冬油菜抗寒性研究提供理论依据。
1材料和方法
1.1试验材料
本试验分别以8个不同抗寒等级的冬油菜品种为材料,品种特性如表1。
1.2试验设计
本试验采用大田试验与盆栽试验相结合的方法,大田试验种植于甘肃省油菜工程技术研究中心兰州新区上川试验基地,年均气温10.3 ℃,最冷月平均最低温约-10 ℃,极端低温-28 ℃左右,冬季负积温-570 ℃;年平均日照时数为2 446 h,无霜期为140 d;年降水量 327 mm。试验于2013年8月25日播种,行距20 cm,株距7~8 cm,播前施尿素150 kg/hm2、磷酸二铵150 kg/hm2作为基肥,出苗后及时间苗、定苗,其他管理同大田生产。试验小区 2 m2(2 m×1 m),重复3次,随机区组排列。待幼苗生长至5~6片真叶时,统计其植物学形态,测定干物质积累量。
盆栽试验于2013年10月13日种植于甘肃农业大学校园温室大棚中,播种于营养钵中,营养钵高16 cm,直径12 cm。待幼苗生长至5~6片真叶时,放入低温试验箱,按24 ℃→10 ℃→5 ℃→0 ℃→-5 ℃→-10 ℃各48 h依次降温后取样测定生理生化指标。

表1 品种名称与来源
1.3测定指标与方法
1.3.1越冬率及植物学形态统计枯叶期前(2013年11月16日)记载越冬前基本苗数,翌年返青后(2014年3月18日)调查存活苗数,越冬率=越冬后苗数/冬前苗数×100%。越冬前油菜生长至5~6片真叶时,每品种随机选取5株,观察记载其植物学形态特征,根据《油菜种质资源描述规范和数据标准》[14]对其赋值。
1.3.2干物质积累的测定冬前气温下降至0 ℃时(11月3日),随机取5株发育良好的健壮株,整株采挖带回实验室冲洗干净,用吸水纸吸干,室温晾干测定根长、根颈直径,称量地上部鲜质量、地下部鲜质量;然后将叶片和根部放入牛皮纸袋中于90 ℃烘箱直至烘干,称其干质量。根/冠=地下部鲜质量(干质量)/地上部鲜质量(干质量)。
1.3.3生理生化特性测定超氧化物歧化酶(SOD)活性测定采用氮蓝四唑(NBT)光还原法[15];过氧化物酶(POD)活性测定采用愈创木酚比色法[15];过氧化氢酶(CAT)活性测定采用紫外吸收法[15];丙二醛(MDA)含量的测定采用硫代巴比妥酸法[15];可溶性蛋白含量测定采用考马斯亮蓝G250法[15];可溶性糖含量测定采用蒽酮显色法[15]。
1.4隶属函数分析
利用隶属函数法对各品种进行抗寒性综合评价[16-17],其公式为:
Uij=(Xij-Xjmin)/(Xjmax-Xjmin) 正相关
