
广玉兰在自然降温条件下的光合响应机制研究
2016-02-06 07:25:59陈丽培王国霞杨玉珍刘艳萍刘瑞霞
河南农业科学 2016年1期
陈丽培,王国霞,杨玉珍*,刘艳萍,刘瑞霞
(1.郑州师范学院 生命科学学院,河南 郑州 450044; 2.河南省林业科学研究院,河南 郑州450008)
广玉兰在自然降温条件下的光合响应机制研究
陈丽培1,王国霞1,杨玉珍1*,刘艳萍2,刘瑞霞1
(1.郑州师范学院 生命科学学院,河南 郑州 450044; 2.河南省林业科学研究院,河南 郑州450008)
以郑州市15年生广玉兰的叶片为材料,运用 Li-6400光合测定仪测定秋冬季广玉兰叶片净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)等,研究其在秋冬自然降温过程中的光合响应机制。结果表明:在秋冬季节,随着平均气温的下降,广玉兰叶片的Pn、Gs、Tr均呈下降趋势,Ci则呈上升趋势。在温度下降过程中,广玉兰叶片仍具有一定的光合能力,并表现出明显的日变化规律,即Pn、Gs、Tr的日变化均呈先升高后下降的单峰曲线,Ci日变化呈“V”字型曲线,Pn不存在“午休”现象。
广玉兰; 自然降温; 光合速率; 气孔导度; 响应机制
低温是影响植物生长发育的主要环境因子[1],也是低纬度树木向高纬度区域引种的主要限制因子。广玉兰(Magnoliagrandiflora)别称洋玉兰、荷花玉兰,属木兰科木兰属常绿乔木[2]。广玉兰枝繁叶大、四季常青、花期较长且洁白芳香,不仅具有很高的观赏价值,而且具有较强抗烟尘、抗污染等特性,是城市园林绿化和环境美化的主要树种[3]。广玉兰广泛栽培于长江以南和长江流域,作为向高纬度地区引种和应用的重要树种,其面临着低温胁迫问题。目前,我国对广玉兰低温胁迫的生理响应研究较多,刘艳萍等[4]通过研究低温处理后不同类型广玉兰叶片低温半致死温度以及相对电导率和叶片组织细胞结构的变化,揭示了6种不同类型广玉兰的抗寒性;周建等[5]通过测定低温处理后广玉兰叶片中脯氨酸含量、相对电导率、丙二醛含量、可溶性糖含量及超氧化物歧化酶、过氧化物酶活性的变化,探讨其影响机制;张亚红等[6]研究自然降温过程中,广玉兰叶片细胞质膜相对透性、丙二醛含量、脯氨酸含量、超氧化物歧化酶活性的动态变化过程,揭示广玉兰的抗寒性。上述研究主要针对低温胁迫下广玉兰生理生化特性的研究,而对于光合生理生态因子的研究则相对较少。光合作用是地球上植物重要的代谢反应,也是植物受低温胁迫最敏感的生理过程之一[7]。因此,研究植物光合作用对低温胁迫的响应对于减轻植物低温伤害、向高纬度城市引种和应用具有重要的理论和实践意义。鉴于此,本研究以郑州市15年生广玉兰叶片为供试材料,通过探索自然降温过程中叶片净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)的动态变化,揭示广玉兰对自然低温胁迫的光合响应机制,以及为广玉兰安全越冬和向北方城市进行园林引种和栽培提供依据。
1 材料和方法
1.1 研究地区概况
