
NDRG2通过抑制β-catenin表达和入核调控乳腺癌细胞增殖
2016-02-09 06:02:39
中国癌症杂志 2016年12期
河北科技大学生物实验中心,河北 石家庄 050018
NDRG2通过抑制β-catenin表达和入核调控乳腺癌细胞增殖
周晓雷,朱重悦,张世光,周志艳,李海潮,邹 卫
河北科技大学生物实验中心,河北 石家庄 050018
背景与目的:乳腺癌是女性发病率最高的恶性肿瘤之一,肿瘤细胞的恶性增殖是造成患者死亡的重要原因。本研究利用具有相同遗传背景但不同增殖能力的乳腺癌细胞模型,研究N-myc下游调节基因2(N-myc downstream regulated gene 2,NDRG2)调控乳腺癌细胞增殖的作用及分子机制。方法:通过蛋白[质]印迹法(Western blot)检测MCF-7/LM-MCF-7细胞中NDRG2蛋白表达水平;构建NDRG2真核表达载体及siRNA干扰片段,通过转染上调或沉默NDRG2表达,流式细胞实验检测细胞增殖能力,Western blot检测β-连环蛋白(β-catenin)表达,免疫荧光染色检测β-catenin细胞定位;共转染MCF-7细胞NDRG2 siRNA和pCMV-Tcfδ,流式细胞实验检测细胞增殖能力。结果:NDRG2的蛋白表达水平同乳腺癌细胞增殖能力负相关;在增殖的LMMCF-7细胞中上调NDRG2表达,细胞增殖指数(proliferation index,PI)由47.18%下降至31.78%(P<0.001);在低增殖的MCF-7细胞中沉默NDRG2表达,PI由32.00%上升至52.59%(P<0.001);Western blot及免疫荧光结果显示,NDRG2可抑制β-catenin表达,并抑制其聚集入核;流式细胞检测结果显示,共转染siRNA和pCMV-Tcfδ的MCF-7细胞增殖能力未增强,进一步说明NDRG2是通过抑制β-catenin的转录调节作用抑制肿瘤细胞增殖。结论:在乳腺癌细胞中,NDRG2的表达水平下调,导致β-catenin聚集入核,并激活下游靶基因,促进乳腺癌细胞增殖。此分子机制对阐明乳腺癌细胞增殖调控机制具有重要意义。
乳腺肿瘤;N-myc下游调节基因2;β-连环蛋白;细胞增殖
乳腺癌是女性常见的恶性肿瘤,全球每年约有138万女性被诊断为乳腺癌,占全部新发肿瘤的23%。我国每年女性乳腺癌发病16.9万例,居女性常见肿瘤首位。有研究表明,肿瘤细胞的恶性增殖和分裂失调是肿瘤发生、发展的基础[1]。癌细胞获得高增殖能力是乳腺癌后期转移的重要条件之一,研究癌细胞增殖调控机制对乳腺癌的诊断和治疗具有重要价值。
N-myc下游调节基因2(N-myc downstream regulated gene 2,NDRG2)是NDRG家族的重要成员,其基因染色体定位于14q11.2,含有16个外显子和15个内含子,表达蛋白含357个氨基酸残基,相对分子质量约41×103。NDRG2具有APC样结构域,受原癌基因N-Myc的表达调控。近年研究发现,NDGR2基因除参与胚胎组织发育、细胞分化及血液免疫外,还与肿瘤进展具有密切关系[2]。NDRG2在肿瘤的发生、发展中起到抑癌基因的作用[3]。在肿瘤细胞中,NDRG2启动子常发生甲基化[4-5],因此在多种肿瘤中,如消化系统肿瘤、甲状腺癌、肺癌、和中枢神经肿瘤中呈显著低表达,且与肿瘤的淋巴转移呈负相关[4,6-10]。但NDRG2是否可抑制乳腺癌细胞增殖及其分子机制目前尚不清楚。
