
CO2浓度对细基江蓠繁枝变型生长和光合特性的影响
2016-03-04 06:13:54秦秀刚仲启铖来琦芳么宗利高鹏程
海洋渔业 2016年1期
秦秀刚,仲启铖,来琦芳,么宗利,周 凯,高鹏程,王 慧
(1.中国水产科学研究院东海水产研究所,中国水产科学研究院盐碱地渔业工程技术研究中心,上海 200090;2.上海海洋大学水产与生命学院,上海 201306)
CO2浓度对细基江蓠繁枝变型生长和光合特性的影响
秦秀刚1,2,仲启铖1,来琦芳1,么宗利1,周 凯1,高鹏程1,王 慧1
(1.中国水产科学研究院东海水产研究所,中国水产科学研究院盐碱地渔业工程技术研究中心,上海 200090;2.上海海洋大学水产与生命学院,上海 201306)
比较了低CO2浓度(充无CO2空气,low CO2,简称LC)、正常CO2浓度(充正常空气,normal CO2,简称NC)、高CO2浓度(充正常空气+0.2%CO2,high CO2,简称HC)3种条件下细基江蓠繁枝变型(Gracilaria tenuistipitatavar.liuiZhangetXia)的特定生长率、光合色素含量、光合放氧速率和叶绿素荧光参数。结果表明:培养6 d后,与NC组相比,LC组细基江蓠繁枝变型特定生长率(SGR)、100μmol·(m2·s)-1光强净光合放氧速率及PSII最大光量子产率(Fv/Fm)均显著降低(P<0.05);类胡萝卜素含量显著增加(P<0.05);而藻红素、叶绿素a含量、PSII实际光量子产率(Fv/Fo)、快速光响应曲线初始斜率(α)、最大电子传递效率(ETRmax)及半饱和光强(Ik)均无显著差异(P>0.05)。与NC组相比,HC组细基江蓠繁枝变型的SGR、Fv/Fm及α显著增加(P<0.05),藻红素含量显著下降(P<0.05),而叶绿素a、类胡萝卜素含量、600μmol·(m2·s)-1光强净光合放氧速率、Fv/Fo、α、ETRmax及Ik均无显著差异(P>0.05)。与LC组相比,HC组细基江蓠繁枝变型的100μmol·(m2·s)-1光强净光合放氧速率、SGR、Fv/Fm、Fv/Fo、α均显著增加(P<0.05),而藻红素、叶绿素a、类胡萝卜素含量却显著下降(P<0.05),600μmol·(m2·s)-1光强净光合放氧速率、ETRmax、Ik无显著变化(P>0.05)。对于细基江蓠繁枝变型,CO2浓度升高降低了光合色素含量,同时提高了光系统光能转换效率,而对藻体饱和光强下净光合放氧速率的促进作用不显著。但是CO2浓度升高却仍然可能通过降低无机碳利用的能量消耗及提高营养盐吸收利用而显著促进藻体的生长。比较不同CO2浓度下细基江蓠繁枝变型生长和光合特性的差异,有助于初步揭示其对大气CO2浓度变化的响应特征。
细基江蓠繁枝变型;CO2;特定生长率;光合色素;叶绿素荧光
由于人类活动的加剧,现有大气CO2浓度已经从工业革命前的280μL·L-1增加至379μL· L-1,而这一浓度在21世纪中后期有可能会加倍[1-2],并且继续以每年3.3%的速度上升[3]。由于大气CO2浓度的增加,海水中溶解的CO2浓度也逐步上升[4]。
大型海藻是海洋生态系统生产力的重要组成部分,占总初级生产力10%左右[5],其固碳效率是陆生植物的10倍[6]。CO2是大型海藻重要的无机碳源,其浓度升高可能会对大型海藻的生长和光合作用产生重要影响。在生长特性方面,研究表明,CO2浓度升高会使龙须菜(Gracilarialemaneiformis)、羊栖菜(Hizikia fusiforme)生长速度加快[1,7],对石莼(Ulva lactuca)生长速率影响不显著[8],而对紫菜属中Pyropia leucostica的生长却有抑制作用[9]。由此可见,大型海藻对CO2浓度升高的响应存在明显的种间差异。在光合特征方面,研究表明,CO2浓度升高会使条斑紫菜(Pyropia yezoensis)的光合作用增强[10],使细基江蓠(Gracilaria tenuistipitata)[11]、智利江蓠(Gracilaria chilensis)光合作用下降,而对江蓠属Gracilariasp.光合作用的影响则不显著[12]。CO2浓度升高还会影响大型海藻的光合色素及其光化学系统。研究表明在高CO2浓度下,细基江蓠叶绿素和藻红素含量会下降50%[13],而莱茵衣藻(Chlamydomonas reinhardtiiDang)的叶绿素a、叶绿素 b和类胡萝卜素含量却显著升高[14];MERCADO等[9]认为,高浓度CO2会使紫菜属Pyropia leucostictePSII最大光量子产率和有效光量子产率下降,同时还会影响从PSII到PSI的电子传递速率。因此CO2浓度升高对大型海藻的影响是多方面的。