郑州市地处34°16′~34°58′N,112°42′~114°13′E,属北温带大陆性季风气候,冷暖适中、四季分明,春季干旱少雨,夏季炎热多雨,秋季晴朗日照长,冬季寒冷少雪。测定期间郑州市气温状况见表1。
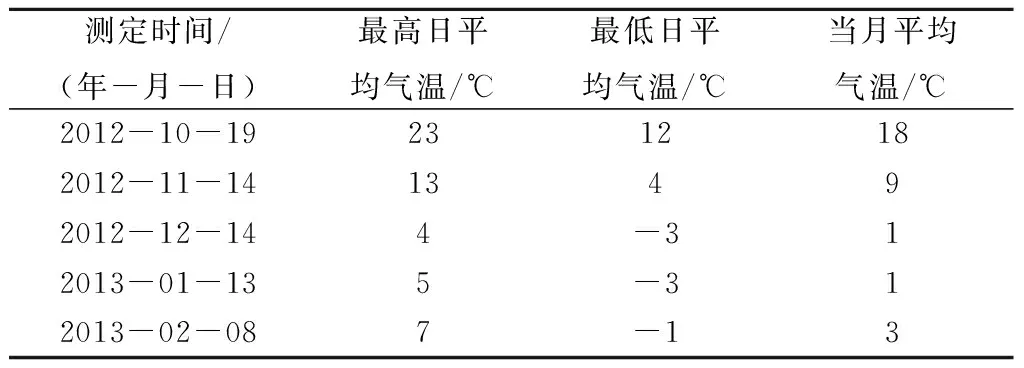
表1 测定期间郑州市气温
1.2 试验材料
试验材料为郑州市某小区的广玉兰叶片,树龄约15 a,树高平均10.5 m,胸径16.0 cm。
1.3 试验设计
于2012年10月至2013年2月,每月的上、中、下旬分别于晴天的10:00—11:00测定1次光合指标。因广玉兰植株高大,且经测定其离体叶片在1 h内均有较高、较稳定的水分供应和光合活力,所以叶片光合指标采用离体测定。从广玉兰树冠中部东、西、南、北4个方向各剪取1个枝条插入水中,环剥去除切口处3 cm左右的韧皮部,并分别从各枝条中选取1片健康、无病虫害的叶片,共4片,每片叶片测定3次,取平均值。另外,每月中旬选择明显降温的晴天9:00—17:00进行光合指标的日变化测定,每2 h测定1次。每次测定选取4片叶,每片叶片测定3次,取平均值。
1.4 测定指标及方法
利用美国li-Cor公司生产的Li-6400便携式光合测定仪测定广玉兰叶片Pn、Gs、Tr、Ci等。测定过程采用开放式气路,环境CO2浓度为400 μmol/mol,空气流速设定为500 μmol/s。
2 结果与分析
2.1 自然降温过程中广玉兰叶片光合因子的变化
2.1.1 Pn和Gs Pn是反映植物光合能力强弱的主要指标,也是反映低温胁迫下植物光响应特征的重要指标[8]。由图1可知,广玉兰叶片Pn在自然降温过程中呈下降趋势。10月广玉兰叶片Pn最高,为4.48 μmol/(m2·s),之后逐渐降低。10月至12月期间Pn降幅较大,12月[0.87 μmol/(m2·s)]比10月降低了80.58%。12月至翌年1月期间Pn变化较平缓,1月仅比12月降低9.19%,基本保持在0.79~0.87 μmol/(m2·s)。2013年1月Pn为测定期内最低值,比2012年10月降低了82.37%。可见,广玉兰光合作用受低温影响较大,叶片Pn明显降低。2013年2月Pn随气温的小幅度回升有所升高。总的来说,广玉兰叶片Pn在自然降温过程中的变化规律反映了其耐寒性和对秋冬季节的适应性。
气孔既是CO2进入叶肉细胞的入口,也是蒸腾作用水分逸出叶片的出口[9],气孔的打开程度影响叶片光合作用的碳固定。自然降温过程中广玉兰叶片Gs的变化如图1所示。Gs和Pn的变化趋势相似,表现出随温度的降低而降低的趋势。其中,10月份Gs最高,为0.074 2 μmol/(m2·s)。之后随温度的降低,Gs迅速降低,到12月降为最低值[0.019 3 μmol/(m2·s)],比10月份降低了73.99%。12月至翌年2月Gs又有所回升。Gs的变化曲线与Pn具有一致性,说明在自然降温过程中,Pn的胁迫效应可能与Gs受抑制有关。
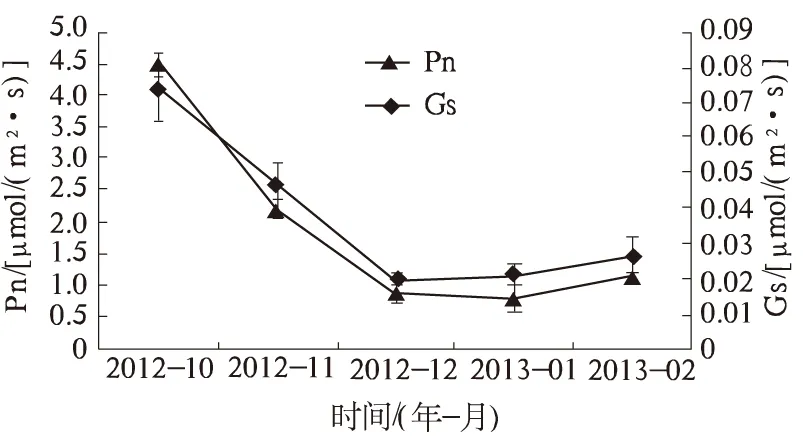