Uij=1-(Xij-Xjmin)/(Xjmax-Xjmin) 负相关
式中:Uij表示i品种j指标的抗寒隶属函数值;Xij表示i品种j指标的测定值;Xjmin表示所有品种j指标的最小值;Xjmax表示所有品种j指标的最大值;i表示某个品种;j表示某项指标。
1.5数据处理
利用Excel 2010 和SPSS 19.0 统计分析软件进行数据分析[18]。
2结果与分析
2.1白菜型冬油菜与甘蓝型冬油菜越冬率比较
白菜型冬油菜与甘蓝型冬油菜越冬率差异显著(图1),白菜型冬油菜陇油系列的越冬率高于天油2号,其中陇油7号最高,为94.50%;美切实和Vision越冬率低于40%,农林42号和黔油331号越冬率为0。
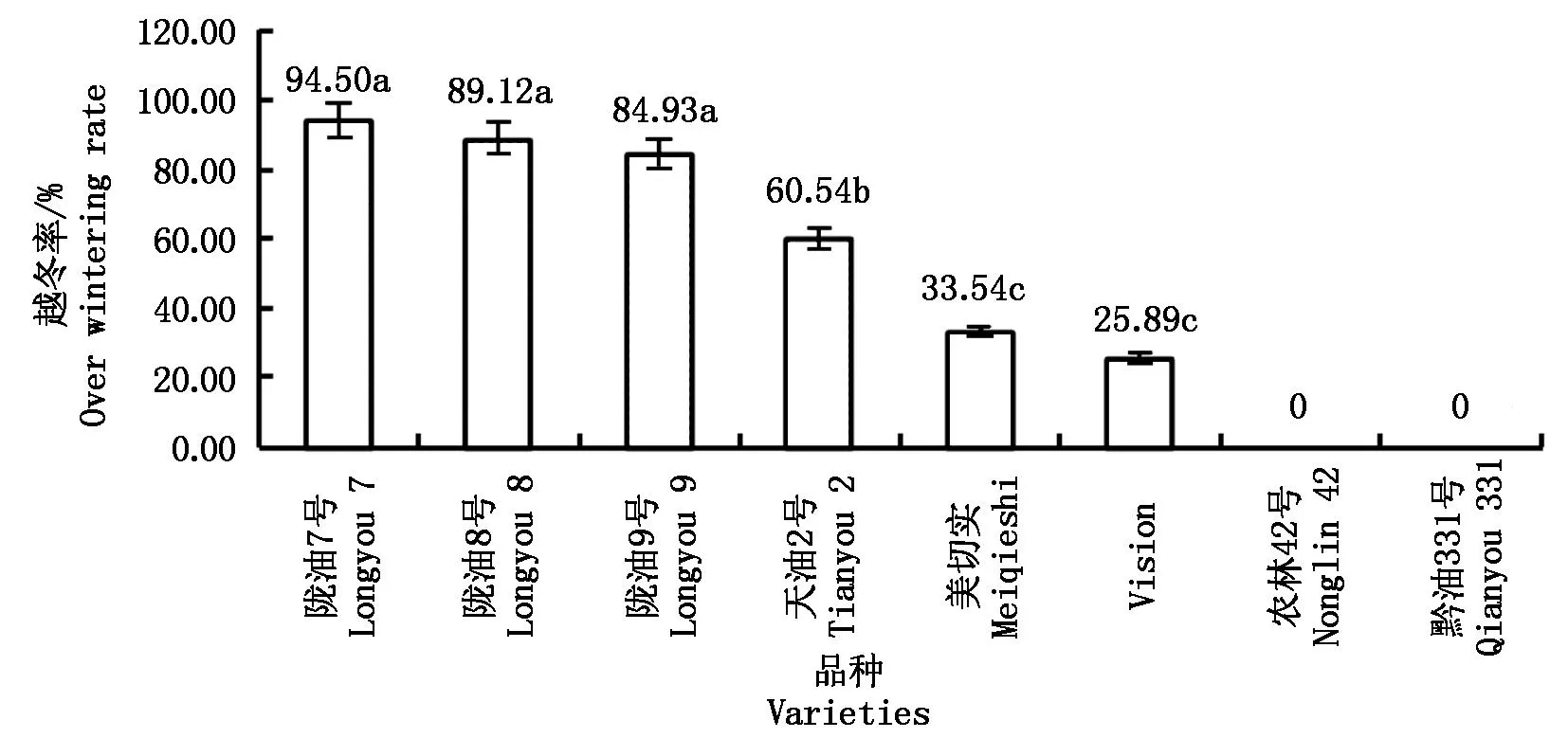
不同字母表示品种间在0.05水平差异显著。表3-9同。
2.2白菜型与甘蓝型冬油菜植物学形态比较
白菜型冬油菜与甘蓝型冬油菜在植物学形态上有较大差异(表2)。白菜型冬油菜苗期生长点洼陷,生长习性为匍匐生长;甘蓝型冬油菜苗期生长点凸起,生长习性为直立生长。
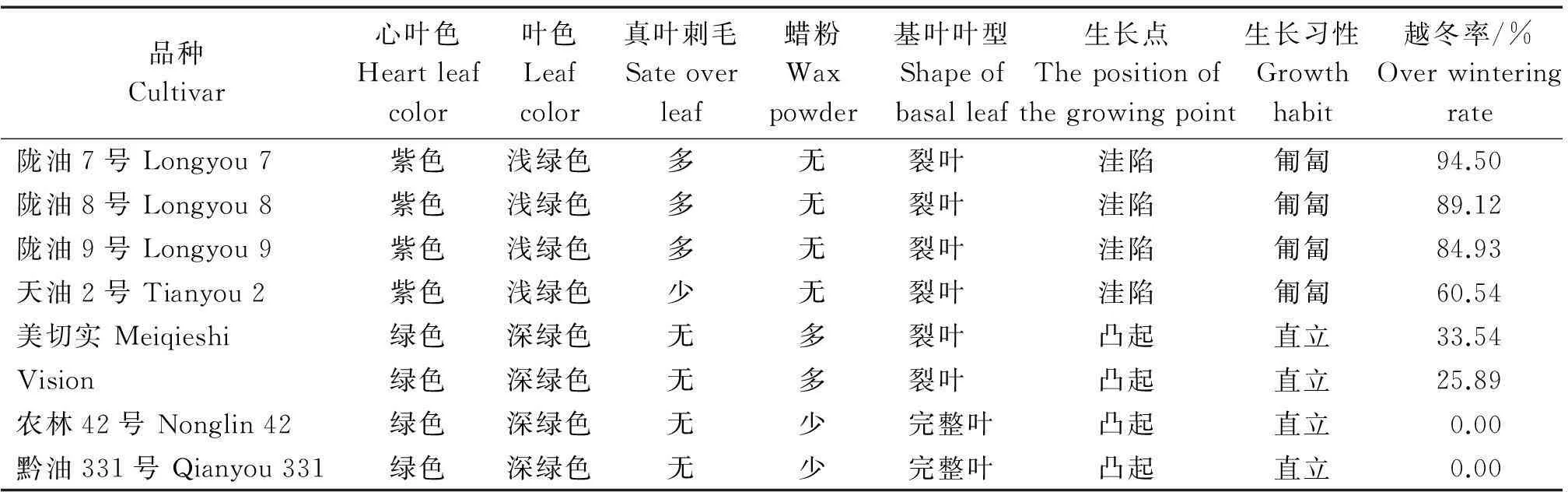
表2 白菜型冬油菜与甘蓝型冬油菜植物学形态比较
2.3白菜型与甘蓝型冬油菜干物质积累比较
白菜型冬油菜地下部鲜质量平均为16.511 g,地下部干质量平均为3.478 g,甘蓝型冬油菜地下部鲜质量平均为4.913 g,地下部干质量平均为0.958g,白菜型冬油菜地下部鲜质量与地下部干质量较甘蓝型冬油菜平均增加了236.1%,263.0%,甘蓝型冬油菜地上部鲜质量与地上部干质量较白菜型冬油菜平均增加了82.1%,98.0%;陇油7号地下部鲜质量、根/冠鲜质量、根/冠干质量、根颈直径分别较天油2号增加了96.0%,67.3%,60.8%,43.9%(表3、图2)。
2.4低温胁迫下白菜型冬油菜与甘蓝型冬油菜保护性酶活性比较
低温胁迫下白菜型冬油菜与甘蓝型冬油菜SOD活性均呈现先升高后降低趋势(表4),在-5 ℃时达到峰值,其中陇油7号最高为797.505 U/(g·min),较CK增加了10.7%,当温度由-5 ℃至-10 ℃过程中,甘蓝型冬油菜SOD酶活性下降趋势明显高于白菜型冬油菜。

图2 白菜型冬油菜与甘蓝型冬油菜干物质积累量比较
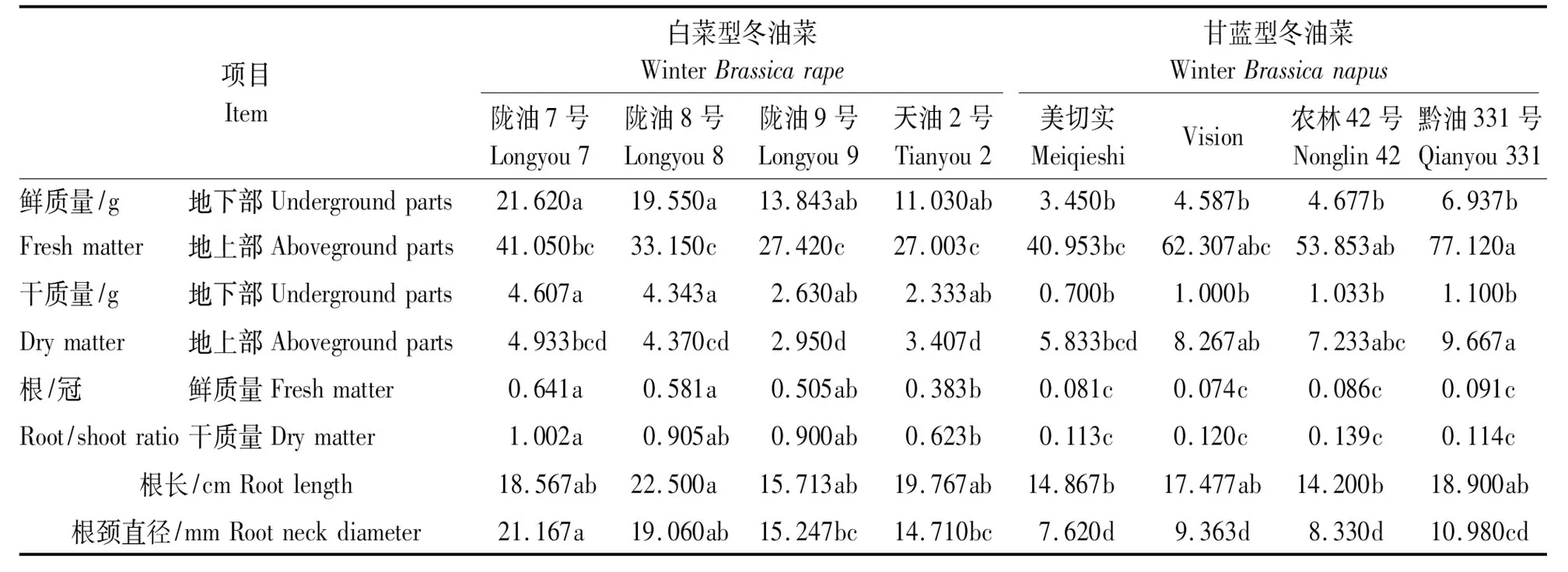
