
认知重评阻断条件化恐惧记忆的习得与表达
——对恐惧反应的长程抑制作用*
2016-02-01 04:30安献丽陈四光
心理学报 2016年10期
安献丽 陈四光
(扬州大学教育科学学院, 扬州 225002)
1 引言
恐惧是人面临威胁时一种适应性的以保护自我的情绪反应。但是, 由具有生命威胁性的应激所引起的不可控制的恐惧反应可能会导致病理性的精神障碍。因此, 探索如何阻断或破坏恐惧情绪的表达是研究者数百年来一直所努力的方向。在众多抑制恐惧表达的方法中, 无伤害性、临床效度高的行为方法备受推崇, 其中行为消退(extinction)得到了最广泛的研究。不幸的是, 大量研究发现条件化恐惧的消退比较微弱且依赖于特定环境(Bouton, ann,2008)。而人在应对应激时, 注意、意识、语言及行为监控等高级认知活动均会参与其中(Delgado, Olsson,& Phelps, 2006)。因此, 常用于动物研究的简单行为消退可能并不能很好地抑制人的恐惧反应, 而高级的、有认知参与的情绪调控技术可能更有效。近年来, 认知情绪调节(Cognitive emotion regulation)逐渐成为了研究者所关注的焦点。
情绪调节指个体理解自己情绪状态, 并主动努力通过一定的策略改变自己不适应的情绪, 最终产生适应性行为的能力。研究发现, 积极有效的情绪调节能够改善人际关系和工作成绩(John & Gross,2004); 反之, 情绪调节困难则会导致精神障碍, 包括抑郁症(Rottenberg, Gross, & Gotlib, 2005)、焦虑症(Mennin, McLaughlin, & Flanagan, 2009)、物质成瘾(Berking et al., 2011)等。Gross通过因素分析把情绪调节策略归为两大类:认知重评和表达抑制。认知重评是从认知上对情绪性刺激或事件的意义进行重新解释以改变情绪体验的策略。表达抑制指对将要发生或正在发生的情绪表达进行抑制, 情绪体验并没有因此改变, 反而会增强情绪唤醒反应(Gross,1998)。因此, 在抑制消极情绪唤醒反应时, 认知重评具有一定的优势(程利, 袁加锦, 何媛媛, 李红,2009)。本研究首先关注认知重评是否能够有效地抑制恐惧反应。
研究发现, 认知重评能够改变个体对负性刺激的情绪唤醒程度, 包括自我报告、生理和神经反应。如给被试呈现负性情绪图片后, 要求被试重新对图片信息进行积极的解释以下调情绪, 就能够显著降低被试对情绪性图片的皮肤电及心率反应(Driscoll,Tranel, & Anderson, 2009), 且能够有效地减少消极情绪体验(Eippert et al., 2007)。并且, 脑功能成像研究发现对消极信息的认知重评能够显著增加外侧前额叶皮层(lPFC)和内侧前额叶皮层(mPFC)的活动、降低杏仁核的激活(Ochsner, Silvers, & Buhle,2012)。PFC的激活程度增加及杏仁核激活程度的降低均有利于抑制消极情绪的反应(Marek, Strobel,Bredy, & Sah, 2013)。因此, 有研究者认为, 认知重评可以通过影响大脑的活动而消除恐惧(Delgado,Nearing, LeDoux, & Phelps, 2008)。
认知重评用于降低条件化恐惧反应的研究还比较缺乏, Delgado等(2008)开展了这项实验。他们在条件化恐惧训练过程中, 在电脑屏幕上给被试呈现两种不同的条件化刺激(conditioned stimulus, CS):蓝色方块和黄色方块, 并告诉被试, 当屏幕上呈现蓝色方块时, 他们可能会受到一次电击(CS+); 而屏幕上出现黄色方块时, 就不会受到电击(CS−)。在每个CS出现前呈现一个线索词, 用于指导被试对呈现的CS是否采取“调节” (regulate)的策略。当不要求被试进行认知重评时, 被试可以自然地感受自己的情绪反应; 而当要求被试进行认知重评时, 被试应该积极地对自己的情绪状态进行认知调节, 即通过想象一些可以使自己平静的自然风景来改变自己的情绪状态。结果发现, 在训练过程中, 认知重评可以有效地降低被试对CS+的恐惧反应。但是,Delgado等的研究侧重于认知重评与行为消退在调控恐惧反应时的神经活动比较, 而没有考察其恐惧抑制作用能否长久持续存在。
