
听障和听力正常人群空间主导性和空间参照框架的交互作用*
2016-02-01 08:51:58王爱君迟莹莹刘晓乐
心理学报 2016年2期
王爱君 沈 路 迟莹莹 刘晓乐 陈 骐 张 明
(1苏州大学心理学系, 苏州 215123) (2东北师范大学心理学院, 长春 130024)
(3华南师范大学心理学院, 广州 510631)
1 引言
视觉信息的皮层加工至少存在两条分离性的神经通路:一条是从初级视觉皮层出发沿着背侧方向到达顶叶的“枕顶通路”, 即背侧通路; 另一条也是从初级视觉皮层出发沿着腹侧方向到达颞叶的“枕颞通路”, 即腹侧通路(Ungerleider & Mishkin,1982)。腹侧通路主要负责对客体属性的知觉, 将视觉输入转换为知觉表征, 包括客体的固有特征以及它们的空间关系。而背侧通路主要负责对客体的空间位置进行加工以及对客体施以行为动作, 如够取和抓握(Griffiths, Marslen-Wilson, Stamatakis, & Tyler,2013; Kim, 2014; Vossel, Weidner, Driver, Friston, &Fink, 2012)。
知觉/反应模型的观点认为, 对手臂范围内(近处空间)和手臂范围外(远处空间)的注意是基于不同的大脑信息加工通路(Berti & Frassinetti, 2000;Christiansen, Christensen, Grünbaum, & Kyllingsbæk,2014; Gilet, Diard, & Bessiere, 2011; Haazebroek,van Dantzig, & Hommel, 2011; Mennemeier, Wertman,& Heilman, 1992; Vuilleumier, Valenza, Mayer,Reverdin, & Landis, 1998)。因为个体可以对近处空间的客体直接地施以行为动作, 因此, 背侧通路负责近处空间的加工。相反, 出现在远处空间的客体,个体无法直接地对其施以行为动作, 因此, 腹侧通路负责对出现在远处空间的客体进行知觉表征。知觉/反应模型的提出是基于对一个枕颞皮层中双侧腹侧通路损伤患者的研究。尽管该病人患有高强度的视觉失认证, 也就是患者既不能正确地识别视觉客体也不能正确地辨认视觉客体, 但是该患者能够正确地对同样的视觉客体施以行为动作(Goodale &Milner, 1992; Goodale, Milner, Jakobson, & Carey,1991)。然而, 有研究表明, 无论是知觉任务还是反应任务, 只要其出现在近处空间就由背侧通路负责加工, 而出现在远处空间就由腹侧通路负责加工(Weiss, Marshall, Zilles, & Fink, 2003)。这些结果表明了, 背侧通路和腹侧通路在近处和远处空间表现出的不同激活模式本质上并不是依赖于任务需求(如, 知觉或者反应) (Pitzalis, Di Russo, Spinelli, &Zoccolotti, 2001), 而是依赖于一个关键性的因素,即背侧通路与腹侧通路的功能分离是与选择了合适的空间参照框架相关(Bruno, 2001)。在大脑中,一个客体既可以以自我参照框架的形式呈现(如,相对于观察者的身体效应器), 也可以以环境参照框架的形式呈现(如, 相对于另一个客体或者与观察者无关的周围事物) (Jiang & Swallow, 2013; Viarouge,Hubbard, & Dehaene, 2014; Vogeley & Fink, 2003)。自我表征通常是由背侧通路将其转化为相应的感觉运动表征(Andersen & Buneo, 2002; Andersen,Snyder, Bradley, & Xing, 1997; Cohen & Andersen,2002; Ma, Hu, & Wilson, 2012), 而环境表征则通常是由腹侧通路将其转化为相应的知觉表征(James,Culham, Humphrey, Milner, & Goodale, 2003; James,Humphrey, Gati, Menon, & Goodale, 2002)。在以往神经心理学的研究中, 要求先前提到的视觉失认证患者在基于自我参照和环境参照的条件下同时完成知觉任务和运动任务(Schenk, 2006)。结果发现,视觉失认证患者在基于自我参照框架条件下两种任务的成绩并未受到影响。相反, 在基于环境参照框架条件下两种任务的成绩受到了影响。实验表明,影响视觉失认证患者完成任务的一个关键性的因素可能是空间参照框架的选取(自我参照vs.环境参照)而不是任务需求(知觉vs.反应)。