项目Item白菜型冬油菜WinterBrassicarape甘蓝型冬油菜WinterBrassicanapus陇油7号Longyou7陇油8号Longyou8陇油9号Longyou9天油2号Tianyou2美切实MeiqieshiVision农林42号Nonglin42黔油331号Qianyou331鲜质量/g地下部Undergroundparts21.620a19.550a13.843ab11.030ab3.450b4.587b4.677b6.937bFreshmatter地上部Abovegroundparts41.050bc33.150c27.420c27.003c40.953bc62.307abc53.853ab77.120a干质量/g地下部Undergroundparts4.607a4.343a2.630ab2.333ab0.700b1.000b1.033b1.100bDrymatter地上部Abovegroundparts4.933bcd4.370cd2.950d3.407d5.833bcd8.267ab7.233abc9.667a根/冠鲜质量Freshmatter0.641a0.581a0.505ab0.383b0.081c0.074c0.086c0.091cRoot/shootra-tio干质量Drymatter1.002a0.905ab0.900ab0.623b0.113c0.120c0.139c0.114c根长/cmRootlength18.567ab22.500a15.713ab19.767ab14.867b 17.477ab14.200b 18.900ab根颈直径/mmRootneckdiameter21.167a19.060ab15.247bc14.710bc7.620d9.363d8.330d10.980cd
温度由24 ℃下降至5 ℃时,白菜型冬油菜CAT活性的上升趋势比甘蓝型冬油菜明显(表5),温度继续降低至0 ℃时,各品种CAT活性达到最大值,陇油7号较CK增加了24.7%,白菜型冬油菜较甘蓝型冬油菜平均增加了38.2%,温度低于0 ℃,各品种CAT活性均下降,甘蓝型冬油菜下降趋势更明显。
随着温度由24 ℃逐渐降低至0 ℃时,POD活性逐渐升高,白菜型冬油菜增加趋势更明显(表6),当温度为10 ℃时品种间无明显差异,但当温度降到5 ℃时,POD活性升高趋势明显,尤其当温度降到0 ℃时,POD活性最高,陇油7号较CK增加了28.6%,白菜型冬油菜较甘蓝型冬油菜平均增加了40.2%,当温度继续降低时,甘蓝型冬油菜POD活性平均较白菜型冬油菜明显降低。

表4 低温胁迫下白菜型冬油菜与甘蓝型冬油菜SOD活性比较
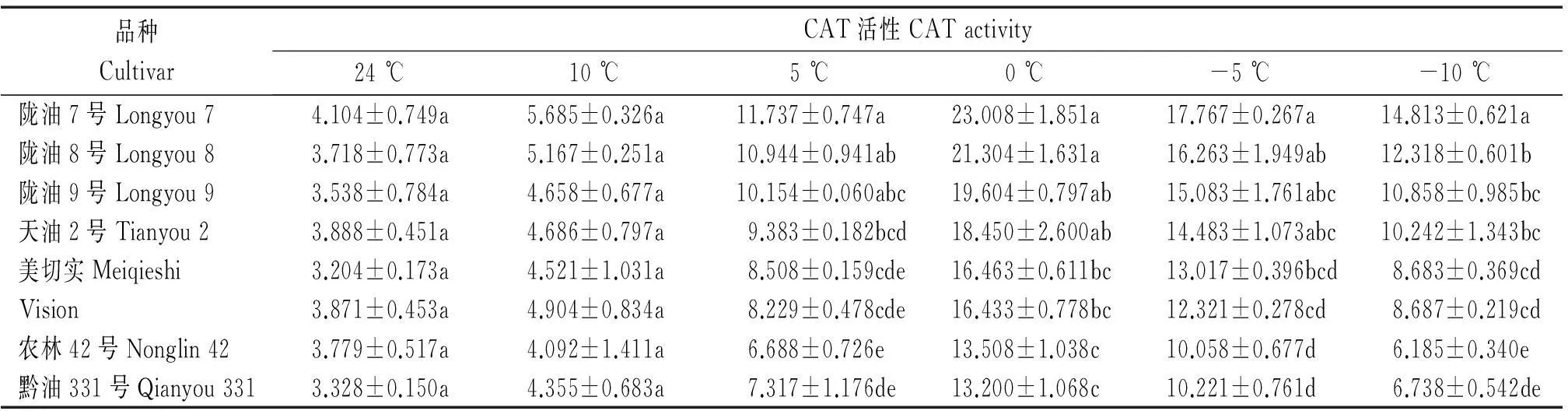