对恐惧记忆的大量研究证明, 如果不能破坏恐惧记忆的痕迹, 对恐惧反应的抑制就只能是暂时的,若经历再次应激、改变新的环境或随时间的推移,恐惧反应就会复发(参见综述(Myers & Davis, 2002))。那么, 认知重评能否破坏恐惧反应的复发, 实现长久抑制恐惧反应的目的呢?与Delgado等的研究相似, 对认知重评调控人类情绪的研究大多数以调控当时的情绪反应为指标, 很少有研究探索其长期效果, 更缺乏对预防恐惧反应复发的研究。如研究者发现, 认知重评可以在15分钟内保持对情绪反应的调控(Erk et al., 2010), 但其长期效应还未知。而无论对应激相关障碍的前临床研究还是临床治疗, 一直存在恐惧反应难以消除而反复出现或应激相关障碍疗效较差而反复复发的问题。基于认知重评能够影响与恐惧相关的脑区活动, 且能够降低相应的恐惧反应, 本研究以恐惧记忆的自发恢复测试为关键指标, 分别考察在恐惧记忆习得与恐惧记忆表达过程中进行认知重评对恐惧反应的长时程表达的影响。
2 实验1:恐惧记忆习得阶段进行认知重评对条件化恐惧的影响
2.1 被试
最初被试为34名扬州大学在读本科生。在最终统计中, 删除了4名被试。根据被试对CS+与CS−的皮肤电反应, 删除了自然感受组2名未能获得条件化恐惧反应的被试, 即他们测试一中的前两个CS+− CS− < 0 μs。又根据认知重评组自我报告的情绪调节成功度是否低于50%, 删除了此组2名未能成功进行认知重评的被试。最终被试30名, 其中男生15名(平均年龄20.80 ± 0.86岁), 女生15名(平均年龄20.20 ± 0.56岁)。
2.2 生理信号记录
皮肤电反应(Skin conductance response, SCR)是本实验主要测量的条件化恐惧反应指标。实验数据通过Biopac MP150 (Biopac Inc., Santa Barbara CA)系统的GSR 100C放大器采集, 采样率250 Hz。数据记录时将预先填充了GEL 100电极胶的Ag-AgCl生物传感器(TSD 203)连接到被试非利手的食指与中指最前端指节指腹侧。SCR数据分析通过Acqknowledge 4.2软件实现。SCR值指CS呈现开始后的0.5~6.5 s内被试皮肤电阻反应的峰−峰值(P-P值), 但如果CS出现时SCR水平持续下降而无上升峰就定义为0。为了使SCR数据趋于正态分布, 最终将原始数据取平方根进行转换。最后, 将转换后的每一个CS+的SCR值减去CS−的SCR均值。
2.3 实验程序
采用部分强化的条件化恐惧范式进行实验,Superlab心理实验设计系统用于刺激的呈现。全部实验包括3个阶段:恐惧记忆习得、恐惧记忆表达(测试一)与恐惧记忆自发恢复(测试二)测试。在习得开始前, 先让被试平静5分钟, 其间采集被试的皮肤电水平基线值。在习得过程中, 先利用指导语将被试随机地分配到认知重评组(n=15)和自然感受组(n=15)进行条件化恐惧习得训练。条件化刺激(conditioned stimulus, CS)为呈现6 s的黄色和蓝色的方块, 分别作为CS+与CS−; 非条件化刺激(unconditioned stimulus,US)为持续750 ms的100 dB强噪音。根据Indovina,Robbins, Núñez-Elizalde, Dunn和Bishop (2011)的研究, 利用强噪音作为US是可以很好地建立条件化恐惧反应的(Indovina et al., 2011)。条件化恐惧获得过程共呈现52次CS, 这52次CS被分成两部分,每部分26次, 中间间隔60 s。在每部分的26次CS中,CS+呈现16次, 其中10次CS+后未伴随US, 6次CS+与强噪音匹配呈现, 即CS+呈现的最后750 ms开始呈现US; CS−呈现10次, 呈现时不伴随强噪音。CS+与CS−随机呈现, CS间呈现间隔10~20 s, 平均15 s。对于每一个被试, 一种特定的色块(如黄色)为CS+, 另一种(如蓝色)为CS−, 作为CS+或CS−的色块颜色在被试间平衡。在恐惧习得阶段, 参照Delgado等的方法(Delgado et al., 2008), 对于自然感受组,指导被试在CS呈现时自然地观看和感受刺激图片及其所诱导的情绪感受; 对于认知重评组, 主要指导被试如何使用认知重评技术, 即CS呈现时通过努力想象以降低图片所诱导的情绪反应。
习得训练开始前指导被试如何进行认知重评,直到被试完全掌握了该技术。即认知重评时要求想象与图片颜色相似的、能让人平静的自然风光。如黄色图片呈现时想象阳光、成片的向日葵、黄色的花等; 蓝色图片呈现时想象蓝天、蓝色大海等。要求被试在习得过程中尽量保持想象与色块颜色对应的同一副风景, 即最能够让其平静的风景。习得训练后, 让所有被试写下CS+的颜色, 并对自己判断的确信度从0%~100%评分, 最终显示所有被试都能够正确判断CS+的颜色, 且确信度均高于50%。另外, 认知重评组还要对自己进行情绪调节的成功度进行0%~100%评分。