β-连环蛋白(β-catenin)作为Wnt信号通路中的关键因子,通过T细胞特异性转录因子(T cellspecific transcription factor,Tcf)或其他转录因子与特定的DNA序列相结合,激活靶基因的转录[11],在乳腺癌的发生、发展中发挥重要调控作用。肿瘤细胞可通过上调Dsh蛋白或下调Frp1蛋白表达激活经典的Wnt途径,并上调β-catenin表达[12-13]。肿瘤细胞也可通过非Wnt通路机制稳定β-catenin表达,如通过调控PTEN、ILK、IKKA、Pin1及P53等因子的表达进而调节β-catenin[14]。前期研究发现,ERK1/2信号通路的激活可上调β-catenin表达,并促进其转移入核,发挥转录调节作用,促进乳腺癌细胞增殖和迁移[15-16],但β-catenin是否受到NDRG2的调控需要进一步研究。
在前期研究中,我们应用严重联合免疫缺陷小鼠从人乳腺癌细胞系MCF-7中筛选和建立了具有高转移能力的转移亚克隆LM-MCF-7。与MCF-7细胞相比,LM-MCF-7细胞具有更强的增殖和迁移能力,动物实验显示,其具有成瘤早、转移快和转移部位广泛等特点。基因芯片研究结果显示,在人的21 329种基因中,有67种基因的表达水平在LM-MCF-7细胞中和MCF-7细胞中差异有统计学意义[15]。本研究应用MCF-7和LM-MCF-7细胞模型研究NDRG2对乳腺癌细胞增殖的调控作用。
1 材料和方法
1.1 细胞株及主要试剂
人乳腺癌细胞系MCF-7、LM-MCF-7、pCMV-tag2b质粒和pCMV-Tcfδ质粒(表达的Tcfδ具有蛋白结合位点但缺少DNA结合位点,可竞争性抑制Tcf的转录调控功能)为本实验室储备;RPMI1640培养基、胎牛血清和LipofectamineTM2000购自美国Invitrogen公司;兔抗人NDRG2多克隆抗体、鼠抗人β-actin单克隆抗体和碘化丙啶购自美国Sigma-Aldrich公司;兔抗人β-catenin多克隆抗体购自美国Santacruz公司;小量总RNA提取试剂盒购自德国Qiagen公司;QuantScript RT试剂盒购自天根公司。靶向NDRG2的小RNA干扰片段由广州市锐博生物科技有限公司合成,NDRG2 siRNA正义序列:5’-ACAUCCUGGCGAGAUAUGCUCUUAA-3’,反义序列:5’-UUAAGAGCAUAUCUCGCCAGGA UGU-3’;对照siRNA正义序列:5’-UUCUCCGA A C G U G U C A C G U T T-3’,反义序列:5’-ACGUGACACGUUCGGAGAATT-3’。其余试剂为国产分析纯。
1.2 细胞培养
MCF-7和LM-MCF-7细胞采用含10%胎牛血清、100 U/mL青霉素和100 U/mL硫酸链霉素的RPMI1640培养基,于37 ℃、CO2体积分数为5%的条件下培养。
1.3 pCMV-NDRG2真核表达载体构建
用小量总RNA提取试剂盒提取MCF-7细胞总RNA,利用QuantScript RT试剂盒制备cDNA。根据NDRG2(GENEBANK:NM_201535.1)mRNA CDS序列设计引物:上游加入EcoRⅠ酶切位点,下游加入SalⅠ酶切位点。上、下游引物序列分别为5’-GAATTCATGGCGGAGCTGCAGGA GGTG-3’和5’-GTCGACTCAACAGGAGACCTCC ATGGT-3’。以反转录的cDNA为模板。用上述引物PCR扩增,获得目的基因片段(PCR反应参数:94 ℃变性5 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸90 s,共20个循环,72 ℃延伸7 min)。
扩增产物经EcoRⅠ/SalⅠ双酶切纯化后,与pCMV重组,转化DH5α菌。提取转化菌质粒,经双酶切和DNA测序分析,对NDRG2序列进行鉴定,得到重组真核表达载体pCMVNDRG2。
1.4 细胞转染
将培养至覆盖率达80%的MCF-7和LMMCF-7细胞用于转染实验。将8 μg质粒(或80 nmol/L siRNA)和24 μL LipofectamineTM2000分别加入到500 μL的OMEM培养基中,室温静置5 min,然后将二者混匀并于37 ℃静置15 min。随后将质粒(或siRNA)/LipofectamineTM2000/OMEM混合物加入细胞中,37 ℃温育6 h后换成无抗生素的完全培养基,继续培养72 h后进行蛋白[质]印迹法(Western blot)或流式细胞实验。