表5 低温胁迫下白菜型冬油菜与甘蓝型冬油菜CAT活性比较
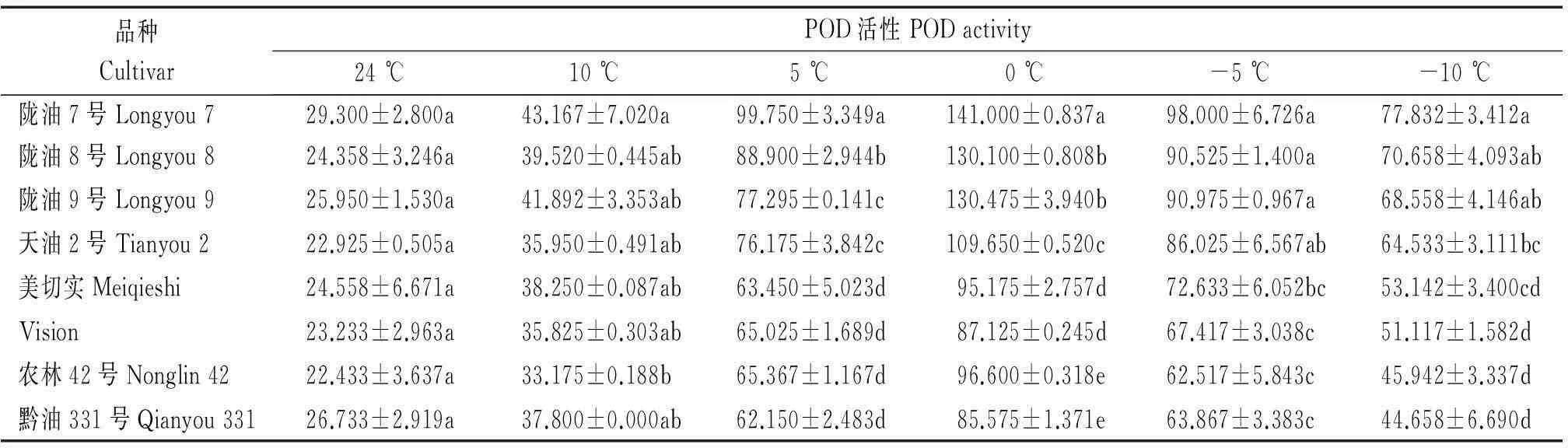
表6 低温胁迫下白菜型冬油菜与甘蓝型冬油菜POD活性比较
2.5低温胁迫下白菜型冬油菜与甘蓝型冬油菜可溶性蛋白含量比较
随着温度的降低SP含量呈先升高后降低再升高的趋势(表7),温度从24 ℃下降到0 ℃时,各品种的SP含量均上升,但白菜型冬油菜上升趋势更明显,在0 ℃时,各品种SP含量达到最大值,白菜型冬油菜较甘蓝型冬油菜平均增加了32.3%,温度继续下降到-5 ℃时,SP含量呈现下降趋势,温度降低到-10 ℃时,各品种SP含量明显增加,且白菜型冬油菜SP含量始终高于甘蓝型冬油菜。
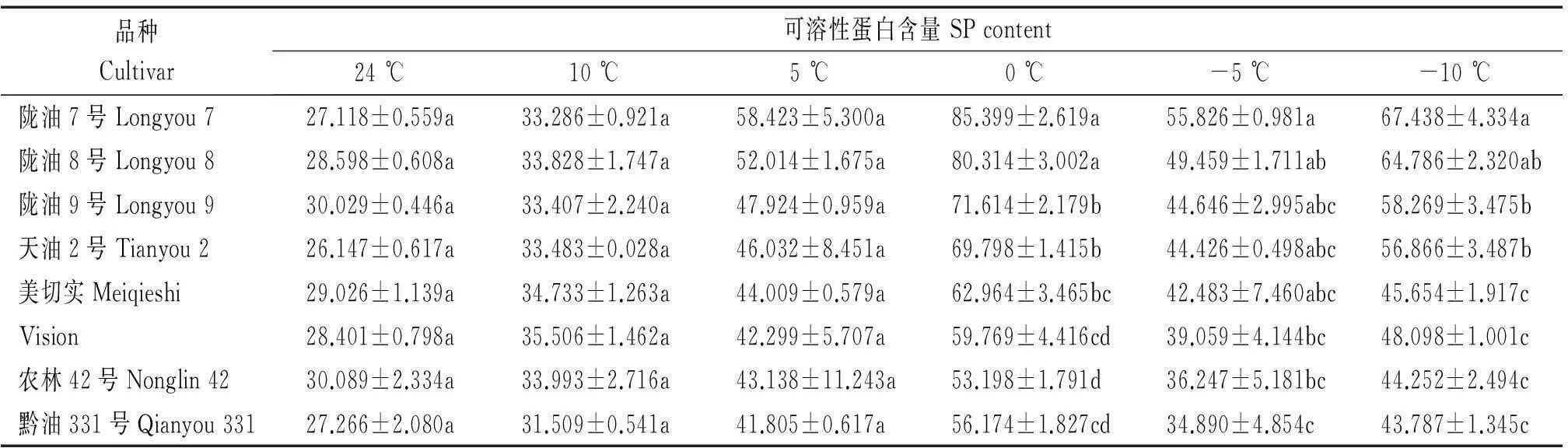
表7 低温胁迫下白菜型冬油菜与甘蓝型冬油菜可溶性蛋白含量比较
2.6低温胁迫下白菜型冬油菜与甘蓝型冬油菜可溶性糖含量比较
当温度由24 ℃降低到10 ℃时,不同类型冬油菜SS含量均缓慢增加(表8),温度降低至5 ℃时,各品种SS含量明显增加,白菜型冬油菜较甘蓝型冬油菜平均增加了31.3%,当温度降低到0 ℃时,SS含量均有所下降,甘蓝型冬油菜比白菜型冬油菜下降趋势更明显,随着温度继续降低,各品种冬油菜SS含量又开始上升,尤其是-10 ℃白菜型冬油菜的SS含量较甘蓝型冬油菜平均增加了71.4%。

表8 低温胁迫下白菜型冬油菜与甘蓝型冬油菜可溶性糖含量比较
2.7低温胁迫下白菜型冬油菜与甘蓝型冬油菜丙二醛含量比较
随着温度的降低,各品种MDA含量呈上升趋势(表9),温度由24 ℃降低到5 ℃,各品种MDA含量均上升,品种间无显著差异,当温度由0 ℃降低至-10 ℃,各品种MDA含量均上升,甘蓝型冬油菜MDA含量的上升趋势比白菜型冬油菜更明显,特别是在-10 ℃时,甘蓝型冬油菜MDA含量较白菜型冬油菜平均增加了52.8%。
2.8相关性分析
相关性分析结果得到,生长点、根/冠鲜质量、根/冠干质量、根颈直径与越冬率呈极显著正相关,相关系数分别为0.923,0.956,0.946,0.890,地下部鲜质量和地下部干质量与越冬率呈显著正相关,而生长习性、地上部鲜质量、地上部干质量与越冬率呈极显著负相关(表10)。
2.9隶属函数分析