条件化恐惧习得后24 h进行表达测试(测试一),即随机呈现10次CS+与10次CS−, 不呈现US, CS间间隔10~20 s。表达测试30天后, 恐惧记忆自发恢复测试(测试二), 过程与测试一相同。
2.4 数据统计
基线皮肤电水平(Skin conductance level, SCL)指条件化恐惧习得开始前5分钟的皮肤电反应均值。CS与US呈现后的0.5~6.5 s的SCR值分别为条件化与非条件化恐惧反应。为了能够将被试对CS的条件化恐惧反应与对US的非条件化恐惧反应区分开, 恐惧获得阶段未统计匹配US的CS+。本研究统计的恐惧水平指CS+减CS−的SCR差值。但在求这一SCR差值时, 先将每一阶段对CS-的SCR反应求均值, 再用每一个CS+的SCR值减去这个均值。最后,将每两个SCR差值平均后作为一个组块(block)进行分析。组间SCR基线水平比较采用独立样本t检验; 认知重评对恐惧记忆的习得、表达和自发恢复的影响, 均采用组别(认知重评和自然感受)×CS呈现过程的重复测量两因素方差分析。对恐惧记忆的自发恢复, 还对两组测试一的B5(最后一个组块)与测试二的B1(第一个组块)进行组别(认知重评和自然感受)×测试(测试一和测试二)两因素方差分析。事后检验采用Bonferroni方法矫正。
2.5 实验结果
首先, 为了防止被试的基础皮肤电水平对条件化恐惧习得造成影响, 先考察在未给予任何刺激的情况下, 自然感受与认知重评两组被试的基础SCL是否相同。独立样本t检验显示, 两组间不存在显著差异,M自然感受=1.66,SE=0.09;M认知重评=1.70,SE=0.12;t(28) =0.27,p=0.79, Cohen’d=0.10。因此, 基线SCL高低可能不会影响恐惧习得与测试过程中SCR的组间差异。
自然感受组与认知重评组在习得过程中呈现出了不同的反应趋势(图1A)。组别(认知重评和自然感受)×CS呈现过程两因素重复测量方差分析发现, 在习得过程上存在显著的组别主效应,F(1, 28)=7.74,p< 0.01, η2=0.22, 及显著的CS呈现过程主效应,F(9, 252)=3.21,p=0.001, η2=0.10。事后检验发现, 在B9与B10上存在显著的组间差异,t(280)=3.30,p=0.011;t(280)=3.09,p=0.023, 说明在恐惧记忆习得过程中进行认知重评有效地降低了个体的条件化恐惧反应。
如果认知重评是通过阻断恐惧记忆的习得而降低恐惧反应的, 那么认知重评也必然会降低恐惧记忆的表达与自发恢复水平。组别×CS呈现过程两因素方差分析显示(图1B), 在恐惧记忆测试一中存在显著的组别主效应,F(1, 28)=5.26,p< 0.05, η2=0.16, 但组别与CS呈现过程之间的交互效应不显著。事后检验显示, 在前两个CS呈现时(B1)存在显著的组间差异,t(140)=2.92,p< 0.05, 提示认知重评破坏了恐惧记忆的表达。两因素方差分析还发现, 测试一的SCR水平存在极其显著的CS呈现过程主效应,F(4, 112)=10.39,p< 0.001, η2=0.27。说明随着CS的呈现, 被试的条件化恐惧反应逐渐地降低。事后检验发现, 在自然感受组, B1的SCR水平极其显著地高于B3、B4和B5,t(112)=3.43,p< 0.01;t(112)=5.13,p< 0.001;t(112)=3.81,p< 0.001; 在认知重评组, B1的SCR水平也极其显著地高于B4,t(112)=3.19,p< 0.01。在测试一的最后几个CS, 两组被试的条件化恐惧反应均降到了非常低的水平,两组之间无差异。
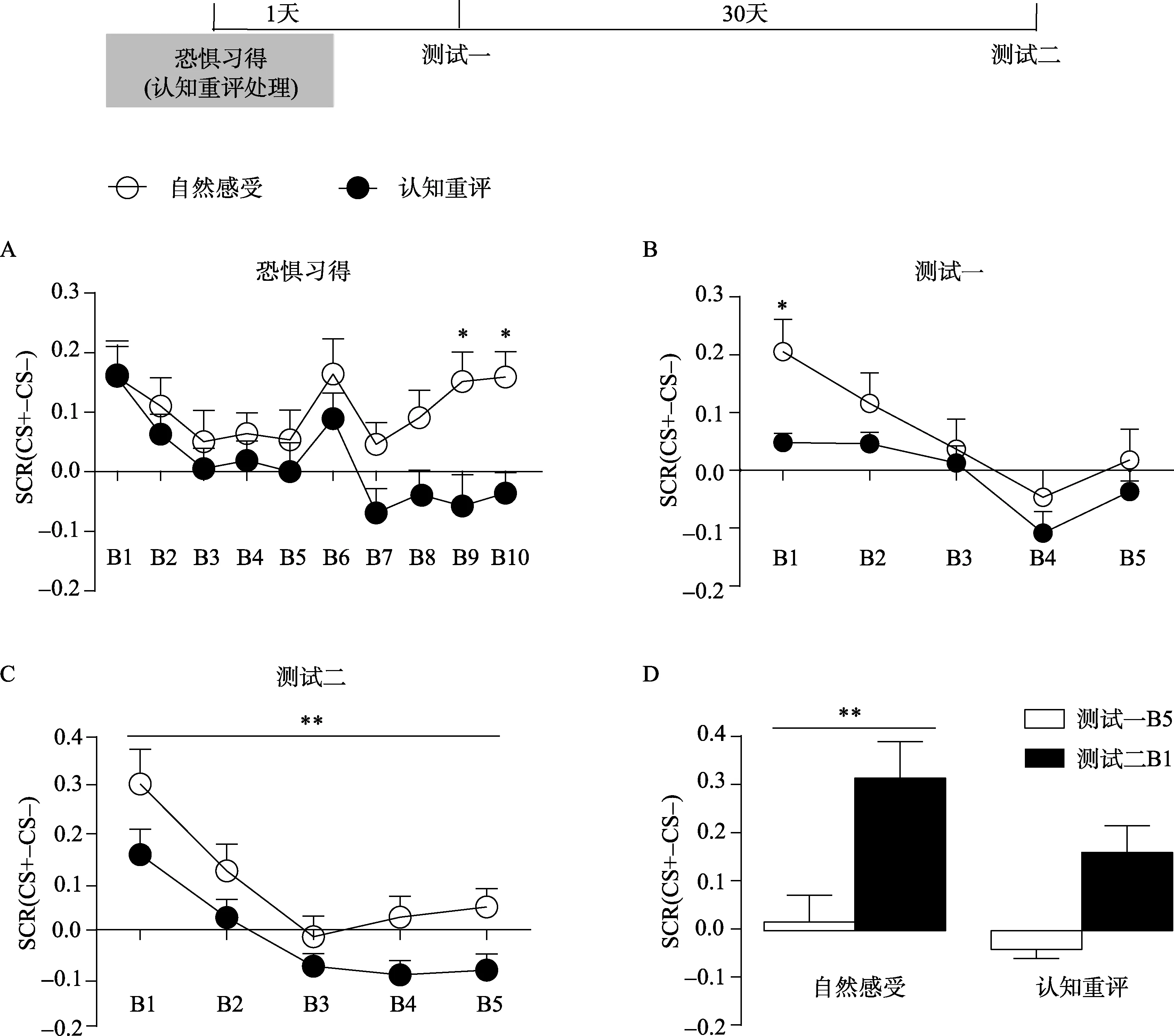
图1 习得阶段进行认知重评对条件化恐惧反应的影响(Mean ± SE)。*p < 0.05, **p < 0.01