1.5 流式细胞实验检测乳腺癌细胞增殖
用胰蛋白酶消化收集正常培养及转染后的乳腺癌细胞,PBS洗2遍,1 000×g离心5 min,70%的冰乙醇固定,4 ℃过夜。上机检测前细胞用PBS洗2遍后重悬在含50 mg/L RNA酶和25 mg/L碘化丙锭的PBS中,室温避光温育15 min后用FACS Calibur流式细胞仪进行单色荧光细胞流式检测。每个样本采集1×104个细胞,用ModFit软件(购自美国Verity Software House公司)分析结果。每组实验重复3次。
1.6 免疫荧光检测β-catenin细胞定位
将正常培养及转染后的细胞接种在放有盖玻片的六孔板中,37 ℃过夜培养。随后用-20 ℃预冷的无水甲醇于4 ℃固定15 min,用含5%Triton X-100的PBS洗细胞,5 min×2次。加入10%血清,室温封闭20 min。加入兔抗人β-catenin抗体,室温温育2 h。PBS洗细胞,5 min×3次。滴加FITC标记的羊抗兔荧光二抗,室温温育1 h。PBS洗细胞,5 min×3次。用DAPI对细胞核进行染色,室温温育1 h,PBS洗细胞,5 min×3次。封片后,置于荧光显微镜下观察结果。
1.7 Western blot
收获正常培养和转染后的细胞,用预冷的PBS洗细胞2次,随后加入细胞裂解液(10 mmol/L Tris-HCl,pH=8.0,0.1 mmol/L EDTA,150mmol/L NaCl,1%NP-40,1 mmol/L PMSF,1%SDS)冰上放置20 min,4 ℃,12 000×g离心20 min,收集上清定量分析,取30 μg总蛋白进行12%SDS-PAGE电泳,随后电转印至PVDF膜上,5%脱脂奶粉封闭2 h,加入相应一抗,室温温育2 h,TBS-T洗膜后加入相应二抗室温温育1 h,TBS-T洗膜后,应用增强型ECL显色试剂盒于暗室曝光显影,实验均重复3次。
1.8 统计学处理
2 结 果
2.1 过表达NDRG2抑制乳腺癌细胞增殖
Western blot结果显示,与高增殖能力的LM-MCF-7细胞相比,NDRG2在低增殖能力的MCF-7中表达较高(如图1A)。NDRG2表达同肿瘤细胞增殖能力负相关。转染MCF-7细胞NDRG2 siRNA,沉默NDRG2表达,流式细胞实验检测结果显示,与转染对照siRNA组相比,细胞增殖指数(proliferation index,PI)由32.00%上升至52.59%(P<0.01)。利用pCMV-tag2b真核表达载体,构建pCMV-NDRG2载体(图1B)。转染LM-MCF-7细胞pCMV-NDRG2质粒,过表达NDRG2(图1C),流式细胞实验检测结果显示,与转染对照pCMV-tag2b组相比,PI由47.18%下降至31.78%(P<0.01,图1D)。以上结果说明,NDRG2表达水平同细胞增殖能力负相关,NDRG2表达下调是乳腺癌细胞获得高增殖能力的重要原因。
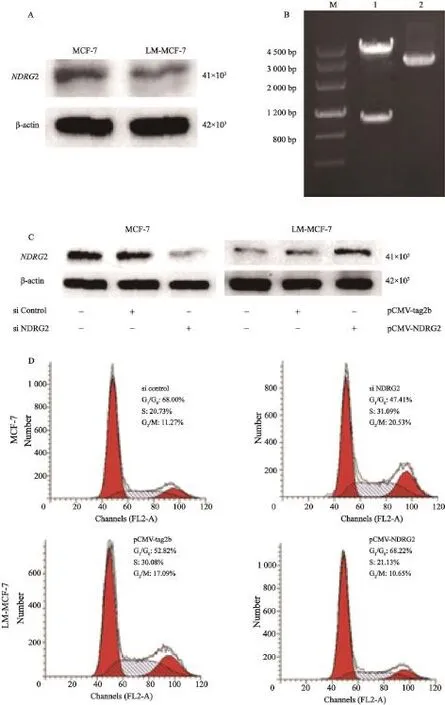
图 1 过表达NDRG2抑制乳腺癌细胞增殖Fig. 1 The proliferation of breast cancer cells was inhibited by over-expressing NDRG2
2.2 NDRG2抑制β-catenin表达并促进其磷酸化