以往研究表明, 背侧通路和腹侧通路在解剖上的差异导致了两者分别主要负责近处空间和远处空间的加工以及环境参照框架和自我参照框架的加工, 且以往研究较多的是分别关注空间主导性(近处空间 vs.远处空间)和空间参照框架(自我参照vs.环境参照)。但关于空间主导性和空间参照框架这两个维度的交互作用则关注较少。近期一项研究采用听力正常群体为研究对象, 考察了空间主导性和空间参照框架之间交互作用的神经机制。结果发现, 当腹侧通路负责的知觉表征与背侧通路负责的感觉运动表征存在交互作用时, 顶枕联合区(parietal-occipital junction, POJ)产生了更高的激活。研究认为, 顶枕联合区参与了背侧通路与腹侧通路交互作用的神经表征(Chen, Weidner, Marshall, &Fink, 2012)。这个问题之所以关键是因为, 其能够有效地处理三维环境中近处空间和远处空间的客体信息与日常生活的空间判断能力(不同参照框架)相结合的任务。来自神经心理学的研究也表明, 空间主导性可能会不同程度地影响着自我参照判断或者环境参照判断。如, 单侧视觉空间忽视症患者(即自我参照框架受损的患者)在近处空间或者远处空间完成二分任务时可以选择性地影响其基于环境参照的表征(Berti & Frassinetti, 2000; Berti, Smania,& Allport, 2001; Vuilleumier et al., 1998)。
本研究选取听障被试为主要研究对象, 将空间主导性与空间参照框架相结合, 目的在于考察听障人群中这两种维度之间潜在的交互作用。此外, 我们选用听力正常人群作为实验被试, 将听障人群与听力正常人群两组被试的成绩进行比较, 有利于考察空间主导性与空间参照框架的交互作用是否在听力丧失之后会发生改变。以往研究发现, 当要求听障人群完成视知觉任务时, 先天听障被试对运动刺激在外周视野的活动得到了增强。如, 当目标的时间或空间属性不确定时, 相对于听力正常被试,听障被试对外周视野呈现的运动刺激反应更快且更准确(Stevens & Neville, 2006)。但是对于中央视野呈现的刺激, 听力正常被试与听障被试的表现不存在显著差异(Bavelier, Dye, & Hauser, 2006)。此外,听障被试背侧通路的视觉运动区(middle temporal/medial superior temporal, MT/MST)对于外周视野出现的刺激表现出更高的激活, 而听力正常被试背侧通路的视觉运动区对中央视野出现的刺激有着更高的激活(Bavelier et al., 2001)。神经层面的证据也表明, 视觉运动刺激的加工(Beauchamp, Cox, &DeYoe, 1997; O’Craven, Rosen, Kwong, Treisman, &Savoy, 1997)和外周视野刺激的加工(Clavagnier, Prado,Kennedy, & Perenin, 2007; Prado et al., 2005)均由背侧通路负责。因此, 听障人群背侧通路的视觉功能更易受影响而具有可塑性的变化(Bavelier et al.,2001; Lomber, Meredith, & Kral, 2010)。此外, 有研究认为, 先天性的耳聋会引起前庭蜗神经的部分损伤, 不仅包括耳蜗功能, 还包括前庭传入。而前庭传入纤维的破坏是引起平衡缺陷的原因之一(Siegel,Marchetti, & Tecklin, 1991), 所以, 听障人群的本体平衡感会表现出比正常人差(Angelaki, Klier, &Snyder, 2009)
综上所述, 听障被试是因为背侧通路改变可能导致自我参照框架的变化, 进而导致空间主导性和空间参照框架的交互作用模式发生了变化, 还是因为其本体平衡感本身存在缺陷导致自我参照框架的变化, 进而导致空间主导性和空间参照框架的交互作用模式发生了变化?为了验证上述观点, 我们进行了本研究。实验1操纵了近处空间和远处空间客体自我参照下的位置和环境参照下的位置(图 1),要求听障和听力正常被试分别在近处空间和远处空间针对相同的物理刺激(实验中的叉子)做出自我参照框架和环境参照框架的判断。通过这样的实验设计, 主要考察空间主导性与空间参照框架之间潜在的交互作用是否在听力丧失之后会发生改变。但由于听障被试前庭蜗神经的部分损伤, 会导致听障被试的主观平衡感可能受到了影响(Siegel et al.,1991), 为了排除听障被试和听力正常被试在以自我参照判断任务中的差异是由基本的本体平衡能力上的差异造成的。实验2采用open loop实验考察听障和听力正常被试的空间判断能力差异是否是由于主观平衡感的差异所引起。如果有差异, 则实验1的结果必须考虑这种差异对实验结果造成的影响。如果没有差异, 则可以推断实验的效应主要是由于听障被试背侧通路发生了改变所引起的。
2 实验 1:听障和听力正常人群空间主导性和空间参照框架的交互作用
2.1 方法
2.1.1 被试
听障被试为 17名来自长春市特殊教育学校的学生(男生 7名, 女生 10名, 平均年龄为 17.5岁),听力丧失的年龄均为3周岁之前, 双耳听力损失大于 90 dB, 他们均熟练掌握了手语, 并且出生后就将其作为其第一语言。听力正常被试为 17名来自东北师范大学大学一年级学生(男生8名, 女生9名,平均年龄为 18.5岁)。所有被试均为右利手, 且视力或者矫正视力正常。实验所选取的听障被试为特殊师范教育和职业教育的学生, 与所选取的听力正常被试年龄接近, 且本实验的任务为空间参照框架判断, 所以两组被试受教育水平上的略微差异并不是一个影响实验结果的主要因素。所有被试在实验之前具有知情同意权, 且在完成实验后给予相应的报酬或礼品。