同样, 组别×CS呈现过程两因素方差分析也显示(图1C), 在恐惧记忆测试二中存在极其显著的CS呈现过程主效应,F(4, 112)=14.67,p< 0.001, η2=0.34。即随着CS的呈现, 被试的条件化恐惧反应逐渐地降低。事后检验发现,在自然感受组, B1的SCR水平极其显著地高于B2、B3、B4和B5,t(112)=3.00,p< 0.01;t(112)=5.27,p< 0.001;t(112)=4.59,p<0.001;t(112)=4.23,p< 0.001; 在认知重评组, B1的SCR水平也显著地高于B3、B4和B5,t(112)=3.86,p< 0.01;t(112)=4.15,p< 0.001;t(112)=3.99,p< 0.01。这说明两组在测试二开始时均有一定程度的恐惧表达。那么, 两组的恐惧有没有显著地自发恢复呢?两因素反差分析还发现, 虽然组别与CS呈现过程不存在显著的交互效应, 但测试二的SCR水平存在显著的组别主效应,F(1, 28)=10.29,p=0.003, η2=0.27。这说明在测试二, 认知重评组的SCR水平极其显著地低于自然感受组。并且, 利用组别(认知重评和自然感受)×测试(测试一和测试二)两因素方差分析将测试二的B1与测试一的B5进行分析(图1D), 发现存在显著的组别与测试主效应,F(1, 28)组别=4.41,p< 0.05, η2=0.14;F(1, 28)测试=17.13,p< 0.001, η2=0.38, 但二者交互效应不显著。事后检验显示, 在自然感受组, 测试二B1的SCR水平极其显著地高于测试一的B5,t(28)=3.49,p< 0.01; 但在认知重评组, 测试二的B1与测试一的B5不存在显著差异,t(28)=2.36,p> 0.05。这提示, 认知重评有效地降低了恐惧记忆的自发恢复。
3 实验2:恐惧记忆表达阶段进行认知重评对条件化恐惧的影响
3.1 被试
最初被试为34名扬州大学在读研究生与本科生。在最终统计中, 删除了6名被试。首先删除4名未能获得条件化恐惧反应的被试, 即他们测试一中的前两个CS+− CS−< 0 μs。又根据被试在恐惧习得后自我报告对CS+颜色判断的确信度, 删除了1名确信度等于50%的被试。最后, 删除1名报告了错误CS+颜色的被试。最终被试28名, 其中男生13名(平均年龄20.54 ± 1.85岁), 女生15名(平均年龄23.27 ± 2.22岁)。
3.2 生理信号记录
同实验1。
3.3 实验程序
条件化恐惧的习得、测试一、测试二等基本过程与实验1相同。不同的是, 在条件化恐惧习得过程中, 所有的被试都是在自然感受的条件下接受条件化恐惧训练。在表达测试时(测试一), 再随机将被试分为认知重评组(n=14)和自然感受组(n=14)。在测试一开始前, 认知重评组接受认知重评技术的训练, 并在测试过程中采用此技术来调控自己的情绪反应。对于自然感受组, 不接受认知重评技术的训练, 需在测试一过程中自然地感受刺激呈现所带来的情绪感受。
与实验1相同, 条件化恐惧习得训练后, 让所有被试写下CS+的颜色, 并对自己判断的确信度从0%~100%评分。在测试一后, 要求认知重评组对自己情绪调节的成功度进行0%~100%评分。
3.4 数据统计
同实验1
3.5 实验结果
图2A所示, 自然感受组与认知重评组在习得过程中具有相同的SCR水平。组别(认知重评和自然感受)×CS呈现过程两因素重复测量方差分析显示, 不存在显著的组别主效应,F(1, 26)=0.34,p=0.566。这提示两组在测试一、测试二中的恐惧反应差异并不是由于习得恐惧水平的不同造成的。
在表达测试(测试一)时进行认知重评实验处理,组别×CS呈现过程两因素方差分析显示(图2B), 存在显著的组间主效应,F(1, 26)=4.77,p=0.038, η2=0.15; 以及显著的CS呈现过程主效应,F(4, 104)=7.21,p< 0.001, η2=0.22, 说明随着CS的呈现, 被试的条件化恐惧反应逐渐地降低。Bonferroni事后检验发现, 在B1上组间差异不显著, 但在B2上存在边缘显著的组间差异,t(130)=2.36,p=0.098。由于在表达测试过程中被试的恐惧反应逐渐降低, 即出现了恐惧反应的消退, 这提示自然感受组与认知重评组具有不同的消退速度, 即认知重评促进了恐惧反应的消退。在测试一的最后几个CS, 两组被试的条件化恐惧反应均降到了非常低的水平, 两组之间无差异。

图2 表达阶段(测试一)进行认知重评对条件化恐惧反应的影响(Mean ± SE)。*p < 0.05