本研究通过Western blot检测改变NDRG2表达水平后,β-catenin的表达水平及磷酸化水平变化。转染MCF-7细胞siRNA,沉默NDRG2表达,Western blot结果显示,同转染对照siRNA组相比,β-catenin表达水平显著上调,且磷酸化的p-β-catenin水平下降(图2A)。转染LM-MCF-7细胞pCMV-NDRG2表达载体,上调NDRG2表达水平。Western blot结果显示,β-catenin表达水平下降,并且磷酸化的p-β-catenin表达水平上调(图2B)。以上结果说明,NDRG2能够抑制β-catenin表达,并且促进β-catenin磷酸化,促使其进入泛素化降解途径。
2.3 NDRG2抑制β-catenin进入细胞核
β-catenin发挥促增殖作用主要依赖其在细胞质中聚集并进入细胞核,与Tcf/LEF结合形成Tcf/LEF/β-catenin复合体,特异性启动和激活下游靶基因的转录[17]。利用β-catenin特异性抗体,通过免疫荧光技术对MCF-7和LM-MCF-7中β-catenin在细胞中的定位进行分析。结果发现,β-catenin在高增殖能力的LM-MCF-7细胞中存在明显的核定位,而在低增殖能力的MCF-7细胞中主要存在于细胞质。利用RNA干扰方法沉默MCF-7细胞中的NDRG2表达后,免疫荧光结果发现,β-catenin由细胞质转移进入细胞核(图3A)。在LM-MCF-7细胞中过表达NDRG2后,β-catenin主要定位于细胞质,而核定位的β-catenin接近消失(图3A)。免疫荧光结果说明,NDRG2能够抑制β-catenin聚集入核。为进一步验证NDRG2抑制β-catenin的转录调节作用,进而抑制乳腺癌细胞增殖,我们共转染MCF-7细胞NDRG2 siRNA和pCMV-Tcfδ,在降低NDRG2表达的同时,过表达Tcfδ(具有蛋白结合位点,但缺失DNA结合位点的Tcf突变体,能够同野生型Tcf竞争性结合β-catenin,抑制下游基因的转录激活[15])。流式细胞检测结果显示,干扰NDRG2表达后,与对照组相比,MCF-7细胞PI明显上升至53.25%(P<0.01);干扰NDRG2的同时过表达Tcfδ,与pCMV-tag2b相比,细胞PI下降至40.78%(P<0.05%,图3B)。流式细胞结果表明,在乳腺癌细胞中,NDRG2表达下降,促进了β-catenin入核,并与Tcf结合促进下游因子转录。此分子机制是乳腺癌细胞获得高增殖能力的重要原因。
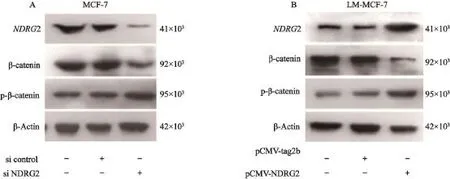
图 2 NDRG2调控β-catenin表达及其磷酸化Fig. 2 NDRG2 regulated the expression and phosphorylation of β-catenin
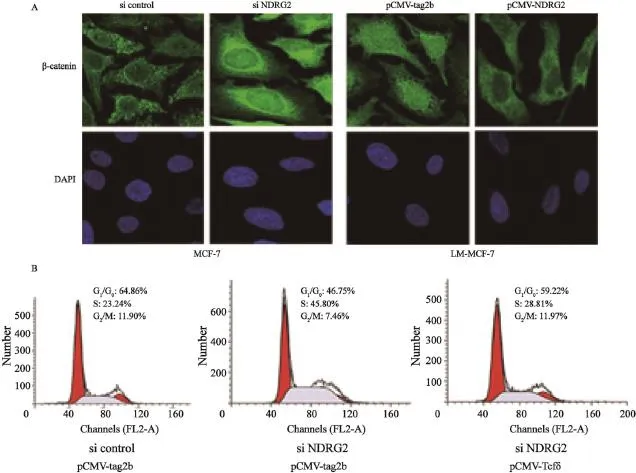
图 3 NDRG2抑制β-catenin进入细胞核Fig. 3 Translocation of β-catenin into the nucleus was inhibited by NDRG2
3 讨 论
细胞增殖和分裂失调是乳腺肿瘤发生、发展的基础,深入研究癌细胞的增殖调控机制对肿瘤的诊断和治疗具有重要意义。目前研究已证实,NDRG2除参与正常细胞的生长、分化以及应激反应外,还与肿瘤的发生、发展和转归密切相关。肿瘤组织的NDRG2表达显著低于癌旁及正常组织,表达水平随肿瘤TNM分期增加或恶性度增加而降低[2]。本研究利用具有相同遗传背景但具有不同增殖能力的乳腺癌细胞系MCF-7和LM-MCF-7[15]探明NDRG2在调控乳腺癌细胞增殖中的作用,发现NDRG2的表达水平与乳腺癌细胞的增殖能力呈负相关,提高NDRG2的表达水平可显著抑制乳腺癌细胞增殖;相反,RNAi沉默NDRG2表达后,细胞增殖能力显著增强。目前已有多项研究证实,NDRG2在乳腺肿瘤中起到抑癌基因的作用,通过多种分子机制抑制乳腺肿瘤生长、细胞增殖及转移。在乳腺癌细胞SKBR-3和MCF-7中过表达NDRG2能够抑制细胞增殖,下调促血管生成因子VEGF和HIF-1α的表达并抑制血管新生,进而抑制裸鼠移植瘤生长[18-19]。