植物抗寒性是由多个因素控制的复杂的生理生化过程,隶属函数法可以综合并准确说明植物的抗寒性强弱。本试验利用隶属函数法结合各项指标进行抗寒性综合评价。利用白菜型冬油菜的抗寒等级划分并结合表11各品种的平均隶属度,将试验材料分为6级:0.80~1.00为超强抗寒,定为Ⅰ级;0.70~0.79为强抗寒,定为Ⅱ级;0.60~0.69为抗寒,定为Ⅲ级;0.40~0.59为耐寒,定为Ⅳ级;0.21~0.40为不抗寒,定为Ⅴ级;0~0.20为极不抗寒,定为Ⅵ级。从该分级来看,陇油7号属超强抗寒品种,陇油8号属强抗寒品种,陇油9号属抗寒品种,天油2号属耐寒品种,美切实和Vision为不抗寒品种,农林42号和黔油331号为极不抗寒品种,完全符合实际越冬率高低情况,说明该分级正确。
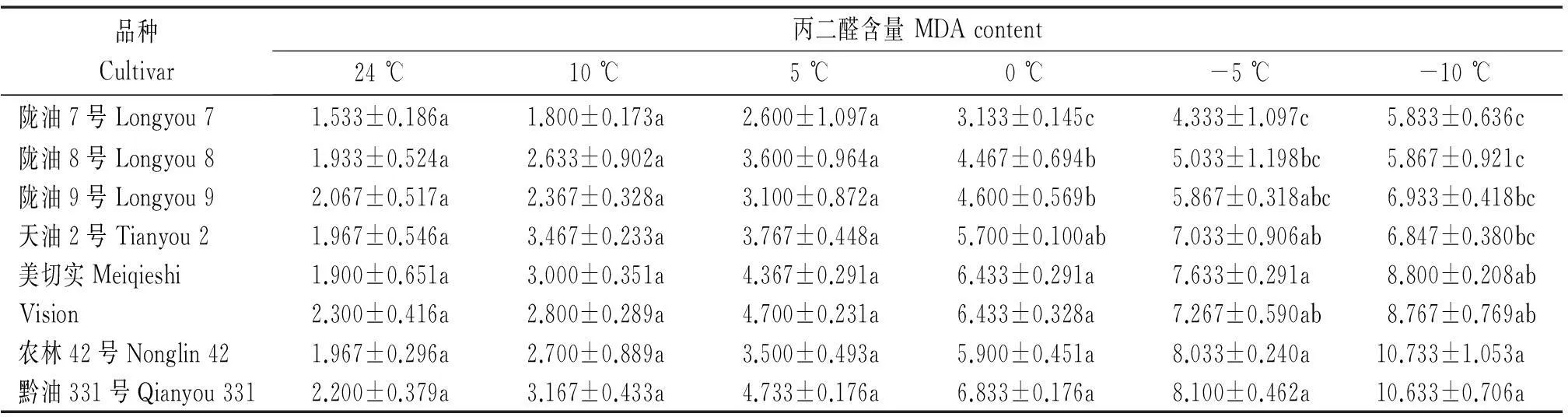
表9 低温胁迫下白菜型冬油菜与甘蓝型冬油菜丙二醛含量比较
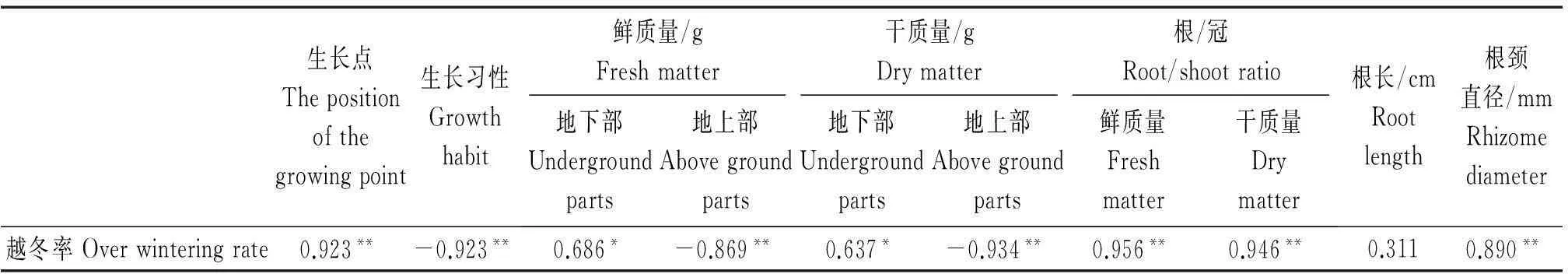
表10 相关性分析
注:*.相关系数达0.05水平显著;**.相关系数达0.01水平显著。
Note:*.The correlation coefficient of 0.05 significant level;**.Correlation coefficient of 0.01 significant level.
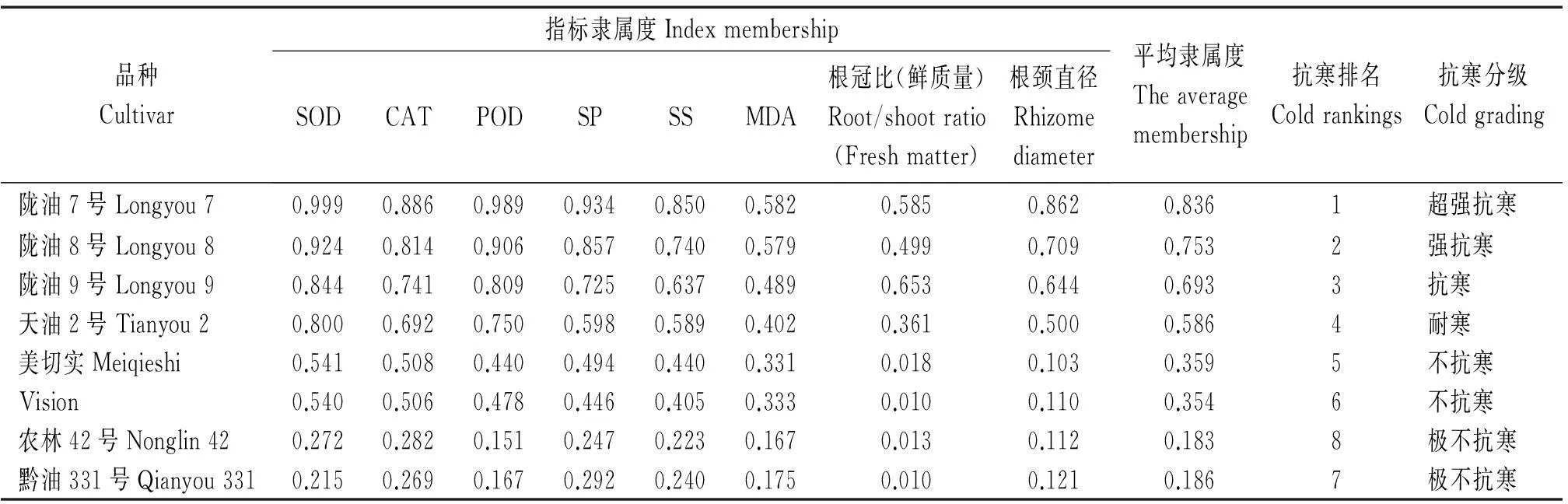
表11 隶属函数分析
3讨论与结论
越冬率的高低决定冬油菜抗寒性的强弱,而品种越冬率之所以高表现在植物学形态上苗期生长点洼陷,幼苗匍匐生长,相比较甘蓝型冬油菜更有利于抵御极端低温天气带来的损伤和死亡,甘蓝型冬油菜生长点突出,幼苗直立生长,在冬季风力较大的天气茎秆受冻容易被折断。营养生长阶段白菜型冬油菜干物质积累主要集中在地下部分,有发达的根系,光合有机产物被优先分配运输到地下部,在越冬期间,冬油菜地上部死亡,维持越冬所需的代谢能量全部由地下部供给,强抗寒性品种充足的干物质积累为越冬消耗提供保障[19]。
生理水平上冬油菜利用保护性酶、调节性物质、丙二醛来提高自身抗寒性。冬油菜叶片遭受低温胁迫后,细胞内氧自由基增加,破坏细胞结构,严重时细胞失水过多导致叶片死亡,此时,保护性酶不断增加,及时有效清除细胞内氧自由基[20-21],缓解低温带来的伤害,可溶性蛋白不断增加,降低了细胞失水带来的伤害[22],Heber与Kacperska-Palacz等[23-24]指出冬小麦在低温下可溶性蛋白质含量与其抗寒性显著正相关。Li等[25]指出植物抗寒性与可溶性糖之间的平行关系,一般认为,抗寒性强的植物积累更多的糖类物质,这些糖类物质使细胞渗透势降低,保护脱水后的蛋白质,还可促进脱落酸(ABA)的积累,间接诱导蛋白质的合成[26]。