
外倾个体的自我参照加工程度效应*
2016-02-01 09:18:39钟毅平杨子鹿欧阳益蔡荣华李慧云傅小兰
心理学报 2016年8期
范 伟 钟毅平 杨子鹿 李 琎 欧阳益 蔡荣华 李慧云 傅小兰
(1湖南师范大学, 认知与人类行为湖南省重点实验室, 长沙 410082)
(2中国科学院心理研究所, 脑与认知科学国家重点实验室, 北京 100101)
1 引言
心理学家们普遍认为, 在潜意识下的自我, 它不仅仅对人类的情绪情感产生以及需要、动机等心理倾向性的激发起着重要作用, 而且还对人类的认知活动有着最直接的影响。Rogers等人最早提出了自我参照效应, 是指当信息与自我概念有关时, 个体会进行快速的加工和更好的记忆(Rogers, Kuiper,& Kirker, 1977)。此后, 越来越多的学者开始关注这一热点问题。有大量的学者考察了自我参照加工的神经机制及其影响因素。周爱保等(2013)发现相对于非自我相关物主代词“他的”, 自我相关物主代词“我的”引发了更大的theta频带(4~8 Hz)能量活动(周爱保等, 2013)。Brédart 等人采用被试的生日作为自我参照刺激, 结果发现当目标人物的生日月份与被试自己的生日同月份时, 被试回忆的正确率更高(Brédart, François, & Guimond, 2013)。钟毅平、范伟等发现对比集体自我参照刺激, 个体自我参照刺激虽然容易获得更为精细的加工, 但也容易受到情境的影响(钟毅平等, 2012)。Fan等人考察了国旗诱发的自我参照加工, 结果发现了对比熟悉刺激和陌生刺激, 自我参照刺激诱发了更大的P3 (Fan et al.,2011)。Zhao等人考察了4类名字刺激, 发现了集体自我存在的神经科学证据(Zhao et al., 2009)。回顾前人的文献, 我们发现, 上述研究大多是以自我相关和非自我刺激去激活被试行为和神经层面上的差异来考察自我相关效应, 但并没有考虑到这些刺激与自我的相关程度如何。在现实生活中, 刺激与自我的接近程度不同, 对于个体而言往往意义也会不一样, 对高自我相关刺激的加工比低自我相关刺激的加工具有更大的生理和社会意义。这就是为什么同样都是名字, 而且都能比无关自我刺激引起更大的注意, 但是听到自己的名字会比听到朋友的名字引起更大警觉性的原因。可见, 人脑的不同反应, 可能反映了对待不同自我相关程度的刺激, 脑的加工是存在差异的。
Fan等人在自我参照效应的基础上提出了自我参照加工的程度效应, 指与低自我相关的刺激相比,高自我相关的刺激会得到大脑更快和更精细的加工(Fan et al., 2013)。还有一些学者采用了面孔刺激作为自我相关程度不同的刺激来考察自我参照效应。如Ma和Han比较了被试自己的面孔和朋友的面孔, 结果发现对比朋友的面孔, 被试自己的面孔在右侧梭状回的激活更大(Ma & Han, 2012)。Keyes和他的同事比较了被试自己的面孔, 朋友的面孔以及陌生人的面孔, 结果发现对比其他面孔, 被试对于自己面孔的加工更快(Keyes, Brady, Reilly, & Foxe,2010)。Sui等人的研究和Caharel等人的研究也表明了相比其他熟悉面孔, 人脑对自己面孔的认知加工是一种自动的加工, 发生在面孔结构编码之后, 并且这种加工是独立进行的, 与任务无关(Caharel,Courtay, Bernard, Lalonde, & Rebai, 2005; Sui, Zhu, &Han, 2006)。此外, 还有一些学者采用名字作为实验材料, 直接或间接考察了自我参照加工的程度效应。例如, Chen 等人直接考察了不同相关程度刺激物的自我参照效应, 结果发现对比所在的省份、国家的名字和熟悉的名字, 被试自己的名字激发了更大的P3波幅(Chen et al., 2011)。