图1 自然降温对广玉兰叶片Pn、Gs的影响
2.1.2 Tr 蒸腾作用是一个复杂的生理过程,不但能够降低叶片的温度,加强水分和无机离子到叶片的运输,同时会引起气孔关闭,制约CO2向叶片的扩散进程,与光合作用存在着相互依赖的关系。Tr是反映蒸腾作用强弱的主要指标,Tr的变化是由于外界环境因子的变化而产生的自适应调整[10]。广玉兰叶片Tr在温度降低过程中呈下降趋势(图2)。从10月开始,Tr随温度的降低而降低,在12月降为最低值[0.25 mmol/(m2·s)]。其中10—12月Tr下降幅度大,相对于10月,11月、12月分别降低了36.25%、67.79%。这与从秋季到冬季过度期间光照强度、温度波动大有关。Tr在12月至翌年2月间呈波动性变化,基本稳定在0.25~0.30 mmol/(m2·s)。
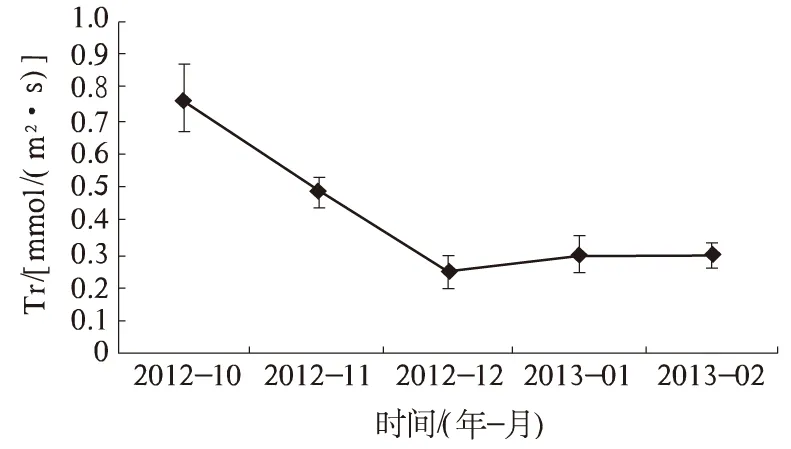
图2 自然降温对广玉兰Tr的影响
2.1.3 Ci CO2是植物进行光合作用的必要原料之一[11]。它通过气孔进入叶肉细胞参与植物的光合生理过程。图3表明,广玉兰叶片的Ci随温度降低呈不同程度的升高趋势。这与Pn、Gs、Tr随温度降低而降低的趋势相反。说明低温胁迫下,广玉兰气孔抑制不是光合作用的主要原因。从10月开始,Ci迅速上升,并于12月达到最高值(294.702 μmol/mol),比10月升高了83.94%。随后Ci在1月、2月稍微有所下降,2月Ci为286.92 μmol/mol,仍比10月升高了79.09%。总的来说,广玉兰叶片Ci在10—12月升高幅度大,12月至翌年2月稍有下降,整体表现为升高趋势。
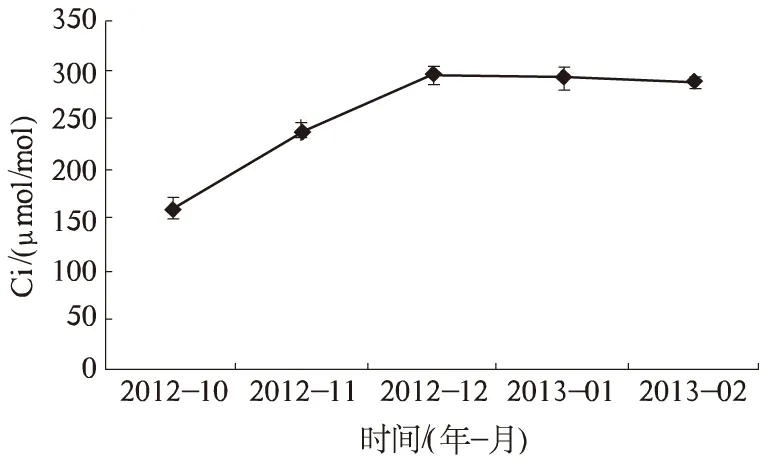
图3 自然降温对广玉兰Ci的影响
2.2 自然降温过程中广玉兰叶片光合因子的日变化
2.2.1 Pn 由图4可知,Pn的日变化为单峰曲线,没有明显的光合“午休”现象。大致是9:00—11:00 Pn迅速上升,11:00达到最大值。10月、11月、12月、翌年1月、2月Pn最大值分别为4.70、2.24、0.90、0.76、1.15 μmol/(m2·s)。说明此时的温度和光强可能在一天当中最有利于进行光合作用。从11:00开始,Pn不同程度地降低,至17:00降至最低值。10、11月份11:00—13:00 Pn的降幅较大,13:00时Pn分别为3.36、1.88 μmol/(m2·s),比11:00分别降低了28.51%、16.07%。这可能与10、11月该时间点光照强度和空气干燥度较大有关。总的来说,在自然降温过程中广玉兰叶片Pn的变化规律,反映了其内在规律和抗寒性。
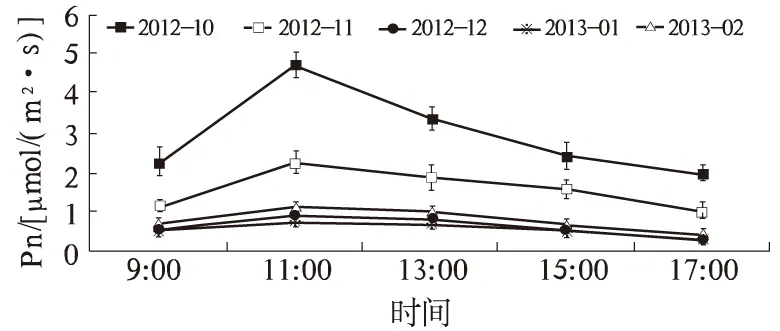
图4 广玉兰不同月份Pn的日变化