细基江蓠繁枝变型(Gracilaria tenuistipitata var.liuiZhangetXia)是我国热带、亚热带常见的大型经济海藻,其对温度、光照、盐度等环境因子均具有较大幅度的耐受性,在我国的广东、福建等地通过大规模人工栽培,已经具备比较高的产量[15],具有良好的经济价值。本研究以细基江蓠繁枝变型作为研究对象,通过测定和比较在低、正常、高CO2浓度3种条件下细基江蓠繁枝变型的特定生长率、光合色素含量、光合放氧速率及叶绿素荧光参数等,分析不同CO2浓度对其生长和光合特性的影响。本研究有助于初步揭示细基江蓠繁枝变型对大气中CO2浓度变化的生理生态响应,以期为大型海藻无机碳利用机制的研究提供参考。
1 材料与方法
1.1 实验材料
实验所用细基江蓠繁枝变型取自海南琼海养殖池塘。选取健康藻体,用软毛刷除去表面附着杂藻,而后用大量消毒海水冲洗,于5 L三角烧瓶中进行扩大培养。
1.2 扩大培养条件
盐度为20,温度为(25±1)℃,光照强度100 μmol·(m2·s)-1,光周期12 L∶12D,采用VSE培养液[16],pH为8.3±0.1,每2 d更换1次培养液,24 h持续充气。
实验开始前1周,取长势相似的健康藻体,置于装有200 mL培养液的250 mL高型烧杯中暂养备用。
1.3 实验设计
以充正常空气(CO2浓度为0.03%)为对照组(normal CO2,以下简称NC),分别设置低CO2浓度和高CO2浓度两个处理:低CO2浓度组(low CO2,以下简称LC)持续充入经0.5 mol·L-1NaOH溶液(以酚酞作为指示剂,当溶液颜色变为浅红色则更换)去除CO2的空气;高CO2浓度组(high CO2,以下简称HC)同时充入正常空气和0.2%CO2。
准确称取(1.00±0.01)g暂养藻体,分别置于装有200 mL培养液高型烧杯中,然后分别充入不同CO2浓度的空气(流量为25 L·h-1),其余实验条件同扩大培养条件。每组设置3个重复,培养周期为6 d。
1.4 指标测定
培养结束后,分别测定各组的特定生长率(SGR)、光合色素含量、净光合放氧速率和叶绿素荧光参数等,各指标测定方法如下:
1.4.1 特定生长率
每2 d称取各组藻体鲜生物量,SGR按下式计算:

式中,W0为实验开始时藻体鲜生物量(g),Wt为实验结束时藻体鲜生物量(g),t为培养天数(d)。
1.4.2 光合色素含量
叶绿素a和类胡萝卜素含量参照DERE等[17]方法测定,藻红素含量参照达维斯[18]方法测定。
1.4.3 净光合放氧速率
称取培养6 d后约(0.080±0.005)g的藻体,将其剪成长约1~5 mm片段,加入实验培养液,适应0.5~2 h,从而尽可能减少细胞损伤对光合放氧速率的影响,采用液相氧电极Chlorolab 2(Hansatech,英国)分别在光照强度为100μmol ·(m2·s)-1和600μmol·(m2·s)-1条件下测定净光合放氧速率,其中100μmol·(m2·s)-1为培养条件下的光照强度,600μmol·(m2·s)-1为仪器测得最大饱和光强。
1.4.4 叶绿素荧光参数
叶绿素荧光参数采用调制叶绿素荧光仪Phyto-PAM(WALZ,德国)测定,测量前将样品剪成长约1~5 mm片段,置于2 mL样品杯中,分别加入实验培养液。将待测样品暗适应10 min,打开光强为0.3μmol·(m2·s)-1红光,然后提供强度约为4 000μmol·(m2·s)-1饱和脉冲光,脉冲时间为0.8 s,测量过程由Phytowin软件控制,仪器获得参数包括最大光量子产率(Fv/Fm)、实际光量子产率(Fv/Fo)、快速光响应曲线初始斜率(α)、最大电子传递速率(ETRmax)和半饱和光强(Ik)。