在MDA-MB-231细胞中过表达NDRG2,能够激活BMP-4通路,并通过抑制MMP-9活性抑制细胞侵袭[20]。Kim等[21]研究发现,在乳腺癌细胞中过表达NDRG2能够抑制NF-κB信号通路的激活,进而下调COX-2表达,抑制肿瘤细胞迁移和侵袭。NDRG2还可通过激活p38 MAPK途径,抑制细胞因子信号转导抑制因子1磷酸化,进而抑制细胞增殖和迁移[22]。在乳腺癌细胞中过表达NDRG2可促进SAPK和JNK磷酸化,促进肿瘤细胞凋亡[22]。除乳腺肿瘤外,NDRG2对多种肿瘤生长、肿瘤细胞的增殖、迁移和侵袭具有抑制作用。在大肠癌SW620细胞中,上调NDRG2表达可降低c-Jun磷酸化水平,减弱转录调节因子AP-1活性,进而抑制cyclinD1表达,抑制细胞增殖[23]。在肝癌细胞中,NDRG2还可通过上调p38磷酸化水平抑制细胞增殖[24];通过下调CD24的表达抑制细胞黏附、迁移和侵袭[25]。NDRG2还可通过促进葡萄糖转运蛋白GLUT1降解阻断肿瘤细胞能量代谢,抑制肿瘤生长[26]。过表达NDRG2可增强GSK-3β活性,促进Snail降解,进而上调E-cadherin抑制肿瘤细胞侵袭[27]。NDRG2还可通过抑制MMP2和laminin332信号通路并同时降低Rho GTPase活性,抑制由TGFβ1介导的肿瘤细胞侵袭[28]。
β-catenin蛋白是介导Wnt信号传导的关键分子,其在细胞内的表达水平决定着Wnt信号通路的开放或关闭,在乳腺癌发生、发展过程中起到关键作用[29]。β-catenin在细胞内的水平受到严格调控,由GSK-3β、Axin及APC组成的复合体能够使β-catenin磷酸化,并进入泛素化降解途径。当磷酸化被抑制时,β-catenin在细胞质中聚集并进入细胞核,形成Tcf/LEF/β-catenin复合体,特异性启动和激活下游靶基因[30]。本研究发现,在乳腺癌细胞中上调NDRG2水平可促进β-catenin磷酸化,下调细胞内β-catenin水平;免疫荧光发现,NDRG2可抑制细胞质的β-catenin转移入核,阻碍其发挥转录激活作用。近年来多项相关研究均证实,乳腺癌的发生与β-catenin表达异常有关。β-catenin的表达激活可增强cyclinD1表达,进而促进肿瘤细胞异常增殖[29];C-myc基因启动子上有Tcf结合位点,β-catenin/TCF复合物可促进其基因表达,从而导致肿瘤细胞增殖[14]。乳腺癌细胞中有多种途径可稳定调节β-catenin,如Pin1蛋白在80%的乳腺癌中上调,Pin1能够抑制APC蛋白作用,从而稳定β-catenin处于高表达水平[31]。PTEN基因的丢失及异常激活Akt蛋白能够激活β-catenin,引起乳腺癌细胞增殖[32]。肿瘤细胞中p53蛋白能够通过GSK-3β依赖性和Siah依赖性机制稳定β-catenin表达,促进肿瘤细胞增殖[33]。我们的前期研究还发现,在乳腺癌细胞中,激活的MLCK-ERK1/2正反馈信号途径通过促进β-catenin高表达及转移入核,激活cyclin D1和Survivin表达,维持乳腺癌细胞高增殖和高迁移能力[15]。在肿瘤细胞中,β-catenin作为信号因子受到多种信号途径调控,对肿瘤细胞增殖发挥着关键调控作用。
综上所述,NDRG2作为肿瘤抑制因子,在调控乳腺癌细胞增殖中发挥重要的抑制作用。NDRG2抑制β-catenin表达,阻碍其转移入核发挥抑制转录调节作用是乳腺癌细胞获得高增殖能力的重要原因。此分子调控机制的阐明为乳腺癌的诊断和治疗提供了重要理论依据。
[1]CHEN X, DUAN N, ZHANG C, et al. Survivin and tumorigenesis: molecular mechanisms and therapeutic strategies[J]. J Cancer, 2016, 7(3): 314-323.