低温伤害迫使细胞膜发生膜脂过氧化反应,增加的MDA加速了细胞膜脂的过氧化,导致电解质渗漏及细胞水势下降,从而破坏体内的正常代谢[27-28],刘艳阳等[29]的试验结果已证明MDA的含量可以作为小麦抗寒性的鉴定指标,本研究结果也表明,低温胁迫下冬油菜叶片中的保护性酶、可溶性物质、MDA均升高,与前人在西葫芦、茄子和黄瓜等蔬菜作物研究上不完全一致;与杨宁宁等[30]和孟凡珍等[9]在冬油菜与大白菜上的研究结果相似,这可能是由于作物种类不同造成的。低温胁迫后CAT、POD、SP率先被启动,在0 ℃达到峰值,是一类与冷害相关的保护性物质,而SOD、SS分别在-5,-10 ℃启动达到最大,是抵御冻害相关的保护性物质。
利用相关性分析和隶属函数分析在多指标测定的基础上对材料进行综合评价,结果可靠[17]。张文娥等[16]与张朝阳等[31]研究表明,利用平均隶属度评价葡萄抗寒性与地被植物耐热性最为科学。植物的抗寒性是由多个因素控制的复杂的生理生化过程,所以单独的一个指标不能准确的反应品种的抗寒性强弱。目前,与植物抗寒相关基因和低温调控蛋白的研究已成为热门话题,因此,还需将抗寒基因、低温调控蛋白等分子水平上的理论与本研究相结合进行深入研究。
白菜型冬油菜比甘蓝型冬油菜在形态学及生理水平上都具有明显的优势,形态学上的优势使其有利于抵御极端低温天气,提供维持越冬及冬后返青结实所需的代谢能量;生理水平上,低温胁迫后保护性酶活性、调节性物质含量增加,能够有效地保护细胞膜结构,MDA含量减少,可以缓解低温对冬油菜叶片的伤害,从而保证高越冬率。
参考文献:
[1]孙万仓,马卫国,雷建民,等.冬油菜在西北旱寒区的适应性和北移的可行性研究[J].中国农业科学,2007,40(12):2716-2726.
[2]刘自刚,张长生,王月,等.不同生态区冬前低温下白菜型冬油菜不同抗寒品种(系)的比较[J].作物学报,2014,40(2): 346-354.
[3]马峰,张华锋,张怀友,等.低温胁迫对不同西葫芦品种抗寒性生理特性的影响[J].安徽农学通报,2015,21(13): 55-57.
[4]武雁军,刘建辉.低温胁迫对厚皮甜瓜幼苗抗寒性生理生化指标的影响[J].西北农林科技大学学报:自然科学版,2007,35(3):139-143.
[5]张继宁,阎世江.低温胁迫对茄子幼苗生理指标的影响[J].河北科技师范学院学报,2013,27(3): 1672-1675.
[6]佘文琴,刘星辉.低温胁迫下杧果叶片若干生理生化变化[J].福建农业大学学报,2001,30(2):180-184.