1.5 数据统计分析
实验所得数据使用SPSS 13.0统计软件进行统计分析。使用One-way ANOVA方差分析对不同CO2浓度条件下江蓠的生长和光合特性的差异进行分析,若差异显著,则使用Duncan’s多重比较,进行组间两两差异分析,以P<0.05作为差异显著性水平。
2 结果与分析
2.1 CO2浓度对细基江蓠繁枝变型特定生长率的影响
不同CO2浓度对细基江蓠繁枝变型生长的影响如图1所示。在不同CO2浓度下,随着培养时间的延长,细基江蓠繁枝变型鲜生物量均逐渐增加。在LC组、NC组和HC组细基江蓠繁枝变型的特定生长率分别为9.4%、14.8%、20.3%,其中LC组的特定生长率显著低于NC(P<0.05),HC组的特定生长率显著高于NC组(P<0.05)。在本实验设置的CO2浓度范围内,CO2浓度越高,江蓠生长速率越快。

图1 不同CO2浓度条件下细基江蓠繁枝变型鲜生物量Fig.1 Fresh biomass of Gracilaria tenuistipitata var.liui Zhang et Xia under different CO2concentrations
2.2 CO2浓度对细基江蓠繁枝变型光合色素含量的影响
不同CO2浓度对细基江蓠繁枝变型光合色素含量的影响如图2所示。与NC组相比,LC组细基江蓠繁枝变型藻红素和叶绿素a含量无显著性差异(P>0.05),但类胡萝素含量显著增加(P<0.05);HC组叶绿素a和类胡萝卜素含量差异不显著(P>0.05),但藻红素含量显著降低(P<0.05)。HC组藻红素、叶绿素a和类胡萝素含量均显著低于LC组(P<0.05)。

图2 不同CO2浓度下细基江蓠繁枝变型藻体藻红素、叶绿素a和类胡萝卜素的含量Fig.2 The content of phycoerythrobilin,chlorophyll a and carotenoid in Gracilaria tenuistipitata var.liui Zhang et Xia under different CO2concentrations
2.3 CO2浓度对细基江蓠繁枝变型净光合放氧速率的影响
如图3所示,在100μmol·(m2·s)-1光照强度下,LC组净光合放氧速率为19.46μmol O2·(h·g)-1,显著低于NC组的31.24μmol O2·(h·g)-1和HC组的32.74μmol O2·(h·g)-1(P<0.05);而在600μmol·(m2·s)-1光照强度下,细基江蓠繁枝变型净光合放氧速率在3个CO2组之间不存在显著性差异(P>0.05)。
2.4 CO2浓度对细基江蓠繁枝变型叶绿素荧光特性的影响
2.4.1 最大光量子产率和实际光量子产率
如图4所示,不同CO2浓度下细基江蓠繁枝变型的最大光量子产率均具有显著性差异(P<0.05),HC组、NC组、LC组分别为0.54、0.56、0.63;对于细基江蓠繁枝变型的实际光量子产率,HC组、NC组、LC组分别为0.40、0.46、0.53,与NC组相比,LC组和HC组与之均不存在显著性差异(P>0.05),但LC组显著小于HC组(P<0.05)。
2.4.2 快速光响应曲线初始斜率α、最大电子传递速率和半饱和光强

图3 在100μmol·(m2·s)-1和600μmol·(m2·s)-1光强下不同CO2浓度组细基江蓠繁枝变型的净光合放氧速率Fig.3 Photosynthetic oxygen evolution rate of Gracilaria tenuistipitata var.liui Zhang et Xia under different CO2concentrations at the solar radiation of 100μmol·(m2·s)-1and 600μmol·(m2·s)-1
如图5所示,细基江蓠繁枝变型的最大电子传递速率(ETRmax)和半饱和光强(Ik)在3个CO2浓度下均不存在显著差异(P>0.05)。HC组江蓠初始斜率α显著高于LC组和NC组(P<0.05),分别为0.188、0.191、0.222,而LC组和NC组之间不存在显著差异(P>0.05)。