为了进一步考察认知重评是否能够通过影响恐惧记忆的表达而抑制其复发, 对条件化恐惧自发恢复测试(测试二)进行组别×CS呈现过程两因素方差分析(图2C), 结果显示, 在恐惧记忆测试二中虽然不存在显著的组间及CS呈现过程主效应, 但二者之间存在显著的交互效应,F(4, 104)=2.86,p<0.05, η2=0.10。Bonferroni事后检验显示, 在测试二, 认知重评组B1的SCR水平显著地低于自然感受组,t(130)=2.70,p< 0.05。事后检验还显示, 在自然感受组, B1的SCR水平极其显著地高于B4,t(104)=3.92,p< 0.01; 而在认知重评组, CS呈现过程中被试的恐惧反应始终处于较低的水平, 未显示出对CS反应的差异。进一步利用组别(认知重评和自然感受)×测试(测试一和测试二)两因素方差分析对测试二的B1与测试一的B5进行分析(图2D), 发现存在极其显著的测试主效应,F(1, 26)=8.93,p<0.01, η2=0.26, 以及显著的组别主效应,F(1, 26)=10.49,p< 0.01, η2=0.29。事后检验显示, 在自然感受组, 测试二B1的SCR水平边缘显著地高于测试一的B5,t(26)=2.20,p=0.074; 在认知重评组, 测试二的B1与测试一的B5无显著差异,t(26)=2.03,p> 0.05。以上结果均提示, 在恐惧记忆表达阶段进行认知重评也能够实现对恐惧反应的长程抑制。
4 讨论
本研究主要考察了认知重评对人类条件化恐惧记忆习得与表达的影响。我们首先发现, 在恐惧习得过程中进行认知重评一方面降低了习得过程中被试的恐惧反应, 另一方面还有效地抑制了习得后24 h恐惧记忆的表达及1个月后恐惧记忆的自发恢复。这说明认知重评可能能够通过破坏条件化恐惧记忆的习得而达到长久抑制恐惧表达的目的。
虽然利用认知重评来降低条件化恐惧反应的研究还比较缺乏, 但是对情绪性图片或情绪性视频的大量研究在某种程度上支持我们的研究结果。这些研究给被试呈现负性情绪图片或情绪性视频片段后, 要求被试重新对图片或视频信息进行积极的解释以降低他们的消极情绪反应, 发现认知重评可以有效地减少消极的主观情绪感受及相应的生理反应, 如皮肤电、皱眉肌肌电反应等(Driscoll et al.,2009; 原琳, 彭明, 刘丹玮, 周仁来, 2011)。Driscoll等(2009)发现了被试的恐惧反应可以被情绪调节策略所抑制。他们先给被试呈现积极或消极的情绪图片,其中部分图片伴随95分贝的噪音探测刺激, 要求被试在实验过程中进行情绪调节或自然感受, 结果发现情绪调节显著地降低了图片呈现过程中被试的眨眼惊吓、皮肤电及心率反应, 这提示利用认知解释可以有效地抑制个体的恐惧反应。情绪调节理论认为, 认知重评是一种先行聚焦策略, 这种策略能够在情绪反应被充分激活之前通过认知来改变情绪性事件的意义而达到降低消极的情绪反应(Gross,2002)。因此, 从理论上可以推测, 由条件化恐惧训练、情绪图片及情绪视频等引起的消极情绪均能够被认知重评所抑制。最近一项以蜘蛛或蛇为CS, 以微弱电刺激为US进行条件化恐惧训练的研究发现,认知重评有效地抑制了对CS+的恐惧反应, 且同时抑制了应激后皮质醇及去甲肾上腺素活动的产物α-淀粉酶的释放(Raio, Orederu, Palazzolo, Shurick, &Phelps, 2013)。这两种物质在应激事件发生后的大量释放会破坏PFC的功能而导致个体恐惧记忆的形成(Arnsten, 2009)。因此, Raio等(2013)认为, 很可能在条件化恐惧习得过程中, 认知重评抑制了应激激素的释放而保护了对PFC功能的损害。而PFC在应激过程中能够通过抑制杏仁核的活动阻断个体的恐惧表达(Rosenkranz & Grace, 2002)。由于杏仁核是条件性恐惧获得与巩固的关键脑区, 因此,在认知重评条件下, 被试的PFC能够有效地控制杏仁核的激活, 从而破坏了对恐惧信息的加工编码。
Ochsner等(2012)根据以往研究总结认为, 认知重评能显著增加负性情绪刺激诱导的PFC活动,并同时降低杏仁核的激活。Eippert等(2007)让被试在观看威胁性图片时进行认知重评也发现, 被试的杏仁核活动能够被显著地抑制, 且这种抑制效应依赖于PFC是否被激活。同样, 善于使用认知重评策略的个体在加工负性面部表情时的PFC活动更强,而杏仁核的激活更低(Drabant, McRae, Manuck, Hariri,& Gross, 2009)。Olsson和Phelps (2007)认为, 可以通过对应激事件的认知解释、进而影响杏仁核的活动来改变个体的恐惧反应水平。因此, 虽然我们未能对认知重评抑制恐惧习得及表达的神经机制进行研究, 但从以往研究我们可以推论, 认知重评很可能通过改变了条件化恐惧习得过程中相关的神经活动而破坏了对恐惧信息的编码, 从而导致恐惧记忆在短程及长程表达中均得到了显著的抑制。