2.1.2 实验仪器和刺激
整个实验在一个微暗的实验室内进行, 听障被试在完成实验过程中由主试和手语老师共同对其进行实验的指导。远处空间的刺激通过笔记本电脑投影到大屏幕上呈现, 被试眼镜距离屏幕中央的距离为226 cm。近处空间的刺激通过14英寸IBM笔记本电脑呈现, 被试眼镜距离屏幕中央的距离为50 cm。远处空间刺激与近处空间刺激的视角保持一致。实验的刺激材料为一个盘子和盘子上的一个叉子。盘子的直径为 15°视角, 叉子近端尾部的视角为2.5°。以自我为参照条件下(相对于身体的中矢面)叉子的位置和以环境为参照条件下(相对于盘子的中矢面)叉子的位置成正交变化。叉子相对于被试身体的中矢面有不同的4种位置, 形成了4种以自我参照条件下叉子的位置:‒5°、‒3.5°、3.5°和 5°。叉子相对于盘子的中矢面有4种不同的位置, 形成了 4种以环境为参照条件下叉子的位置:‒2.4°、‒1.7°、1.7°和 2.4° (如图 1)。

图1 实验刺激材料(引自Zhang et al., 2014)
2.1.3 实验设计和实验程序
实验为2 (空间主导:近处空间vs.远处空间) ×2 (任务类型:自我参照vs.环境参照)的被试内设计。听障被试与听力正常被试分成两组, 分别要求其针对同样的刺激在近处空间和远处空间完成空间参照框架判断任务。在做自我参照判断的任务中, 要求被试判断叉子位于自己身体中线的左侧还是右侧。在做环境参照判断的任务中, 要求被试判断叉子位于盘子的左侧还是右侧。无论在近处空间还是远处空间做参照框架判断任务, 都要求一半的被试针对左侧的刺激用右手的食指按鼠标左键, 而对右侧的刺激用右手的中指按鼠标的右键。
实验中, 两种任务类型以组块的方式呈现给被试, 即自我参照判断和环境参照判断组块交替呈现。本研究中的实验并未采用中央注视点去引导被试的注意, 因为实验中若出现“+”, 被试在完成自我参照判断的任务时更加倾向将注视点作为参照物来判断叉子的位置。实验中, 每个组块呈现之前,以指导语的方式(3 s)告诉被试即将完成的任务类型。组块中每个试次呈现的时间共1650 ms (其中,目标刺激呈现时间为 150 ms, 被试反应时间为1500 ms), 10个试次为一组, 两种任务类型在近处空间和远处空间各 8组, 即每个被试需要完成320个试次。正式实验为11 min左右。实验中听障被试在进行正式实验之前需要进行5 min左右的练习来熟悉实验任务, 以确保实验的正确率。听力正常被试在进行正式实验之前需要进行3 min左右的练习来熟悉实验任务。
2.2 结果与分析
2.2.1 听障被试
剔除反应错误的试次; 剔除反应时小于 100 ms和大于1500 ms的试次; 剔除正负3个标准差以外的试次; 总剔除的试次占总试次的18.6%。表1显示了各条件下被试的平均反应时、错误率。将错误率进行2 (空间主导:近处空间vs.远处空间) × 2 (任务类型:自我参照vs.环境参照)被试内的重复测量方差分析。结果表明, 空间主导性和任务类型的主效应显著均不显著,Fs < 1。两者的交互作用也不显著,F(1,16) = 2.40,p> 0.05。

表1 听障被试各条件下的平均反应时(ms)和错误率(%)
将正确试次下的反应时进行 2 (空间主导:近处空间vs.远处空间) × 2 (任务类型:自我参照vs.环境参照)被试内的重复测量方差分析。结果表明,空间主导性的主效应显著,F(1,16) = 9.22,p< 0.01,η2= 0.36, 被试对远处空间条件下目标的反应(547 ms)显著慢于近处空间条件下目标的反应(527 ms)。任务类型的主效应显著,F(1,16) = 9.44,p< 0.01, η2=0.37, 被试对环境参照条件下目标的反应(516 ms)显著快于自我参照条件下目标的反应(559 ms)。此外, 空间主导性与任务类型的交互作用显著,F(1,16) = 4.33,p =0.05, η2= 0.21 (图 2)。进一步进行简单效应分析, 当目标出现在远空间时, 被试对环境参照条件下目标的反应(530 ms)显著快于自我参照条件目标的反应(564 ms),t(16) = 2.13,p< 0.05,d= 0.75; 当目标出现在近空间时, 被试对环境参照条件下目标的反应(501 ms)显著快于自我参照条件目标的反应(553 ms),t(16) = 3.93,p =0.001,d=1.38。
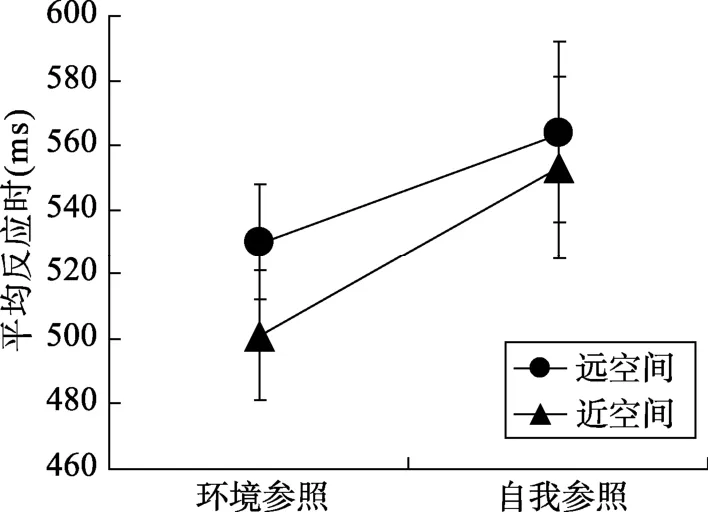
图2 听障被试各条件下的平均反应时和标准误(ms)
2.2.2 听力正常被试
剔除反应错误的试次; 剔除反应时小于 100 ms和大于1500 ms的试次; 剔除正负3个标准差以外的试次; 总剔除的试次占总试次的8.4%。表2显示了各条件下被试的平均反应时、错误率。将错误率进行2 (空间主导:近处空间vs.远处空间) × 2 (任务类型:自我参照vs.环境参照)被试内的重复测量方差分析。结果表明, 仅任务类型的主效应显著,F(1,17) = 8.21,p< 0.05, η2= 0.34, 环境参照条件下错误率(5%)显著少于自我参照条件下错误率(7.5%)。其他主效应和交互作用均不显著,Fs <1。