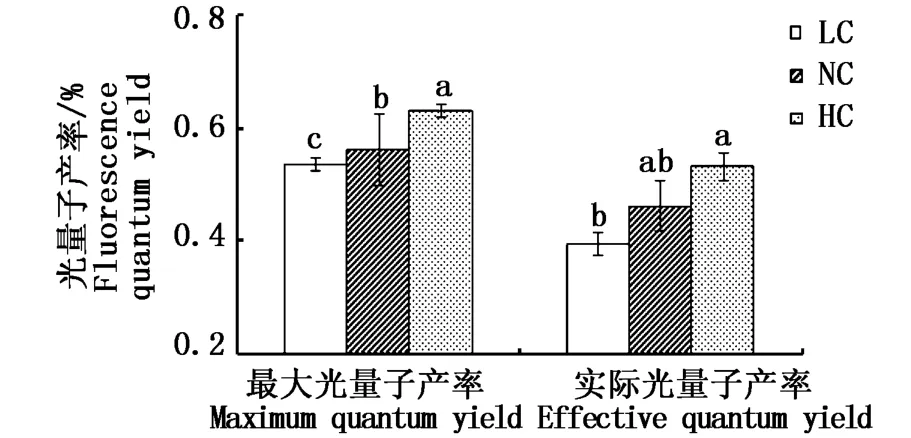
图4 不同CO2浓度下细基江蓠繁枝变型叶绿素PSII光量子产率Fig.4 Quantum yield of PSII of Gracilaria tenuistipitata var.liui Zhang et Xia under different CO2concentrations
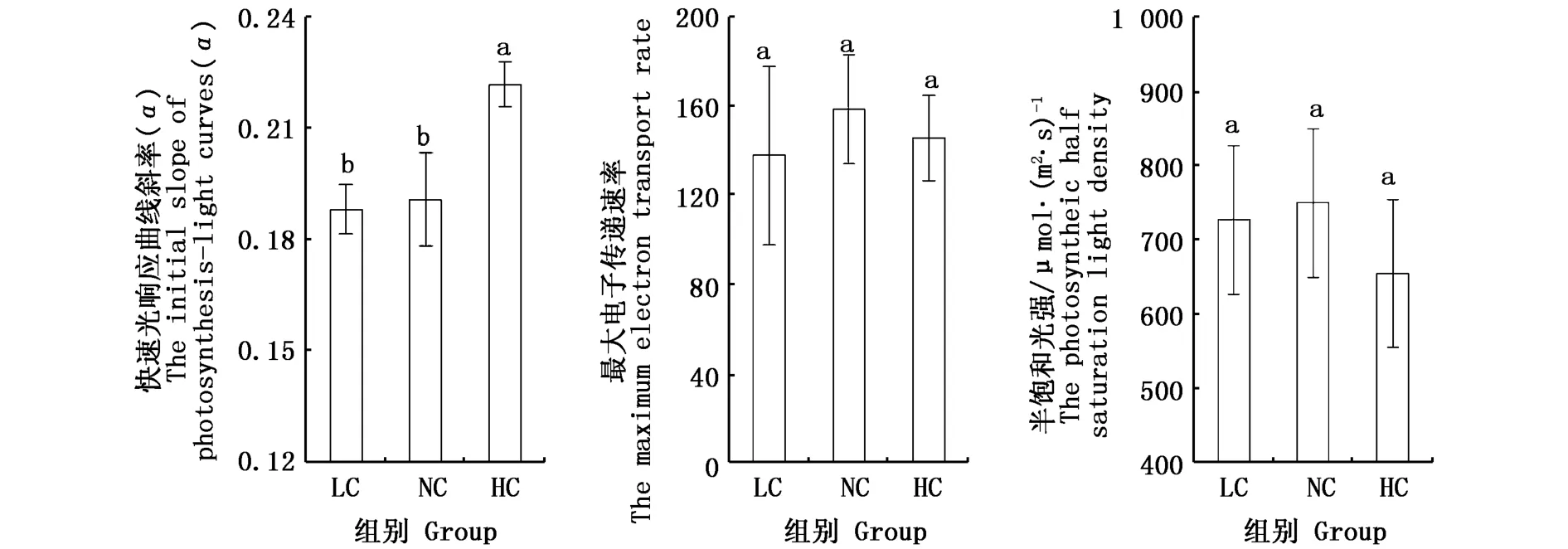
图5 不同CO2浓度下细基江蓠繁枝变型叶绿素快速光响应曲线初始斜率(α)、最大电子传递速率(ETRmax)、半饱和光强(Ik)Fig.5 The initial slope of photosynthesis-light curves(α),themaximum electron transport rate(ETRmax)of chlorophyll,the photosynthetic half saturation light density(Ik)of Gracilaria tenuistipitata var.liui Zhang et Xia under different CO2concentrations
3 讨论
3.1 CO2浓度对细基江蓠繁枝变型光合色素含量与光化学系统的影响
叶绿素a是藻类光合作用的主要色素,类胡萝卜素和藻红素则能够辅助叶绿素捕获光能,其中叶绿素a主要是吸收红光,而藻红素是红藻、蓝藻、隐藻和部分甲藻特有的捕光色素蛋白[19]。本研究中,从LC组、NC组到HC组,总体上看随着CO2浓度升高,细基江蓠繁枝变型藻体的藻红素、叶绿素a和类胡萝卜素含量呈下降趋势,其中HC组藻体的3种光合色素含量均显著低于LC组,说明CO2浓度升高在一定程度上会导致光合色素含量的下降。GORDILLO等[20]在钝顶螺旋藻[Spirulina platensis(Arthrospira)]中同样发现高CO2浓度会使光合色素含量下降。细基江蓠繁枝变型色素含量下降一方面可能是高CO2浓度促进江蓠生长后,单位重量中藻体的光合色素含量降低导致的;另一方面可能是江蓠应对环境CO2浓度升高的一种适应策略:CO2浓度升高后,细基江蓠繁枝变型可用于光合作用的底物浓度增加,短期内光合作用可能会提高。但光合作用增加会使大量可溶性碳水化合物在藻体内积累,可溶性碳水化合物会抑制藻体光合作用[21],且碳水化合物的积累需要消耗能量[22]。因此当CO2浓度持续升高时,细基江蓠繁枝变型通过降低光合色素含量而对光合作用实施负反馈,防止过多的无机碳积累。这种在高CO2下光合色素含量降低的效应还有可能是一种“色素节约(pigment economy)”现象,藻体只有通过更大程度地下调其光合中心色素含量,才能避免传递更多的光能到光系统中,避免对自身产生伤害[23]。
本研究中,与细基江蓠繁枝变型光合色素含量随着CO2浓度升高呈下降趋势相反,叶绿素荧光参数(Fv/Fm、Fv/Fo和快速光响应曲线初始斜率α)却随着CO2浓度升高呈现上升趋势。其中HC组藻体上述3个叶绿素荧光参数均显著高于LC组。PSII最大光化学量子产量反映PSII反应中心最大光能转换效率;Fv/Fo代表PSII的潜在活性[23],说明CO2浓度升高在一定程度上会显著提高PSII利用光能的效率和潜在活性,而快速光响应曲线初始斜率α增加也反映了CO2浓度升高增强了细基江蓠繁枝变型藻体对光能的利用能力。光系统光能转换效率的增加能在一定程度上补偿CO2浓度升高导致光合色素含量的下降,从而使光合作用速率保持相对稳定。
3.2 CO2浓度对细基江蓠繁枝变型光合放氧的影响