另外, 我们还发现, 在恐惧记忆表达过程中进行认知重评在降低恐惧表达的同时也显著抑制了1个月后恐惧记忆的自发恢复, 这也可能是因为在表达阶段进行认知重评抑制了杏仁核的活动并同时增强了PFC的激活程度引起的。Delgado等(2008)通过对条件化恐惧训练过程中认知重评的研究发现, 认知重评所造成的被试神经系统活动的改变与消退过程相似, 即背外侧及腹内侧PFC的活动增强而杏仁核激活程度降低。在条件化恐惧表达测试过程中, 被试的恐惧水平随CS的呈现逐步降低, 这相当于一次消退训练。而PFC激活水平的增加能够促进消退过程。因此, 在表达阶段进行认知重评很可能是由于改变了个体神经系统的活动而促进了被试的消退学习, 最终导致了对恐惧反应的长时程抑制。
记忆理论也支持我们的上述发现。从记忆理论角度来说, 记忆习得先于对记忆信息的编码巩固,因此, 干预条件化恐惧记忆的习得就能够破坏恐惧记忆在大脑中的编码, 之后的恐惧记忆也不能得到有效表达(Heath et al., 2015; Yang, Li, & Huang, 2013)。并且, 由于恐惧记忆的巩固在习得后会即刻发生(Abel & Lattal, 2001), 那么在习得过程中进行认知重评, 很可能会使认知重评的作用延伸到巩固阶段,而对记忆巩固的阻断同样会抑制恐惧记忆的长程表达。与恐惧记忆的习得不同, 干预条件化恐惧记忆的表达是在习得后24 h, 此时恐惧记忆已得到巩固, 但这并不意味着巩固记忆痕迹就无法改变。首先, 我们的研究发现在表达阶段进行认知重评很可能是通过促进恐惧反应的消退起效的。而增强恐惧记忆的消退就能够抑制恐惧记忆的自发恢复(Mao et al., 2013)。其次, 由于CS的短暂呈现会导致恐惧记忆的再巩固。在我们的实验中CS是在一定时间间隔下逐步呈现的。虽然CS的多次呈现导致了恐惧反应的消退, 但是, 在前几个CS初次呈现时, 有可能导致恐惧记忆的再巩固。而在这个过程中认知重评已经在起作用, 那么恐惧记忆的再巩固就有可能被抑制。而对再巩固的破坏同样会导致对恐惧反应长程表达的阻断(Nader, 2015)。当然, 这一机制目前还无法区分认知重评对恐惧消退过程的影响。另外,由于认知重评技术的使用能够成为习惯化的行为方式(Hartley & Phelps, 2010), 个体在恐惧表达阶段使用此策略后, 在再次面对条件化刺激后可能会习惯化地使用认知重评技术, 从而能够部分抑制恐惧记忆的长程表达。
5 小结
面对应激时该如何减少负面情绪来保持健康的心理状态是研究者以及普通大众所关心的重要话题。虽然众多研究都发现认知重评能够有效地减少消极情绪, 但以往的认知重评相关研究一般都关注于对信息比较丰富的、不同场景的情绪图片或情绪视频的情绪反应, 要求被试对呈现在面前的、场景不同的图片或视频重新进行意义解释。这些研究一方面缺乏对高应激场景的探索, 另一方面与临床实践相距较远。Delgado等(2008)认为, 利用经典的条件化恐惧模型进行认知重评研究, 要求被试进行调节的刺激是反复呈现的同一个刺激, 而这一刺激在初始时不具有任何情绪意义, 被试进行认知重评时需要进行想象才能改变自己的情绪反应。而基于想象的认知重评过程与传统的认知行为疗法非常接近。临床研究确实发现, 利用认知重评下调情绪不但能够促进心理治疗过程中患者恐惧水平的下降, 还能够增强暴露疗法的疗效, 阻断被试恐惧情绪的复发(Kamphuis & Telch, 2000)。Pile, Barnhofer和Wild (2015)对健康成人的研究也证明, 改变对应激信息的评价可以有效地降低被试的PTSD症状。我们的实验从基础研究角度证明, 积极地通过想象来进行认知重评可以有效地降低应激过程中及恐惧情绪表达过程中个体的恐惧反应, 且这种效应能够在较长的时间内持续存在。这提示在面对应激时,个体如果能够成功地利用想象进行认知重评, 就有可能避免应激事件对个体的心理或精神造成伤害。
Abel, T., & Lattal, K. M. (2001). Molecular mechanisms of memory acquisition, consolidation and retrieval.Current Opinion in Neurobiology, 11(2), 180−187.
Arnsten, A. F. T. (2009). Stress signalling pathways that impair prefrontal cortex structure and function.Nature Reviews Neuroscience, 10(6), 410−422.