[2]HU W, FAN C, JIANG P, et al. Emerging role of N-myc downstream-regulated gene 2 (NDRG2) in cancer[J]. Oncotarget, 2016, 7(1): 209-223.
[3]GOLESTAN A M, MOJTAHEDI Z P, GHALAMFARSA G P, et al. The effects of NDRG2 overexpression on cell proliferation and invasiveness of SW48 colorectal cancer cell line[J]. Iran J Med Sci, 2015, 40(5): 430-439.
[4]PIEPOLI A, COTUGNO R, MERLA G, et al. Promoter methylation correlates with reduced NDRG2 expression in advanced colon tumour[J]. BMC Med Genomics, 2009, 2: 11.
[5]FENG L, XIE Y, ZHANG H, et al. Down-regulation of NDRG2 gene expression in human colorectal cancer involves promoter methylation and microRNA-650[J]. Biochem Biophys Res Commun, 2011, 406(4): 534-538.
[6]LORENTZEN A, LEWINSKY R H, BORNHOLDT J, et al. Expression profile of the N-myc downstream regulated gene 2 (NDRG2) in human cancers with focus on breast cancer[J]. BMC Cancer, 2011, 11: 14.
[7]MORDALSKA A, LATEK J, FERENC T, et al. Evaluation of NDRG2 gene expression in primary papillary thyroid carcinoma and in metastases of this neoplasm to regional lymph nodes[J]. Thyroid Res, 2010, 3(1): 6.
[8]PARK M Y, CHOI S C, LEE H S, et al. A quantitative analysis of N-myc downstream regulated gene 2 (NDRG2) in humantissues and cell lysates by reverse-phase protein microarray[J]. Clin Chim Acta, 2008, 387(1-2): 84-89.
[9]SHI H, LI N, LI S, et al. Expression of NDRG2 in esophageal squamous cell carcinoma[J]. Cancer Sci, 2010, 101(5): 1292-1299.
[10]SKIRIUTE D, TAMASAUSKAS S, ASMONIENE V, et al. Tumor grade-related NDRG2 gene expression in primary and recurrent intracranial meningiomas[J]. J Neurooncol, 2011, 102(1): 89-94.