2.2.2 Gs 广玉兰叶片的Gs日变化曲线为单峰曲线(图5),与Pn日变化曲线(图4)存在同步性。10月、11月、12月、翌年1月、2月Gs日变化均在11:00达到峰值,分别为0.075、0.046、0.018、0.021、0.025 μmol/(m2·s)。午后,随着气温升高,光照加强,蒸腾失水较多,Gs逐渐下降,于17:00降为最低值。其中,在11:00—17:00 Gs的降低过程中,11:00—13:00的降低幅度较大,13:00—15:00的降幅较平缓。表明广玉兰叶片气孔在早晨打开后一直处于开放状态,11:00—13:00的强光照、高气温使蒸腾作用加强(图6),气孔为避免水分的过度丧失,迅速调低开度,Gs在该时间段明显降低。

图5 广玉兰不同月份Gs的日变化
2.2.3 Tr 图6表明,Tr日变化曲线表现为先升高再降低的单峰曲线,与Pn、Gs日变化曲线大致相似。Pn、Gs日变化峰值出现在11:00,Tr则随着一天中温度的不断升高,于13:00达到峰值。这可能与11:00过后空气光照和气温逐渐强,湿度相对变低,叶片与外部环境的蒸汽压梯度增强,叶片蒸腾作用于13:00左右加剧有关。其后Tr随温度的降低而降低,17:00降为最低值。另外,从Gs日变化中还可看出,10月、11月9:00—13:00 Gs的上升幅度及13:00—17:00的下降幅度均比其他月份明显。

图6 广玉兰不同月份Tr的日变化
2.2.4 Ci 广玉兰叶片Ci日变化呈早晚高而中午低的“V” 字型曲线(图7)。Ci从早晨开始逐渐下降,在13:00左右降为最低值,10月、11月、12月、翌年1月、2月分别为95.16、203.32、270.32、265.89、257.00 μmol/mol,比9:00的Ci值分别降低了50.05%、22.10%、12.96%、10.16%、8.03%。由此可知,广玉兰叶片Ci在9:00—13:00过程中,10月下降幅度较大,11月、12月、1月次之,2月下降幅度最小。13:00之后,随时间推移Ci逐渐升高,但17:00 Ci值仍小于9:00的Ci值。

图7 广玉兰不同月份Ci的日变化
3 结论与讨论
光合作用是植物获得物质和能量的重要代谢过程,受许多环境因子的影响。低温是各种非生物逆境胁迫中对光合作用影响较大的环境因子。关于光合作用对低温胁迫响应的研究较多,一般认为,植物受低温影响时,由于光合机构的破坏、相关酶活性降低等因素,Pn均表现一定程度的下降,植物光合能力降低。杨华庚等[12]研究低温胁迫对油棕幼苗光合作用的影响发现:温度由25 ℃降至4 ℃,气孔因素是油棕幼苗光合作用下降的主要原因;温度下降至1 ℃,光合机构遭受破坏,致使油棕幼苗光合能力降低。周建等[5]、张亚红等[6]认为,低温对光合作用的影响与植物类囊体膜及超氧化物歧化酶等活性的降低有关;曾韶西等[13]在研究低温处理黄瓜幼苗时发现,低温胁迫使子叶中叶绿素含量下降,导致Pn降低;郭延平等[14]对温州蜜柑以及赵军等[15]对水稻的研究表明,低温胁迫导致Rubisco活性下降。本研究中广玉兰在自然降温过程中,叶片Pn明显降低(12月Pn比10月降低了80.58%),但仍然具有一定的光合能力。许大全[16]认为,引起植物光合速率下降的自身因素有气孔部分和非气孔部分2类。随着温度的降低,植物Pn下降,Gs下降,而Ci升高时,Pn降低的主要原因一定是非气孔因素。