[7]朱珊,熊宏亮,黄仁良,等.低温胁迫对水稻生理指标的影响[J].江西农业学报,2013,25(7): 10-12.
[8]左芳,范宝莉.低温胁迫下小麦生理生化及蛋白组分变化研究[J].安徽农业科学,2009,37(4):1430-1431.
[9]孟凡珍,张振贤,于贤昌.田间低温胁迫对大白菜某些理化特性的影响研究[J].中国生态农业学报,2005,13(2):84-86.
[10]胡胜武,黄继英,徐爱遐,等.甘蓝型油菜抗寒性与某些渗透物质的关系[J].西北农业学报,1994,3(1):24-28.
[11]朱慧霞,孙万仓,邓斌,等.白菜型冬油菜品种的抗寒性及其生理生化特征[J].西北农业学报,2007,16(4):34-38.
[12]蒲媛媛,孙万仓.白菜型冬油菜抗寒性与生理生化特性关系[J].分子植物育种,2010,8(2):335-339.
[13]王晓军,于凤芝,矫健.旱地野生三叶草形态特征及抗寒性研究[J].黑龙江农业科学,2007(6):62-63.
[14]伍晓明,陈碧云,陆光远.油菜种质资源描述规范和数据标准[M].北京:中国农业出版社,2007.
[15]邹琦.植物生理学实验指导[M].北京:中国农业出版社,2000.
[16]张文娥,王飞,潘学军.应用隶属函数法综合评价葡萄种间抗寒性[J].果树学报,2007,24(6):849-853.
[17]玉苏甫·阿不力提甫,阿依古丽·铁木儿,帕提曼·阿布都热合曼,等.利用隶属函数法综合评价梨砧木抗寒性[J].中国农业大学学报,2014,19(3): 121-129.
[18]徐向宏,何明珠.试验设计与Design-Expert、SPSS应用[M].北京:科学出版社,2010.
[19]王晓楠,付连双,李卓夫,等.低温驯化及封冻后不同抗寒性小麦品种的形态建成及生理基础分析[J].作物学报,2009,35(7):1313-1319.
[20]南丽丽,师尚礼,陈建纲,等.不同根型苜蓿根系对低温胁迫的响应及其抗寒性评价[J].中国生态农业学报,2011,19(3): 619-625.
[21]刘海卿,武君艳,孙万仓,等.不同叶龄期叶面喷施ABA对北方白菜型冬油菜抗寒性的影响[J].草业学报,2015,24(9): 173-180.
[22]姜丽娜,张黛静,宋飞,等.不同品种小麦叶片对拔节期低温的生理响应及抗寒性评价[J].生态学报,2014,34(15): 4251-4261.
[23]Heber U.Cold-resistance of winter wheat[J].Planta,1959,54(1):34-37.