[11]DING Y, SU S, TANG W, et al. Enrichment of the betacatenin-TCF complex at the S and G2phases ensures cell survival and cell cycle progression[J]. J Cell Sci, 2014, 127(Pt 22): 4833-4845.
[12]NAGAHATA T, SHIMADA T, HARADA A, et al. Amplification, up-regulation and over-expression of DVL-1, the human counterpart of the drosophila disheveled gene, in primary breast cancers[J]. Cancer Sci, 2003, 94(6): 515-518.
[13]UGOLINI F, CHARAFE-JAUFFRET E, BARDOU V J, et al. WNT pathway and mammary carcinogenesis: loss of expression of candidate tumor suppressor gene SFRP1 in most invasive carcinomas except of the medullary type[J]. Oncogene, 2001, 20(41): 5810-5817.
[14]HATSELL S, ROWLANDS T, HIREMATH M, et al. Betacatenin and Tcfs in mammary development and cancer[J]. J Mammary Gland Biol Neoplasia, 2003, 8(2): 145-158.
[15]ZHOU X, LIU Y, YOU J, et al. Myosin light-chain kinase contributes to the proliferation and migration of breast cancer cells through cross-talk with activated ERK1/2[J]. Cancer Lett, 2008, 270(2): 312-327.
[16]ZHOU X L, QIN X R, ZHANG X D, et al. Downregulation of Dickkopf-1 is responsible for high proliferation of breast cancer cells via losing control of Wnt/beta-catenin signaling[J]. Acta Pharmacol Sin, 2010, 31(2): 202-210.
[17]CUELLO-CARRION F D, SHORTREDE J E, ALVAREZOLMEDO D, et al. HER2 and beta-catenin protein location: importance in the prognosis of breast cancer patients and their correlation when breast cancer cells suffer stressful situations[J]. Clin Exp Metastasis, 2015, 32(2): 151-168.
[18]朱财林, 李南林, 王 廷, 等. NDRG2对乳腺癌细胞增殖的影响[J]. 医学争鸣, 2009, 30(11): 961-963.
[19]MA J, LIU W, YAN X, et al. Inhibition of endothelial cell proliferation and tumor angiogenesis by up-regulating NDRG2 expression in breast cancer cells[J]. PLoS One, 2012, 7(2): e32368.
[20]SHON S K, KIM A, KIM J Y, et al. Bone morphogenetic protein-4 induced by NDRG2 expression inhibits MMP-9 activity in breast cancer cells[J]. Biochem Biophys Res Commun, 2009, 385(2): 198-203.
[21]KIM M J, KIM H S, LEE S H, et al. NDRG2 controls COX-2/ PGE(2)-mediated breast cancer cell migration and invasion[J]. Mol Cells, 2014, 37(10): 759-765.
[22]PARK Y, SHON S K, KIM A, et al. SOCS1 induced by NDRG2 expression negatively regulates STAT3 activation in breast cancer cells[J]. Biochem Biophys Res Commun, 2007, 363(2): 361-367.
[23]KIM Y J, YOON S Y, KIM J T, et al. NDRG2 suppresses cell proliferation through down-regulation of AP-1 activity in human colon carcinoma cells[J]. Int J Cancer, 2009, 124(1): 7-15.
[24]LIU X, NIU T, LIU X, et al. Microarray profiling of HepG2 cells ectopically expressing NDRG2[J]. Gene, 2012, 503(1): 48-55.
[25]ZHENG J, LI Y, YANG J, et al. NDRG2 inhibits hepatocellular carcinoma adhesion, migration and invasion by regulating CD24 expression[J]. BMC Cancer, 2011, 11: 251-259.
[26]MA J, LIU W, GUO H, et al. N-myc downstream-regulated gene 2 expression is associated with glucose transport and correlated with prognosis in breast carcinoma[J]. Breast Cancer Res, 2014, 16(2): R27.
[27]KIM Y J, KANG H B, YIM H S, et al. NDRG2 positively regulates E-cadherin expression and prolongs overall survival in colon cancer patients[J]. Oncol Rep, 2013, 30(4): 1890-1898.
[28]LEE D C, KANG Y K, KIM W H, et al. Functional and clinical evidence for NDRG2 as a candidate suppressor of liver cancer metastasis[J]. Cancer Res, 2008, 68(11): 4210-4220.