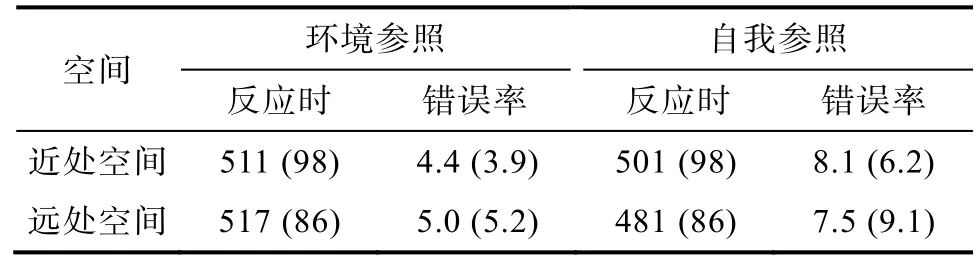
表2 听力正常被试各条件下的平均反应时(ms)和错误率(%)
将正确试次下的反应时进行 2 (空间主导:近处空间vs.远处空间) × 2 (任务类型:自我参照vs.环境参照)被试内的重复测量方差分析。结果表明,空间主导性的主效应不显著,F<1。任务类型的主效应显著,F(1,16) = 13.31,p< 0.005, η2= 0.45, 被试对环境参照条件下目标的反应(514 ms)显著慢于自我参照条件目标的反应(491 ms)。此外, 空间主导性与任务类型的交互作用显著,F(1,16) = 5.40,p<0.05, η2= 0.25 (图 3)。进一步进行简单效应分析, 当目标出现在远处空间时, 被试对环境参照条件下目标的反应(517 ms)显著慢于自我参照条件目标的反应(481 ms),t(16) = 3.64,p< 0.005,d= 0.64; 当目标出现在近处空间时, 被试对环境参照条件下目标的反应(511 ms)与自我参照条件目标的反应(501 ms)无显著差异,t(16) = 1.55,p >0.05。
通过听障被试和听力正常被试的数据可以看出, 听障被试的结果表现为环境参照框架下的反应(516 ms)快于自我参照框架下的反应(559 ms), 但是听力正常被试却表现出相反的模式, 即环境参照框架下的反应(514 ms)慢于自我参照框架下的反应(491 ms)。而且, 将两组被试的结果进行对比可以看出, 听障(516 ms)和听力正常(514 ms)被试在环境参照框架条件下的反应时之间并无显著差异,t<1。但自我参照框架条件下的反应时之间存在显著差异,t(16) = 2.21,p =0.05,d= 0.78, 表现为, 听障被试在自我参照条件下的反应(559 ms)慢于听力正常被试自我参照框架下的反应(491 ms)。
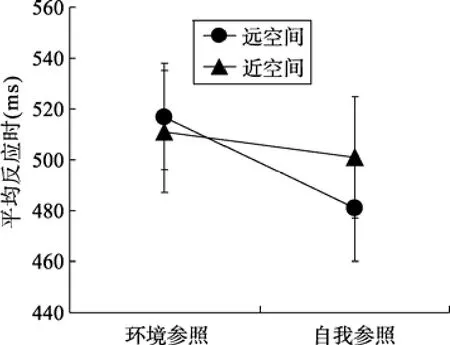
图3 听力正常被试各条件下的平均反应时和标准误(ms)
同时我们也可通过两组被试的数据看出, 听障和听力正常被试的空间主导性和空间参照框架具有不同的交互作用模式。听障被试在近空间与远空间做环境参照框架判断的反应时存在差异, 表现为远空间条件下的反应时长于近空间条件下的反应时。但是, 听障人群在近空间与远空间做自我参照框架判断的反应时不存在差异。而听力正常被试却表现出相反的模式, 近处空间与远处空间做环境参照框架判断的反应时不存在差异, 但在近处空间与远处空间做自我参照框架判断的反应时存在差异,表现为远处空间条件下的反应时短于近处空间条件下的反应时。
3 实验 2:听障和听力正常人群本体平衡感的差异
为了排除实验效应的出现是由于听障人群的本体平衡感比正常人差, 进而表现出对以自我参照框架下位置的感觉能力存在缺陷。我们采用 open loop实验来考察听障和听力正常被试的空间判断能力差异是否由于主观平衡感的差异所引起。有研究在猴子的背侧通路中与够取和抓握反应相关的区域发现了编码手的位置反馈信息的神经元(Galletti,Kutz, Gamberini, Breveglieri, & Fattori, 2003; Galletti,Gamberini, Kutz, Baldinotti, & Fattori, 2005; Pitzalis et al., 2013)。因此, 在够取和抓握行为中, 对手的位置进行实时的视觉加工可能依赖于背侧通路。采用fMRI技术通过比较close loop条件下(伸手时能看到自己的手)和 open loop条件下(看不到自己手的位置)的够取和抓握行为, 可以考察人类是否也存在类似的够取和抓握区域来调节对手的视觉反馈, 并且发现, 视觉运动区(middle temporal cortex,MT)和上枕皮层(superior occipital cortex, SO)参与对手的运动的视觉反馈, 而这些区域都位于背侧通路(Thaler & Goodale, 2011)。实验过程中确保被试不能看到自己的手(实验装置如图 4), 由于参与对手的运动视觉反馈的脑区都位于背侧通路, 实验中看不到手的运动就实现不了对手运动的视觉反馈。此外, 背侧通路既是视觉反馈的通路, 也是触觉/本体感觉反馈的通路(Milner, 2012; Whitwell &Buckingham, 2013)。实验过程中被试即使看不到视觉客体, 只要有触觉反馈, 也能够促进抓握行为(Bruno& Franz, 2009; Franz, Hesse, & Kollath, 2009)。但在本实验中并未给予被试包含位置信息的触觉反馈,在这过程中被试也不能根据触碰的位置调整判断,因为被试在完成“点”追随任务后也不知道是否准确。因此, 是没有所谓触觉反馈的位置信息对任务判断造成的影响。所以, 即使听障和听力正常人群的触觉反馈存在差异, 但在本任务中, 触觉并未对任务判断产生作用。所以, 通过open loop实验的设计和装置使得被试在完成实验任务时既没有视觉反馈也没有触觉反馈(一定程度上限制了背侧通路的使用), 而完全依赖于本体的主观感觉。
3.1 方法
3.1.1 被试
实验1所有被试均参与本实验。
3.1.2 实验仪器和设备