相反,当Pn下降,Gs下降,Ci也下降时,Gs是Pn降低的主要因素[16]。本研究发现,广玉兰在自然越冬过程中叶片Pn、Gs、Tr均呈现不同程度降低,Ci则随着温度降低而升高。可见,非气孔因素是影响广玉兰叶片Pn下降的主要原因。说明,随着温度的降低,广玉兰叶片的光合机构和影响卡尔文循环的酶受到损伤,进而使叶片的光合活性降低。但广玉兰叶片Pn下降的具体原因还有待于进一步研究。周建等[17]在研究低温胁迫对广玉兰幼苗光合作用的影响时发现,低温胁迫不但导致广玉兰幼苗光合作用的气孔抑制,还使PSⅡ反应中心失活,引起其光能原初捕捉能力和光能同化率减弱,增加了通过热辐射消耗的光能比例,最终使广玉兰幼苗光合能力降低。这个结果与本研究结果有差异,可能与所选广玉兰的树龄、天气状况等有关。
低温胁迫引起广玉兰光合作用下降,但不同的低温对光合作用影响程度不同。本研究发现,在自然降温过程中广玉兰2012年10月、11月Pn高于其他月份,处于较高水平,基本保持在2.20~4.48 μmol/(m2·s)。冬季(12月、1月和2月)Pn处于较低水平,在0.79~1.14 μmol/(m2·s)浮动。说明广玉兰在秋冬季节具有不同的光合能力。另外,广玉兰叶片Pn在10月至12月期间下降幅度较大(12月比10月降低了80.58%)。2012年12月至2013年2月期间Pn降幅较小(1月比12月降低9.19%),这可能与测定时的气温变化有关。
许多树木的Pn在晴天的日变化为双峰曲线,存在光合作用的“午休”现象。本研究测得的2012年10月至2013年2月广玉兰Pn日变化曲线均为先升高再降低的单峰曲线,不存在“午休”现象。说明郑州地区秋冬季节中午时的太阳辐射和气温相对适宜,不足以拉大大气中的水汽饱和差,引起气孔关闭而造成光合作用的“午休”现象。不同月份Pn日变化均在11:00左右出现峰值,其后Pn表现为不同程度的下降趋势,17:00降为最低值。Gs、Tr作为光合作用的重要气态因子,与Pn的日变化相似,均为倒“V”型单峰曲线。Gs日变化在11:00左右出现峰值,与Pn日变化曲线存在同步性。Tr的日变化峰值晚于Pn、Gs,于13:00到达最大值。不同月份Ci日变化均呈“V”字型曲线。从早晨开始逐渐下降,在 13:00左右降为最低值,随后呈不同幅度升高,但17:00的Ci值仍小于9:00的Ci值。
总之,广玉兰叶片在自然降温中的光合响应机制不但对于研究秋冬季节广玉兰的生长、碳的同化量、耐寒性具有十分重要的作用,对于广玉兰向北方地区的引种也提供了重要的理论依据。
[1] Boyer J S.Plant productivity and environment [J].Science,1982,218:443-448.
[2] 汤庚国.树木学[M].北京:中国林业出版社,2005.
[3] 王刚勇.广玉兰在城市绿化中的应用[J].安徽林业,2006 (1):24.
[4] 刘艳萍,朱延林,康向阳,等.不同类型广玉兰的抗寒性评价[J].林业科学,2013,49(6):178-183.