[29]JAMIESON C, SHARMA M, HENDERSON B R. Targeting the beta-catenin nuclear transport pathway in cancer[J]. Semin Cancer Biol, 2014, 27: 20-29.
[30]MORGAN R G, RIDSDALE J, TONKS A, et al. Factors affecting the nuclear localization of beta-catenin in normal and malignant tissue[J]. J Cell Biochem, 2014, 115(8): 1351-1361.
[31]RYO A, NAKAMURA M, WULF G, et al. Pin1 regulates turnover and subcellular localization of beta-catenin by inhibiting its interaction with APC[J]. Nat Cell Biol, 2001, 3(9): 793-801.
[32]DEPOWSKI P L, ROSENTHAL S I, ROSS J S. Loss of expression of the PTEN gene protein product is associated with poor outcome in breast cancer[J]. Mod Pathol, 2001, 14(7): 672-676.
[33]WANG D, WANG Y, KONG T, et al. Hypoxia-induced betacatenin downregulation involves p53-dependent activation of Siah-1[J]. Cancer Sci, 2011, 102(7): 1322-1328.
NDRG2 inhibits the proliferation of breast cancer cells via regulating β-catenin expression and nuclear translocation
ZHOU Xiaolei, ZHU Chongyue, ZHANG Shiguang, ZHOU Zhiyan, LI Haichao, ZOU Wei (Public R&D Center of Bio-Manufacture, Hebei University of Science & Technology, Shijiazhuang 050018, Hebei Province, China)
Background and purpose:Breast cancer is one of the most common malignant diseases in women and its malignant proliferation is the major cause of death. To investigate the effects of N-myc downstream regulated gene 2 (NDRG2) on proliferation of breast cancer cells by using two parallel cell lines (MCF-7 and LM-MCF-7) with different metastatic abilities.Methods:The expression level of NDRG2 in breast cancer cells was detected by Western blot. The effects of overexpressing (or down-regulating) NDRG2 on proliferation of breast cancer cells were investigated by flow cytometry. The expression and location of β-catenin were detected by Western blot and immunofluorescence respectively. NDRG2 blocking the transcription activity of β-catenin was investigated via co-transfecting MCF-7 cells with NDRG2 siRNA and pCMV-Tcfδ (lacking the portion responsible for the protein binding to DNA).Results:The expression level of NDRG2 was negatively related to the proliferation ability of breast cancer cells. Over-expressing NDRG2 (or down-regulating) via transfecting LM-MCF-7 (or MCF-7) cells with pCMV-NDRG2 (or NDRG2 siRNA) could inhibit (or promote) cell proliferation. Interestingly, the results of Western blot, immunofluorescence and flow cytometry revealed that down-regulation of NDRG2 resulted from the down-regulation of β-catenin and blocking its nuclear translocation, which led to losing control of the proliferation of breast cancer cells.Conclusion:NDRG2 inhibitthe proliferation of breast cancer cells via down-regulating the expression of β-catenin and blocking its nuclear translocation, which is significant for exploring the molecular mechanism of proliferation of breast cancer cells.
Breast neoplasms; N-myc downstream regulated gene 2; β-catenin; Cell proliferation
10.19401/j.cnki.1007-3639.2016.12.004
R737.9
A
1007-3639(2016)12-0981-08
2016-04-25
2016-06-27)
河北省高等学校科学研究项目(QN2016020);2015年度河北科技大学五大平台开放基金课题;河北科技大学博士科研启动基金课题。
周晓雷 E-mail: foxlei@live.cn
猜你喜欢
中老年保健(2022年6期)2022-08-19 01:41:48
学苑创造·A版(2020年12期)2020-01-07 14:07:23
中国外汇(2019年15期)2019-10-14 01:00:34
中国生殖健康(2019年2期)2019-08-23 08:11:42
天津医科大学学报(2019年6期)2019-08-13 07:04:42
中国生殖健康(2019年6期)2019-01-06 09:20:12
祝您健康(2018年5期)2018-05-16 17:10:16
作文教学研究(2016年1期)2016-07-05 12:22:47
安徽医科大学学报(2015年9期)2015-12-16 11:09:42
医学研究杂志(2015年8期)2015-06-22 14:00:57