实验刺激为出现在屏幕不同位置上的红色小方框(3 mm × 3 mm)。刺激通过一个22吋的LCD显示器垂直投射在一面镜子上(550 mm × 350 mm)。该镜子安装在一个 600 mm (长) × 400 mm (宽) ×300 mm (高)的去除了底面的纸箱子上, 箱子则置于700 mm高的桌子上。LCD显示器到镜子的距离与镜子到桌面的距离相等, 均为 300 mm, 这样的设置可以使得从镜子里看到的刺激到镜子的距离与镜子到桌面的距离一致, 因此, 让被试可以感觉目标刺激好似呈现在桌面上(图 4)。目标刺激可能出现在屏幕中矢面的左侧或者右侧, 每次都有 10种不同的位置, 距离屏幕中矢线的距离分别为23 mm、37 mm、51 mm、65 mm、79 mm、93 mm、106 mm、120 mm、134 mm和148 mm。每种位置在左右两侧均重复5次, 共计目标出现在左侧50次, 出现在右侧50次。被试用手指出目标的位置通过一张铺在桌面上的白纸(550 mm × 350 mm)记录, 整个手指追随目标的过程通过外置摄像头进行实时记录(图4)。
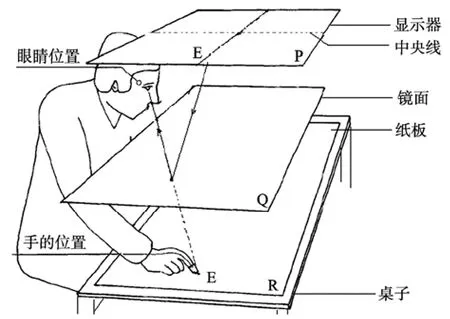
图4 Open loop实验设备。目标点呈现在平面P上, 沿着图中的实线方向呈现, 并被投射到镜子(Q平面)中。被试直立坐在桌子前, 保持头部固定, 眼睛看向镜子中的目标, 确保不能看到自己的手, 手指在白纸上(平面R)指出镜中点的位置。一个摄像头位于被试的左侧以记录被试的整个反应过程(引用并修改自Prablanc, Echallier, Komilis, & Jeannerod,1979)。
3.1.3 实验设计
实验过程中要求被试正直地坐在桌子前, 身体中线对着镜子中的中线, 眼睛看向镜子里的目标,并确保被试不能看到自己的手。将被试用来指出目标位置的手指绑上棉签, 其中左侧目标对应左手,蘸上黑墨水。右侧目标对应右手, 蘸上红墨水。一半被试先用左手追随左侧目标, 后用右手追随右侧目标, 另一半被试则反过来, 达到被试间平衡。实验中要求被试尽可能准确地指出目标的位置, 确定目标的位置后就用手上的棉签点在纸上相应的位置, 时间上不做限制。因此, 手的移动主要是依赖于目标相对于被试以自我参照进行判断的位置上。
实验为2 (被试类型:听障人群vs.听力正常人群) × 2 (反应手:左手vs.右手)的混合设计。因变量为屏幕中目标点距离镜子中线的实际距离与记录在纸上的相应点到中线的距离之差的绝对值。
3.2 结果与分析
将手指追踪的目标位置与实际位置的偏差进行2 (被试类型:听障人群vs.听力正常人群) × 2 (反应手:左手vs.右手)的重复测量方差分析。结果发现, 只有反应手的主效应显著,F(1,32) = 7.07,p<0.05, η2= 0.23。表明了左手的偏差(37 mm)大于右手的偏差(28 mm)。被试类型的主效应和交互作用均不显著,F值均小于1 (如图5)。
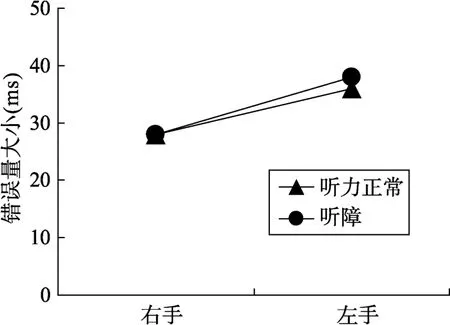
图5 open-loop实验各条件下手追踪的目标位置与目标实际位置的偏差
实验可以看出被试类型的差异并不显著, 而只存在左右手间的显著差异。由于实验中所有被试均为右利手, 因而, 无论听障被试还是听力正常被试,右手的表现都比左手好。尽管在open loop任务中要判断的位置也是以自我参照判断下的坐标, 但由于没有身体运动的视觉反馈, 实验中也没有提供位置信息的触觉反馈, 因而在完成“点”追随反应的任务中被试主要依靠自身的本体平衡感。实验表明了,听障被试和听力正常被试在基本的本体平衡能力上没有差异, 因而可以证实, 两类被试在以自我参照判断任务中的差异可以排除这个影响, 即听障被试是因为背侧通路改变而导致了自我参照框架的变化, 进而导致空间主导性和空间参照框架的交互作用模式发生了变化。
4 讨论
本研究目的在于考察听障人群空间主导性和空间参照框架之间潜在的交互作用, 并且通过将听障人群与听力正常人群两组被试进行比较, 考察空间主导性和空间参照框架之间的交互作用是否在听力丧失之后会发生改变。实验1要求听障和听力正常被试分别在近处空间和远处空间内完成不同空间参照框架的判断。结果发现, 对于听障被试来说, 空间主导性和空间参照框架之间存在交互作用。无论目标出现在远处空间还是近处空间, 被试对环境参照条件下目标的反应均显著快于自我参照条件目标的反应。但在近处空间条件下, 两者的差异更大一些,t(16) = 2.20,p< 0.05,d= 0.77 (图2)。对于听力正常被试来说, 虽然空间主导性和空间参照框架之间也存在交互作用, 但与听障被试的模式不同。当目标出现在远处空间时, 被试对环境参照条件下目标的反应显著慢于自我参照条件目标的反应; 当目标出现在近处空间时, 被试对环境参照条件下目标的反应与自我参照条件目标的反应无差异(图 3)。实验 2要求听障和听力正常被试分别完成open loop实验中。结果发现, 两类被试在本体平衡感上并不存在显著差异, 从而证明了并不是由于听障被试在主体平衡感上存在缺陷对实验效应造成的影响, 而是由于听障被试因背侧通路发生了改变而导致其在完成自我参照框架判断时产生的变化。