植物的光合作用包括光反应和碳反应(暗反应),是将光反应中获得的光合能量传递到碳反应,两者进行偶联的氧化还原反应[24]。CO2浓度的升高能提高部分藻类光合作用的速率[25],但本研究中发现细基江蓠繁枝变型净光合放氧速率却没有显著改变,这可能与CO2浓度升高导致光合色素含量减少和PSII光能利用效率上升有关,这两个过程的动态平衡使光合放氧速率略大于对照组,但未达到显著水平。此外,水体中CO2浓度的升高会降低细基江蓠Rubisco酶活性,使CO2羧化作用受到抑制[5],从而光合放氧速率受到影响;本研究测定的是CO2浓度升高后第六天的净光合放氧速率,CO2浓度增加短时期内会增强植物的光合作用[26-28],但随着细胞内O2含量上升,O2会对Rubisco-PCRC(光合碳还原循环)相关酶产生抑制作用[29],从而形成了O2对光合作用的反馈抑制。因此这两个潜在反应可能会抵消CO2作为底物增加时对光合速率的显著促进作用,从而在本实验条件下表现为不同CO2浓度下光合放氧速率变化差异不显著(P>0.05),这也是细基江蓠繁枝变型应对水环境中CO2浓度升高的适应响应。今后可以进一步监测CO2浓度升高后细基江蓠繁枝变型净光合放氧速率的变化规律。
3.3 CO2浓度对细基江蓠繁枝变型生长的影响
CO2浓度的升高对大型海藻生长的影响在不同种类之间存在很大差异,龙须菜、智利江蓠和羊栖菜的生长会被促进[1,7,12],而紫菜属Pyropia leucosticte[9]和Pyropia linearis[30]生长则会随着CO2浓度升高而受到抑制。本研究中,尽管CO2浓度升高没有显著提升饱和光强下细基江蓠繁枝变型的光合放氧速率,但对其生长的促进作用十分显著。除了对光合色素含量、光系统光能转换效率以及碳-氧平衡影响之外,CO2浓度变化还可能对细基江蓠繁枝变型无机碳的摄入方式产生影响。CO2和浓度的升高对海洋光合自养生物的生长有潜在的激发作用[31],细基江蓠繁枝变型对CO2和都具有很好的利用效果,但的摄入是一个耗能过程[19],CO2浓度升高,更多的无机碳通过直接扩散方式进入细胞,为藻体提供了充足的光合原料,减少了江蓠对水体中的利用所需要消耗的自身ATP,更多能量被节省下来用于生长。而在低CO2浓度下,藻体吸收利用所需的ATP要高于高CO2浓度[10],另外,在低CO2浓度下,藻体要分配更多能量去适应 pH降低的水体环境[32],因此,在低CO2浓度下藻体的生长速率会显著降低。此外CO2浓度升高到一定程度上还可以促进藻体对水体中无机营养盐的吸收和氮在体内的积累[29],高浓度CO2可能促进了细基江蓠繁枝变型对营养盐的均衡吸收,满足了细基江蓠繁枝变型生长所需的营养需求,从而促进其生长。
因此,CO2浓度升高对细基江蓠繁枝变型生长的促进是一个综合反应的体现。
致谢:感谢上海海洋大学何培民老师对本项研究提供的帮助。
[1] 徐军田,高坤山.二氧化碳和阳光紫外辐射对龙须菜生长和光合生理的影响[J].海洋学报,2010,32(5):144-151.
XU JT,GAO K S.The influence of carbon dioxide and solar UVR on the growth,photosynthesis and pigments contents ofGracilaria lemanei formis[J].ACTA Oceanologica Sinica,2010,32(5):144-151.
[2] IPCC.Climate change 2007:The physical Science Basis.Contribution of working group I to the forth assessment report of the intergovernmental panel on climate change[R].Cambridge:Cambridge University Press,2007:36.
[3] CANADELL JG,QUÉRÉC L,RAUPACH M R,et al.Contributions to accelerating atmospheric CO2growth from economic activity,carbon intensity,and efficiency of natural sinks[J].Proceedings of the National Academy of Sciences of the United States of America,2007,104(47):18866-18870.