[24]Kacperska-Palacz A,Dlugokecka E,Breitenwald J,et al.Physiological mechanisms of frost tolerance:Possible role of protein in plant adaptation to cold[J].Biologia Plantarum,1977,19(1):10-17.
[25]Li P H,Skai A.Plant cold hardiness and freezing stress[M].London:Acadimic Press,1978:93.
[26]葛菁,庞磊,李叶云,等.茶树可溶性糖含量的HPLC-ELSD检测及其与茶树抗寒性的相关分析[J].安徽农业大学学报,2013,40(3): 470-473.
[27] 黄敏,陈杰忠.果树抗寒性研究进展[J].亚热带植物科学,2011,40(1):80-84.
[28]张东昱,盖玥,牛俊义,等.白菜型油菜抗寒生理生化特性动态研究[J].甘肃农业大学学报,2011,46(3):43-48.
[29]刘艳阳,李俊周,陈磊,等.低温胁迫对小麦叶片细胞膜脂质过氧化产物及相关酶活性的影响[J].麦类作物学报,2006,26(4):70-73.
[30]杨宁宁,孙万仓.北方冬油菜抗寒性的形态与生理机制[J].中国农业科学,2014,47(3):452-461.
[31]张朝阳,许桂芳.利用隶属函数法对 4 种地被植物的耐热性综合评价[J].草业科学,2009,26(2):57-60.
Study of Difference in Mechanism of Cold Resistance of Winter Rapeseed ofBrassicarapeandBrassicanapus
MA Li,SUN Wancang,LIU Zigang,ZHAO Yanning,YANG Gang,LIU Haiqing,WU Junyan,FANG Yan,LI Xuecai,LIU Linbo,QIAN Wu,HOU Xianfei
(Rapeseed Engineering Research Center of Gansu Province,Gansu Key Laboratory of Crop Improvement and Germplasm Enhancement,Key Laboratory of Arid Land Crop Science in Gansu Province,College of Agronomy,Gansu Agricultural University,Lanzhou730070,China)
Abstract:To explore the reasons for difference in mechanism of cold resistance of winter rapeseed of Brassica rape and Brassica napus.The method of combining field and pot culture was adopted,and using 8 different cold resistance of winter rapeseed varieties as materials,before wintering period with 5 to 6 true leaves,field experiments researched botanical morphology and accumulation of dry matter,the physiological characteristics were measured by pot experiment after the treatment of 24 ℃→10 ℃→5 ℃→0 ℃→-5 ℃→-10 ℃ 48 h.The results showed that winter Brassica rape was creeping growth,growth point was under the ground,accumulation of dry matter was mainly concentrated in the underground part,such as fresh matter of underground parts and dry matter of underground parts of winter Brassica rape had increased,which average increased 236.1%,263.0% than winter Brassica napus respectively.It was showed that the photosynthetic organic product of strong cold resistant varieties was preferentially allocated to the underground part during the vegetative growth stage,to establish a large root system to provide metabolic energy for the safe winter.With the change of temperature,the physiological and biochemical indexes of different varieties was vary greatly,SOD activity of Longyou 7 increased 10.7% than CK at low temperature of-5 ℃,CAT and POD activity of Longyou 7 increased 24.7%,28.6% than CK respectively at 0 ℃,SP content of winter Brassica rape average increased 32.3% than winter Brassica napus at 0 ℃,SS content of winter Brassica rape average increased 71.4% than winter Brassica napus,and MDA content of winter Brassica napus average increased 52.8% than winter Brassica rape at low temperature of-10 ℃,this indicated that the strong cold resistant varieties could protect themselves from damage of low temperature condition,the CAT,POD,SP were protective substance of against chilling injury,but SOD,SS were protective substance of against freezing.Therefore,the winter Brassica rape had obvious advantages in the morphology and physiological level than winter Brassica napus,the morphological advantages was that to withstand cold weather of extreme,to provide metabolic energy of during the wintering period and regreen next year;At physiological levels,the activities of protective enzyme and content of osmotic regulation of winter Brassica rape increased significantly at low temperature stress to protect the structure of cell membrane,and the accumulation of MDA was decreased,to alleviate hurt of winter rapeseed leaves at low temperature,so the over wintering was ensured highly,it was important to provide a theoretical basis for the research on the cold resistance of winter rapeseed of Brassica rape and Brassica napus in the North of China.
Key words:Brassica rape;Brassica napus;Winter rapeseed;Cold resistance;Morphological characteristic;Dry matter accumulation;Physiological and biochemical;Membership function
doi:10.7668/hbnxb.2016.01.024
中图分类号:S565.01
文献标识码:A
文章编号:1000-7091(2016)01-0147-08
作者简介:马骊(1991-),男,甘肃临泽人,在读硕士,主要从事作物遗传育种研究。通讯作者:孙万仓(1957-),男,甘肃会宁人,教授,博士,主要从事油菜育种及十字花科种质资源研究。
基金项目:国家“863”高技术研究发展计划项目(2011AA10A104);国家自然科学基金项目(30960199);国家现代农业产业技术体系建设专项(CARS-13);甘肃农业大学盛彤笙科技创新基金项目(GSAU-STS-1428)
收稿日期:2015-10-14
猜你喜欢
今日农业(2022年1期)2022-11-16 21:20:05
电脑迷(2022年10期)2022-11-08 02:41:30
今日农业(2021年20期)2021-11-26 01:23:56
河北果树(2020年4期)2020-11-26 06:04:28
西藏农业科技(2019年3期)2019-11-04 00:35:14
西藏农业科技(2019年1期)2019-07-25 00:37:02
西藏农业科技(2018年4期)2018-04-25 06:39:28
西藏农业科技(2018年4期)2018-04-25 06:39:26
西南农业学报(2016年6期)2016-04-16 05:12:46
广西林业科学(2016年2期)2016-03-20 05:53:26