我们的实验结果发现, 听力正常被试在环境参照判断条件下的反应时短于自我参照框架条件, 这与以往研究的结果较为一致(Chen et al., 2012)。通过听障和听力正常被试进行对比可以看出, 听障被试在完成环境参照判断中的表现与听力正常被试的表现相当, 而在完成自我参照框架判断的反应时长于听力正常被试。因此可以说明, 听障被试空间参照框架中的环境参照框架判断的能力正常, 而自我参照框架判断的能力受到的一定的损害, 而自我参照框架判断是由背侧通路负责的(Andersen &Buneo, 2002; Cohen & Andersen, 2002; Andersen et al.,1997), 因此, 可以说听障被试自我判断能力的受损是由背侧通路的变化所引起的, 而这又导致了听障被试在空间主导性和空间参照框架之间的交互模式与听力正常被试不同。
听障被试在不同参照框架的表现不同是他们自身长期生活过程中逐渐累积的。由于自我参照框架的表征是依赖于身体效应器(指向行为目标的部位)的坐标信息。而环境参照框架的表征则依赖于外部世界客体位置的坐标信息。比如, 在日常生活中, 当我们走在马路上的时候, 身后不远的地方一辆呼啸而来的汽车正向我们驶来, 这时我们需要将视线立刻转移到这辆车上。在这过程中了解车辆鸣笛的空间方位信息, 对于我们判断我们与车辆之间的位置起到很重要的作用。因此, 自我参照框架的判断中, 来自其他感觉通道的信息也起到很重要的作用, 如声音。而听障人群由于长期缺少听觉信息的传入, 进而会影响他们对自我参照框架的表征,导致他们在对自我参照框架的判断不如听力正常的被试。此外, 由于听障被试长久以来使用手语进行日常的沟通和交流, 为了更好的与人沟通和交流, 他们必须注意到对方手的姿势、位置以及两只手的位置关系与身体之间的位置关系(Heracleous, Beautemps,& Aboutabit, 2010), 而这些讯息均属于环境参照框架的范畴。因此, 听障被试在环境参照框架的判断表现完好, 自我参照框架的判断较差。
有研究认为, 听障人群的本体感觉存在着缺陷,而本体平衡感对于身体倾斜的感知以及空间朝向则较为重要(Angelaki et al., 2009)。因此, 这会导致听障被试对相对于自身空间位置的判断存在一定程度的影响。在我们的实验中为了控制这一问题造成的影响, 通过 open loop实验来对此问题进行说明。实验结果说明了, 听障被试和听力正常被试并不存在着这一问题造成的差异, 因此排除了听障被试自我参照框架表征的损伤是由本体感觉的缺失所引起。
远处空间刺激的加工依赖于腹侧通路, 腹侧通路同样也参与空间参照框架中环境参照框架的表征。而近处空间刺激的加工依赖于背侧通路, 背侧通路同样也参与空间参照框架中自我参照框架的表征。由于听障被试背侧通路受损, 而腹侧通路完好, 因此, 他们的结果表现为, 无论是在近处空间还是在远处空间进行自我参照框架的判断都表现出较长的反应时, 且两者并不存在显著差异。而在进行环境参照框架的判断时, 远处空间条件下的反应时显著长于近处空间条件, 这一结果与我们的假设有些不符合。我们认为, 远处空间的表征和环境参照的表征均由腹侧通路负责, 因此, 远处空间条件下环境参照的表征较易得到表征, 而近处空间条件下环境参照的表征存在背侧与腹侧通路之间的冲突, 因此, 这种条件下大脑中需要有一个将来自背侧通路的信息与来自腹侧通路的信息进行交换的场所, 根据以往研究发现, 顶枕联合区(Parietal-Occipital Junction, POJ)则在这过程中起到了这样的关键作用(Chen et al., 2012)。并且, 在进行环境参照框架的判断时, 相对于远处空间刺激的表征, 顶枕联合区更偏向于对近处空间刺激进行表征(Chen et al., 2012; Quinlan & Culham, 2007; Weiss et al.,2003)。因此, 听障被试的结果表现出, 即使在进行环境参照框架的判断, 近处空间条件下的反应时仍然短于远处空间。听力正常被试的结果正好相反,无论在近处空间还是在远处空间进行环境参照框架判断的反应时均较长, 且两者之间并不存在显著差异。而在进行自我参照框架的判断时, 近处空间条件下的反应时显著长于远处空间条件。相对于近处空间刺激的表征, 顶枕联合区更偏向于对远处空间刺激进行表征(Chen et al., 2012; Quinlan &Culham, 2007; Weiss et al., 2003)。因此, 听力正常被试的结果表现出, 即使在进行自我参照框架的判断, 远处空间条件下的反应时仍然短于近处空间。
5 结论
(1)听障被试由于背侧通路发生了改变, 进而导致了自我参照框架判断的能力受损;
(2)听障被试与听力正常被试在空间主导性和空间参照框架中的交互作用模式不同。
致谢:感谢长春市特殊教育学校段老师和王老师对实验顺利开展提供的帮助与支持。
Andersen, R. A., & Buneo, C. A. (2002). Intentional maps in posterior parietal cortex.Annual Review of Neuroscience,25, 189–220.