[4] XU J,GAO K.Future CO2induced ocean acidificationmediates the physiological performance of a green tide alga[J].Plant Physiology,2012,160(4):1762-1769.
[5] 韩婷婷.大型海藻对不同CO2浓度的光合生理响应及其生态效应[D].青岛:中国科学院研究生院(海洋研究所),2013:1-12.
HAN T T.Photosynthetic physiological responses of macroalgae to different CO2concentrations and their ecological effects[D].Qingdao:The University of Chinese Academy of Sciences,2013:1-12.
[6] NAKANO S,CHANG K H,SHIJIMA A,et al.A usage of CO2hydrate:Convenientmethod to increase CO2concentration in culturing algae[J].Bioresource Technology,2014,172(9):444-448.
[7] ZOU D.Effects of elevated atmospheric CO2on growth,photosynthesis and nitrogen metabolism in the economic brown seaweed,Hizikia fusiforme(Sargassaceae,Phaeophyta)[J].Aquaculture,2005,250(3-4):726-735.
[8] 邹定辉,高坤山,阮祚禧.高CO2浓度对石莼光合作用及营养盐吸收的影响[J].青岛海洋大学学报,2001,31(6):877-882.
ZOU D H,GAO K S,RUAN Z X.Effects of elevated CO2concentration on photosynthesis and nutrients uptake ofUlva lactuca[J].Journal of Ocean University Of Qingdao,2001,31(6):877-882.
[9] MERCADO JM,JAVIER F,GORDILIO L,et al.Effects of different levels of CO2on photosynthesis and cell components of the red algaPorphyra leucostica[J].Journal of Applied Phycology,1999,11(5):455-461.
[10] GAO K,ARUGA Y,ASADA K,et al.Enhanced growth of the red algaPorphyra yezoensisUeda in high CO2concentrations[J].Journal of Applied Phycology,1991,3(4):355-362.
[11] GARCÍA SÁNCHEZ M J,FERNÁNDEZ JA,NIELL F X.Effect of inorganic carbon supply on the photosynthetic physiology ofGracilaria tenuistipitata[J].Plant,1994,194(1):55-61.
[12] GAO K,ARUGA Y,ASADA K,et al.Influence of enhanced CO2on growth and photosynthesis of the red algaeGracilariasp.andG.Chilensis[J].Journal of Applied Phycology,1993,5(6):563-571.
[13] 邹定辉,高坤山.高CO2浓度对大型海藻光合作用及有关过程的影响[J].生态学报,2002,22(10):1750-1757.
ZOU D H,GAO K S.Effects of elevated CO2concentration on the photosynthesis and relatedphysiological processes in marine macroalgae[J].ACTA Ecological Sinica,2002,22(10):1750-1757.
[14] 夏建荣.高浓度CO2对莱茵衣藻光系统II能量流和能量利用效率的影响[J].水生生物学报,2005,29(4):450-455.
XIA J R.Influences of exerna high CO2concentration on the energy fluxes and yield of pSII in the green algaChlamydomonas reinhardtii[J].Acta Hydrobiologica Sinica,2005,29(4):450-455.
[15] 陈昌生,章景荣.福建省细基江蓠繁枝变型人工栽培的探讨[J].集美大学学报(自然科学版),1999,4(2):65-71.
CHEN C S,ZHANG J R.A discussion on the techniques of artificial cultivation ofGracilaria tenuistipitatavar.liui in Fujian[J].Journal of Jimei University(Natural Science),1999,4(2):65-71.
[16] 蒋雯雯,田相利,董双林,等.营养盐浓度变动对菊花江蓠和细基江蓠繁枝变型生长、生化组成及生理的影响研究[J].中国海洋大学学报,2010,40(增刊):83-90.
JIANGW W,TIAN X L,DONG S L,et al.Effects of nutrient concentration changes on growth,biochemical compositions and physiology ofGracilarialichenoides and G.tenuistipitata var.liui[J].Periodical Of Ocean University Of China,2010,40(sup):83-90.
[17] DERE S,GÜNEST,SIVACIR.Spectrophotometric determination of chlorophyll-A,B and total carotenoid contents of some algae species using different solvents[J].Turkish Journal of Botany,1998(22):13-17.
[18] 达维斯C J.海洋植物学.厦门大学植物生态学研究室译[M].厦门:厦门大学出版社,1989:258.
DA W S C J.Marine Botany.Translated by Department of plant ecology,Xiamen University[M].Xiamen:Xiamen University press,1989:258.
[19] 李小伟,么宗利,周 凯,等.高HCO-3浓度对细基江蓠繁枝变型生长及生理生化特征的影响[J].中国水产科学,2013,20(4):843-850.
LIXW,YAO Z L,ZHOU K,et al.Effect of high HCO3-concentrations on the growth,physiology,and biochemistry ofGracilaria tenuistipitata[J],Journal of Fishery Sciences of China,2013,20(4):843-850.