Berking, M., Margraf, M., Ebert, D., Wupperman, P., Hofmann,S. G., & Junghanns, K. (2011). Deficits in emotion- regulation skills predict alcohol use during and after cognitive- behavioral therapy for alcohol dependence.Journal of Consulting &Clinical Psychology, 79(3), 307−318.
Bouton, M. E. (2004). Context and behavioral processes in extinction.Learning & Memory, 11(5), 485−494.
Cheng, L., Yuan, J. J., He, Y. Y., & Li, H. (2009). Emotion regulation strategies: Cognitive reappraisal is more effective than expressive suppression.Advances in Psychological Science, 17(4), 730−735.
[程利, 袁加锦, 何媛媛, 李红. (2009). 情绪调节策略: 认知重评优于表达抑制.心理科学进展, 17(4), 730−735.]
Delgado, M. R., Nearing, K. I., LeDoux, J. E., & Phelps, E. A.(2008). Neural circuitry underlying the regulation of conditioned fear and its relation to extinction.Neuron,59(5), 829−838.
Delgado, M. R., Olsson, A., & Phelps, E. A. (2006). Extending animal models of fear conditioning to humans.Biological Psychology, 73(1), 39−48.
Drabant, E. M., McRae, K., Manuck, S. B., Hariri, A. R., &Gross, J. J. (2009). Individual differences in typical reappraisal use predict amygdala and prefrontal responses.Biological Psychiatry, 65(5), 367−373.
Driscoll, D., Tranel, D., & Anderson, S. W. (2009). The effects of voluntary regulation of positive and negative emotion on psychophysiological responsiveness.International Journal of Psychophysiology, 72(1), 61−66.
Eippert, F., Veit, R., Weiskopf, N., Erb, M., Birbaumer, N., &Anders, S. (2007). Regulation of emotional responses elicited by threat-related stimuli.Human Brain Mapping, 28(5),409−423.
Erk, S., Mikschl, A., Stier, S., Ciaramidaro, A., Gapp, V.,Weber, B., & Walter, H. (2010). Acute and sustained effects of cognitive emotion regulation in major depression.Journal of Neuroscience, 30(47), 15726−15734.
Gross, J. J. (1998). Antecedent- and response-focused emotion regulation: Divergent consequences for experience, expression,and physiology.Journal of Personality and Social Psychology, 74(1), 224−237.