Andersen, R. A., Snyder, L. H., Bradley, D. C., & Xing, J.(1997). Multimodal representation of space in the posterior parietal cortex and its use in planning movements.Annual Review of Neuroscience, 20, 303–330.
Angelaki, D. E., Klier, E. M., & Snyder, L. H. (2009). A vestibular sensation: Probabilistic approaches to spatial perception.Neuron, 64(4), 448–461.
Bavelier, D., Brozinsky, C., Tomann, A., Mitchell, T., Neville,H., & Liu, G. (2001). Impact of early deafness and early exposure to sign language on the cerebral organization for motion processing.Journal of Neuroscience, 21, 8931–8942.
Bavelier, D., Dye, M. W. G., & Hauser, P. C. (2006). Do deaf individuals see better?.Trends in Cognitive Sciences, 10,512–518.
Beauchamp, M. S., Cox, R. W., & DeYoe, E. A. (1997).Graded effects of spatial and featural attention on human area MT and associated motion processing areas.Journal of Neurophysiology, 78, 516–520.
Berti, A., & Frassinetti, F. (2000). When far becomes near:Remapping of space by tool use.Journal of Cognitive Neuroscience, 12(3), 415–420.
Berti, A., Smania, N., & Allport, A. (2001). Coding of far and near space in neglect patients.NeuroImage, 14, S98–S102.
Bruno, N. (2001). When does action resist visual illusions?.Trends in Cognitive Sciences, 5, 379–382.
Bruno, N., & Franz, V. H. (2009). When is grasping affected by the Müllerr-Lyer illusion? A quantitative review.Neuropsychologia, 47, 1421–1433.
Chen, Q., Weidner, R., Weiss, P. H., Marshall, J. C., & Fink, G.R. (2012). Neural interaction between spatial domain and spatial reference frame in parietal-occipital junction.Journal of Cognitive Neuroscience, 24(11), 2223–2236.
Christiansen, J. H., Christensen, J., Grünbaum, T., & Kyllingsbæk,S. (2014). A common representation of spatial features drives action and perception: Grasping and judging object features within trials.PLoS One, 9(5), e94744.
Clavagnier, S., Prado, J., Kennedy, H., & Perenin, M. T.(2007). How humans reach: Distinct cortical systems for central and peripheral vision.Neuroscientist, 13, 22–27.
Cohen, Y. E., & Andersen, R. A. (2002). A common reference frame for movement plans in the posterior parietal cortex.Nature Reviews Neuroscience, 3, 553–562.
Franz, V. H., Hesse, C., & Kollath, S. (2009). Visual illusions,delayed grasping, and memory: No shift from dorsal to ventral control.Neuropsychologia, 47, 1518–1531.
Galletti, C., Gamberini, M., Kutz, D. F., Baldinotti, I., &Fattori, P. (2005). The relationship between V6 and PO in macaque extrastriate cortex.European Journal of Neuroscience, 21, 959–970.
Galletti, C., Kutz, D. F., Gamberini, M., Breveglieri, R., &Fattori, P. (2003). Role of the medial parieto-occipital cortex in the control of reaching and grasping movements.Experimental Brain Research, 153, 158–170.
Gilet, E., Diard, J., & Bessiere, P. (2011). Bayesian action–perception computational model: Interaction of production and recognition of cursive letters.PLoS One,6(6), e20387.
Goodale, M. A., & Milner, A. D. (1992). Separate visual pathways for perception and action.Trends in Neurosciences, 15(1),20–25.
Goodale, M. A., Milner, A. D., Jakobson, L. S., & Carey, D. P.(1991). A neurological dissociation between perceiving objects and grasping them.Nature, 349, 154–156.
Griffiths, J. D., Marslen-Wilson, W. D., Stamatakis, E. A., &Tyler, L. K. (2013). Functional organization of the neural language system: Dorsal and ventral pathways are critical for syntax.Cerebral Cortex, 23(1), 139–147.
Haazebroek, P., van Dantzig, S., & Hommel, B. (2011). A computational model of perception and action for cognitive robotics.Cognitive Processing, 12, 355–365.
Heracleous, P., Beautemps, D., & Aboutabit, N. (2010). Cued Speech automatic recognition in normal-hearing and deaf subjects.Speech Communication, 52, 504–512.
James, T. W., Culham, J., Humphrey, G. K., Milner, A. D., &Goodale, M. A. (2003). Ventral occipital lesions impair object recognition but not object-directed grasping: An fMRI study.Brain, 126, 2463–2475.