[20] GORDILLO F JL,JIMÉNEZC,FIGUEROA F L,etal.Effects of increased atmospheric CO2and N supply on photosynthesis,growth and cell composition of the cyanobacteriumSpirulina platensis(Arthrospira)[J].Journal of Applied Phycology,1998,10(5):461-469.
[21] 徐军田.经济红藻龙须菜和海萝的光生物学研究[D].汕头:汕头大学,2008:39.
XU J T.Photobiological studies on the economic red macroaglae Gracilaria lemaneiformis and Gloiopeltisfurcata[D].Shantou:Shantou University,2008:39.
[22] RAVEN JA,GIORDANOM,BEARDALL J,etal.Algal evolution in relation to atmospheric CO2:carboxylases,carbon-concentrating mechanisms and carbon oxidation cycles[J].Philosophical Transactions of the Royal Society of London B:Biological Sciences,2012,367(1588):493-507.
[23] 郑 伟,钟志海,杨 梓,等.大气CO2增加对不同生长光强下龙须菜光合生理特性的影响[J].生态学报,2014,24(34):7293-7300.
ZHENGW,ZHONG Z H,YANG Z,et al.Effects of elevated CO2concentration on the photosynthetic physiological characteristics ofGracilaria lemaneiformisgrown under different light levels[J].Acta Ecologica Sinica,2014,34(24):7293-7299.
[24] 张绪成,于显枫,马一凡.施氮和大气CO2浓度升高对小麦旗叶光合电子传递和分配的影响[J].应用生态学报,2011,22(3):673-680.
ZHANG X C,YU X F,MA Y F.Effects of nitrogen application and elevated atmospheric CO2on electron transport and energy partitioning in flag leaf photosynthesis of wheat[J].Chinese Journal of Applied Ecology,2011,22(3):673-680.
[25] 杨 英.CO2浓度升高对微藻类生长和光合作用影响的研究[D].汕头:汕头大学,2001:5-6.
YANG Y.Studies on the effects of elevated CO2concentrations on the growth and photosynthesis of microalgae[D].Shantou:Shantou University,2001:5-6.
[26] 刘 露.三种大型海藻光合特性对海洋环境条件变化的响应[D].广州:华南理工大学,2013:4-5.
LIU L.Responses of photosynthetic characteristics to marine environmental changes in three macroalgae[D].Guangzhou:South China University of Technology,2013:4-5.
[27] ZOU D,GAO K,LUO H.Short-and long-term effects of elevated CO2on photosynthesis and respiration in the marine macroalgaHizikia fusiformis(Sargassaceae,phaeophyta)grown at low and high N supplies[J].Journal of Phycology,2011,47(1):87-97.
[28] 邓亚运,邹定辉.大气CO2浓度升高对不同氮生长条件下的两种大型海藻光合作用的影响[J].生态学杂志,2014,33(6):1520-1527.
DENG Y Y,ZOU D H.Effects of elevated atmospheric CO2on photosynthesis ofGracilaria lemaneiformisandUlva conglobatagrown at low and high N supplies[J].Chinese Journal of Ecology,2014,33(6):1520-1527.
[29] LOW-DÉCARIE E,JEWELL M D,FUSSMANN G F,et al.Long-term culture at elevated atmospheric CO2fails to evoke specific adaptation in seven freshwater phytoplankton species[J].Proceedings of the Royal Society of London B:Biological Sciences,2013,280(1754):159-169.
[30] ISRAEL A,KATZ S,DUBINSKY Z,et al.Photosynthetic inorganic carbon utilization and growth ofPorphyra linearis(Rhorophyta)[J].Journal of Applied Phycology,1999,11(5):447-453.
[31] JOHNSON V R,BROWNLEEC,RICKABY R EM,et al.Responses of marine benthic microalgae to elevated CO2[J].Marine Biology,2013,160(8):1813-1824.
[32] HOFMANN L C,STRAUB S,BISCHOF K.Elevated CO2levels affect the activity of nitrate reductase and carbonic anhydrase in the calcifying rhodophyteCorallina officinalis[J].Journal of Experimental Botany,2013,64(4):899-908.