Gross, J. J. (2002). Emotion regulation: Affective, cognitive,and social consequences.Psychophysiology, 39(3), 281− 291.
Hartley, C. A., & Phelps, E. A. (2010). Changing fear: The neurocircuitry of emotion regulation.Neuropsychopharmacology,35(1), 136−146.
Heath, F. C., Jurkus, R., Bast, T., Pezze, M. A., Lee, J. L. C.,Voigt, J. P., & Stevenson, C. W. (2015). Dopamine D1-like receptor signalling in the hippocampus and amygdala modulates the acquisition of contextual fear conditioning.Psychopharmacology, 232(14), 2619−2629.
Hofmann, S. G. (2008). Cognitive processes during fear acquisition and extinction in animals and humans: Implications for exposure therapy of anxiety disorders.Clinical Psychology Review, 28(2), 199−210.
Indovina, I., Robbins, T. W., Núñez-Elizalde, A. O., Dunn, B.D., & Bishop, S. J. (2011). Fear-conditioning mechanisms associated with trait vulnerability to anxiety in humans.Neuron, 69(3), 563−571.
John, O. P., & Gross, J. J. (2004). Healthy and unhealthy emotion regulation: Personality processes, individual differences,and life span development.Journal of Personality, 72(6),1301−1334.
Kamphuis, J. H., & Telch, M. J. (2000). Effects of distraction and guided threat reappraisal on fear reduction during exposure-based treatments for specific fears.Behaviour Research and Therapy, 38(12), 1163−1181.
Mao, S. C., Chang, C. H., Wu, C. C., Orejanera, M. J., Manzoni,O. J., & Gean, P. W. (2013). Inhibition of spontaneous recovery of fear by mGluR5 after prolonged extinction training.PLoS One, 8(3), e59580.
Marek, R., Strobel, C., Bredy, T. W., & Sah, P. (2013). The amygdala and medial prefrontal cortex: Partners in the fear circuit.The Journal of Physiology, 591(10), 2381−2391.
Mennin, D. S., McLaughlin, K. A., & Flanagan, T. J. (2009).Emotion regulation deficits in generalized anxiety disorder,social anxiety disorder, and their co-occurrence.Journal of Anxiety Disorders, 23(7), 866−871.
Myers, K. M., & Davis, M. (2002). Behavioral and neural analysis of extinction.Neuron, 36(4), 567−584.
Nader, K. (2015). Reconsolidation and the dynamic nature of memory.Cold Spring Harbor Perspectives in Biology,7(10), a021782.
Ochsner, K. N., Silvers, J. A., & Buhle, J. T. (2012). Functional imaging studies of emotion regulation: A synthetic review and evolving model of the cognitive control of emotion.Annals of the New York Academy of Sciences, 1251, E1− E24.
Olsson, A., & Phelps, E. A. (2007). Social learning of fear.Nature Neuroscience, 10(9), 1095−1102.
Pile, V., Barnhofer, T., & Wild, J. (2015). Updating versus exposure to prevent consolidation of conditioned fear.PLoS One, 10(4), e0122971.
Raio, C. M., Orederu, T. A., Palazzolo, L., Shurick, A. A., &Phelps, E. A. (2013). Cognitive emotion regulation fails the stress test.Proceedings of the National Academy of Sciences of the United States of America, 110(37), 15139−15144.
Rosenkranz, J. A., & Grace, A. A. (2002). Cellular mechanisms of infralimbic and prelimbic prefrontal cortical inhibition and dopaminergic modulation of basolateral amygdala neuronsin vivo.The Journal of Neuroscience, 22(1), 324−337.
Rottenberg, J., Gross, J. J., & Gotlib, I. H. (2005). Emotion context insensitivity in major depressive disorder.Journal of Abnormal Psychology, 114(4), 627−639.
Yang, Z. H., Li, C. Q., & Huang, F. L. (2013). Melatonin impaired acquisition but not expression of contextual fear in rats.Neuroscience Letters, 552, 10−14.
Yuan, L., Peng, M., Liu, D. W., & Zhou, R. L. (2011). Cognitive appraisal influences negative emotion experience and physiological activity.Acta Psychologica Sinica, 43(8),898−906.
[原琳, 彭明, 刘丹玮, 周仁来. (2011). 认知评价对主观情绪感受和生理活动的作用.心理学报, 43(8), 898−906.]
猜你喜欢
磁共振成像(2022年6期)2022-07-30
数学小灵通(1-2年级)(2021年10期)2021-11-05
数学小灵通(1-2年级)(2020年12期)2021-01-14
音乐生活(2020年12期)2020-01-05
当代陕西(2019年12期)2019-07-12
特别健康(2018年9期)2018-09-26
读友·少年文学(清雅版)(2018年2期)2018-09-10
特别健康(2018年2期)2018-06-29
中国老年学杂志(2017年13期)2017-07-18
小学阅读指南·低年级版(2016年10期)2016-09-10