[5] 周建,尤扬,袁德义.低温胁迫对广玉兰生理特性的影响[J].西北林学院学报,2012,23(6):38-42.
[6] 张亚红,孙存华,蒋继宏,等.自然降温过程中香樟和广玉兰的抗寒性研究[J].安徽农业科学,2012,38(16):8653-8654,8657.
[7] Powles S B.Photoinhibition of photosynthesis induced by visible light[J].Annu Rev Plantphysiol,1984,15(1):15-44.
[8] 来芳,宋纪元,王冉,等.广玉兰苗期光合生理特性[J].江苏农业科学,2013,41(11):202-204.
[9] 司建华,常宗强,苏永红,等.胡杨叶片气孔导度特征及其对环境因子的响应[J].西北植物学报,2008,28(1):125-130.
[10] 董智,马宇飞,李红丽,等.4个紫花苜蓿品种分枝期光合速率、蒸腾速率日变化及其影响因子分析[J].中国草地学报,2009,31(3):67-71.
[11] 林琼影,胡剑,温国胜,等.天目山毛竹叶冬季光合作用日变化规律[J].福建林学院学报,2008,28(1):61-64.
[12] 杨华庚,林位夫.低温胁迫对油棕幼苗光合作用及叶绿素荧光特性的影响[J].中国农学通报,2009,25(24):506-509.
[13] 曾韶西,王以柔,刘鸿先,等.低温光照下与黄瓜子叶叶绿素降低有关的酶促反应[J].植物生理学报,1991,17(2):177-182.
[14] 郭延平,张良诚.温州蜜柑叶片气体交换和叶绿素荧光对低温的响应[J].植物生理学报,2000,26(2):88-94.
[15] 赵军,王以柔.低温锻炼对水稻叶片中Rubisoc的影响[J].植物生理学报,1997,23(2):123-129.
[16] 许大全.光合作用“午睡”现象的生态、生理与生化[J].植物生理学通讯,1990(6):5-10.
[17] 周建,杨立峰,郝峰鸽,等.低温胁迫对广玉兰幼苗光合及叶绿素荧光特性的影响[J].西北植物学报,2009,29(1):136-142.
Photosynthetic Response Mechanism ofMagnoliagrandifloraunder Natural Temperature Falling
CHEN Lipei1,WANG Guoxia1,YANG Yuzhen1*,LIU Yanping2,LIU Ruixia1
(1.School of Life Sciences,Zhengzhou Normal University,Zhengzhou 450044,China; 2.Henan Academy of Forestry,Zhengzhou 450008,China)
A research about photosynthetic response mechanism was studied by measuring the changes of Pn,Gs,Tr,Ci by Li-6400 photosynthesis meter on the 15-year-oldMagnoliagrandifloraunder natural temperature falling in autumn and winter.The results showed that Pn,Gs,Tr ofMagnoliagrandifloradecreased,while Ci increased along with the decline of mean temperature.In addition,the leaves ofMagnoliagrandiflorastill had a certain ability of photosynthesis during the temperature droped,and which appeared an obvious daily change rule,the Pn,Gs,Tr showed a single curve of first increase and then decrease,while Ci showed V-shaped curve.Moreover,Pn had no midday depression.
Magnoliagrandiflora; natural temperature falling; photosynthetic rate; stomatal conduc-tance; response mechanism
2015-06-27
国家林业局“948”项目(2014-4-12);河南省教育厅科学技术研究重点项目(14B180037);郑州市科学技术攻关项目(141PPTGG426)
陈丽培(1981-),女,河南郑州人,讲师,硕士,主要从事森林培育研究。E-mail:chenlp0524@126.com
*通讯作者:杨玉珍(1965-),女,河南郑州人,教授,博士,主要从事森林培育研究。
S685.15
A
1004-3268(2016)01-0114-05
猜你喜欢
作文与考试·小学高年级版(2023年8期)2023-07-17 09:32:52
科学(2022年4期)2022-10-25 02:43:00
干旱地区农业研究(2022年1期)2022-01-28 04:51:30
装备制造技术(2019年12期)2019-12-25 03:06:52
科技创新与品牌(2019年12期)2019-02-06 04:03:29
中国诗歌(2018年5期)2018-11-14 20:52:30
作文评点报·低幼版(2018年30期)2018-09-27 12:21:24
作文评点报·低幼版(2018年30期)2018-09-27 12:21:24
Coco薇(2016年5期)2016-06-03 09:17:41
考试周刊(2016年6期)2016-03-11 08:14:32