Effect of CO2on grow th and photosynthetic characteristics of Gracilaria tenuistipitata var.liui Zhang et Xia
QIN Xiu-gang1,2,ZHONG Qi-cheng1,LAIQi-fang1,YAO Zong-li1,ZHOU Kai1,GAO Peng-cheng1,WANG Hui1
(1.Engineering Technology Research Center for Saline-alkalineWater Fisheries,East China Sea Fisheries Research Institute,Chinese Academy of Fishery Sciences,Shanghai200090,China;2.College of Fisheries and Life Science,Shanghai Ocean University,Shanghai 201306,China)
The increase of CO2concentration in the atmosphere can lead to the increase of dissolved CO2concentration in sea water accordingly.As the inorganic carbon source of photosynthesis for algae,the change of CO2concentration has important influences on the photosynthesis and growth of algae.Gracilaria tenuistipitatavar.liuiZhangetXia,an economic marine macroalgae used as abalone feed and agar raw material,has been widely farmed in the southeastern coast of China.The special growth rate(SGR),photosynthetic pigment content,photosynthetic rates and chlorophyll fluorescence parameters(the maximum quantum yield of PSII photochemistry(Fv/Fm),the effective quantum yield of PSII photochemistry(Fv/Fo),the initial slope of the rapid light curves(α),the maximum electron transport rate(ETRmax)and the photosynthetic half saturation light density(Ik)ofGracilaria tenuistipitatavar.liuiZhang et Xia were measured and compared under different CO2concentrations,to analyze the influence of different CO2concentrations on the growth and photosynthesis of this alga.Thematerials,collected from aquaculture ponds in Qionghaiwere exposed to three CO2concentration levels:low(pumping in CO2-removed air,LC),normal(pumping in ambient air,NC)and high(pumping in ambient air+0.2%CO2,HC).Three parallels were set at each level.(1.00±0.01)g algal thallus was put in each beaker with 24 h aeration.The cultivation conditionswere as follows:the salinity was 20,temperature(25±1)℃,light intensity 100μmol·(m2· s)-1,the ratio of light period and dark period was 12L∶12D,the cultivation water was prepared with VSE nutrient solution sea water and was changed every two days.The cultivation lasted for 6 days.Then the algalthalliwere taken out and dried by plotting paper.The fresh biomasses of the algae before and after the cultivation wereweighed to calculate the specific growth rate.The algal thalli in the similar partswere used to determine the content of chlorophylla,phycoerythrobilin and carotenoid.0.08 g algal thalliwasweighed,cut up into pieces tomeasure the oxygen evolution rate(i.e.photosynthetic rate).Other algal thalliwere used to measure the chlorophyll fluorescence parameters by Phyto-PAM.Results showed that:when thalli in LCwere compared to those in NC,the special growth rate(SGR),the net photosynthetic rate under 100μmol·(m2·s)-1and themaximum quantum yield of PSII photochemistry(Fv/Fm)decreased significantly(P<0.05),while the contents of carotenoid increased markedly(P<0.05).In contrast,no difference was observed for chlorophyll a,phycoerythrobilin,the effective quantum yield of PSIIphotochemistry(Fv/Fo),the initial slope of the rapid light curves(α),the maximum electron transport rate(ETRmax)or the photosynthetic half saturation light density(Ik)(P>0.05).When thalli in HC were compared to those in NC,SGR,Fv/Fmandαincreased significantly(P<0.05),the content of phycoerythrobilin decreased(P<0.05),while chlorophyll a,carotenoid,the net photosynthetic rate under 600μmol·(m2·s)-1had no difference(P>0.05).When thalli in HC were compared to LC,SGR,Fv/Fm,Fv/Foandαincreased significantly(P<0.05),photosynthetic pigments decreased markedly(P<0.05),while the net photosynthetic rate under 600μmol·(m2·s)-1,the maximum electron transport rate(ETRmax)or the photosynthetic half saturation intensity showed no difference(P>0.05).To sum up,forGracilaria tenuistipitatavar.liuiZhangetXia,high CO2concentration can reduce the photosynthetic pigment contents and motivate the conversion efficiency of light energy of photosynthetic system.However,high CO2concentration does not lead to a significant promotion on net photosynthetic rate under saturation light intensity.Even so,high CO2concentration may realize the promotion of the growth ofGracilaria tenuistipitatavar.liuiZhang et Xia by reducing the energy consumption of inorganic carbon utilization,aswell as improving the nutrient utilization.In this study,investigating the growth and photosynthetic characteristics ofGracilaria tenuistipitatavar.liuiZhangetXia under different CO2concentration levels can help understand the ecophysiological adaptive ability of this alga to potential CO2concentration changes.
Gracilaria tenuistipitatavar.liuiZhangetXia;CO2;SGR;photosynthetic pigments;chlorophyll fluorescence
S 917.3
A
1004-2490(2016)01-0057-09
2015-04-24
中国水产科学研究院基本科研业务费专项(2013A0801)
秦秀刚(1989-),男,硕士研究生,主要从事大型藻类固碳及盐碱地水产养殖研究。E-mail:qinxiugang@163.com
来琦芳,研究员。Tel:021-65684655,E-mail:laiqf@ecsf.ac.cn
猜你喜欢
小读者·阅世界(2024年4期)2024-04-29 10:53:45
娃娃乐园·综合智能(2022年9期)2022-08-16 02:00:08
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15 06:35:20
科学大众(2021年9期)2021-07-16 07:02:50
天然产物研究与开发(2018年9期)2018-10-08 03:25:48
小资CHIC!ELEGANCE(2018年24期)2018-08-13 03:31:56
科技创新与应用(2017年20期)2017-07-15 20:43:02
中国调味品(2017年2期)2017-03-20 16:18:21
网络空间安全(2016年3期)2016-06-15 20:27:07
现代食品(2016年24期)2016-04-28 08:12:04