James, T. W., Humphrey, G. K., Gati, J. S., Menon, R. S., &Goodale, M. A. (2002). Differential effects of viewpoint on object-driven activation in dorsal and ventral streams.Neuron, 35, 793–801.
Jiang, Y. V., & Swallow, K. M. (2013). Spatial reference frame of incidentally learned attention.Cognition, 126(3), 378–390.
Kim, H. (2014). Involvement of the dorsal and ventral attention networks in oddball stimulus processing: A meta-analysis.Human Brain Mapping, 35(5), 2265–2284.
Lomber, S. G., Meredith, M. A., & Kral, A. (2010). Cross-modal plasticity in specific auditory cortices underlies visual compensations in the deaf.Nature Neuroscience, 13,1421–1427.
Ma, Y. Y., Hu, X. T., & Wilson, F. A. (2012). The egocentric spatial reference frame used in dorsal–lateral prefrontal working memory in primates.Neuroscience & Biobehavioral Reviews, 36(1), 26–33.
Mennemeier, M., Wertman, E., & Heilman, K. M. (1992). Neglect of near peripersonal space: Evidence for multidirectional attentional systems in humans.Brain, 115(1), 37–50.
Milner, A. D. (2012). Is visual processing in the dorsal stream accessible to consciousness?.Proceedings of the Royal Society, 279, 2289–2298.
O’Craven, K. M., Rosen, B. R., Kwong, K. K., Treisman, A.,& Savoy, R. L. (1997). Voluntary attention modulates fMRI activity in human MT-MST.Neuron, 18, 591–598.
Pitzalis, S., Di Russo, F., Spinelli, D., & Zoccolotti, P. (2001).Influence of the radial and vertical dimensions on lateral neglect.Experimental Brain Research, 136, 281–294.
Pitzalis, S., Sereno, M. I., Committeri, G., Fattori, P., Galati, G.,Tosoni, A., & Galletti, C. (2013). The human homologue of macaque area V6A.NeuroImage, 82, 517–530.
Prablanc, C., Echallier, J. F., Komilis, E. K., & Jeannerod, M.(1979). Optimal response of eye and hand motor systems in pointing at a visual target.Biological Cybernetics, 35,113–124.
Prado, J., Clavagnier, S., Otzenberger, H., Scheiber, C., Kennedy,H., & Perenin, M. T. (2005). Two cortical systems for reaching in central and peripheral vision.Neuron, 48,849–858
Quinlan, D. J., & Culham, J. C. (2007). fMRI reveals a preference for near viewing in the human parieto-occipital cortex.NeuroImage, 36,167–187.
Schenk, T. (2006). An allocentric rather than perceptual deficit in patient D.F.Nature Neuroscience, 9, 1369–1370.
Siegel, J. C., Marchetti, M., & Tecklin, J. S. (1991).Age-related balance changes in hearing-impaired children.Physical Therapy, 71, 183–189.
Stevens, C., & Neville, H. (2006). Neuroplasticity as a doubleedged sword: Deaf enhancements and dyslexic deficits in motion processing.Journal of Cognitive Neuroscience, 18,701–714.
Thaler, L., & Goodale, M. A. (2011). Neural substrates of visual spatial coding and visual feedback control for hand movements in allocentric and target-directed tasks.Frontiersin Human Neuroscience, 5, 92.
Ungerleider, L. G., & Mishkin, M. (1982). Two cortical visual systems. In : D. J. Ingle, M. A. Goodale, & R. J. W.Mansfield (Eds.),Analysis of visual behavior(pp. 549–586).Cambridge, MA: MIT Press.
Viarouge, A., Hubbard, E. M., & Dehaene, S. (2014). The organization of spatial reference frames involved in the SNARC effect.The Quarterly Journal of Experimental Psychology, 67(8), 1484–1499.
Vogeley, K., & Fink, G. R. (2003). Neural correlates of the first-person-perspective.Trends in Cognitive Sciences, 7(1),38–42.
Vossel, S., Weidner, R., Driver, J., Friston, K. J., & Fink, G. R.(2012). Deconstructing the architecture of dorsal and ventral attention systems with dynamic causal modeling.Journal of Neuroscience, 32(31), 10637–10648.
Vuilleumier, P., Valenza, N., Mayer, E., Reverdin, A., &Landis, T. (1998). Near and far visual space in unilateral neglect.Annals of Neurology, 43, 406–410.
Weiss, P. H., Marshall, J. C., Zilles, K., & Fink, G. R. (2003).Are action and perception in near and far space additive or interactive factors?.NeuroImage, 18, 837–846.
Whitwell, R. L., & Buckingham, G. (2013). Reframing the action and perception dissociation in DF: Haptics matters,but how?.Journal of Neurophysiology, 109, 621–624.
Zhang, M., Tan, X. Y., Shen, L., Wang, A. J., Geng, S., & Chen,Q. (2014). Interaction between allocentric and egocentric reference frames in deaf and hearing populations.Neuropsychologia, 54, 68–76.
猜你喜欢
领导文萃(2023年3期)2023-02-15 01:18:16
小资CHIC!ELEGANCE(2022年1期)2022-01-11 00:49:59
数学物理学报(2020年3期)2020-07-27 01:19:46
大连民族大学学报(2020年2期)2020-06-16 03:13:00
时代英语·初中(2018年2期)2018-05-14 13:28:01
时代英语·初中(2018年2期)2018-05-14 13:28:01
时代英语·初中(2018年2期)2018-05-14 13:28:01
时代英语·初中(2018年2期)2018-05-14 13:28:01
法大研究生(2017年1期)2017-04-10 08:55:06
教学月刊小学版·语文(2016年10期)2017-03-02 18:55:50