Tacikowski等人考察了名字和面孔的自我参照效应, 研究结果发现对比不熟悉的名字、熟悉的名字以及自己的名字来说, 被试对自己的面孔的学习更容易(Tacikowski, Jednoróg,Marchewka, & Nowicka, 2011)。Tacikowski和同事还发现对比名人和陌生人的名字来说, 无论在听觉模式下, 还是视觉模式下, 个体自己的名字和重要他人的名字在内侧前额叶皮质(MPFC)上有更大的激活(Tacikowski et al., 2012)。Tateuchi等人也发现,相比陌生人的名字和父母的名字, 被试在早期的前注意加工阶段就能分辨出自己的名字(Tateuchi, Itoh, &Nakada, 2012)。Höller等人也发现, 相比他人的名字和反顺序的自己名字, 被试对正常顺序的自己名字所诱发的α-β波的活动更弱(Höller et al., 2011)。总之, 相比用自我相关和非自我刺激来考察被试自我相关效应的研究, 这些研究都直接或间接采用不同程度的自我相关刺激考察了自我参照加工的程度效应, 得到的结果更精细更可靠。
回顾前人的研究我们还发现, 正性情绪与自我参照加工的有着紧密的联系。例如, Watson和Dritschel等人运用自我参照范式, 在被试对情绪性字词进行自我相关判断时记录其脑电, 结果发现对比其他条件, 被试认为正性情绪词与自己的相关程度更高(Watson, Dritschel, Obonsawin, & Jentzsch, 2007)。陈芸、钟毅平、周海波、周路平和王小艳(2012)采用了阈下启动范式, 被试的任务是判断情绪词是否能够描述自己或他人, 结果发现被试认为正性情绪词更能够描述自己, 存在自我正面偏见(陈芸等,2012)。还有学者发现中间型个体的正性情绪与自我参照加工及程度效应虽然紧密相关, 但二者拥有着独立的加工系统, 互不干扰(钟毅平等, 2014)。对比中间型个体, 外倾个体的表现有很大不同。例如,有大量的研究发现, 人格特质中的外倾性与个体的正性情绪有着更高程度的相关(Amin, Todd Constable, &Canli, 2004; Costa & McCrae, 1990; Eysenck, 1990; 何媛媛, 袁加锦, 伍泽莲, 李红, 2008; 黄敏儿, 郭德俊,2004; 李中权, 王力, 张厚粲, 柳恒超, 2010)。何媛媛等人的研究发现外倾性与正性情绪存在高度相关, 主要表现为外倾个体在日常生活中体验到更多的正性情绪; 外倾个体对正性刺激有更为强烈的愉悦体验和情绪反应; 外倾个体对正性刺激有注意等认知加工偏向; 外倾程度与皮层及皮层下某些组织的激活程度相关(何媛媛等, 2008)。胡艳华等人的研究发现与高神经质比较, 高外倾确实更容易激活其正性情绪, 而且, 更容易对其负性情绪实施减弱调节(胡艳华, 黄敏儿, 2006)。娄熠雪等人发现外倾性人格是与情绪活动密切相关的人格特质(娄熠雪,蔡阿燕, 杨洁敏, 袁加锦, 2014)。以往的这些研究表明高外倾个体爱社交、与他人有更多沟通与交流、比较乐观, 他们对自身的情绪调节能力也有更积极的评价, 在面临情绪事件尤其是负性情绪事件时更有信心调节好自身的情绪, 能够尝试采取多种有效措施以降低外界事件带来的情绪冲击, 因而能够体验到更多的正情绪。所以, 对比中间型个体,外倾性人格的自我参照加工及程度效应可能会展现不一样的特性。
综上所述, 尽管目前没有研究表明外倾个体在社会适应性, 工作状况, 收入水平, 寿命等方面与其他个体是否存在差异, 但由于外倾个体对正性情绪刺激投入了更多的注意资源, 更容易受到环境中正性情绪的干扰, 从而会使其他认知加工, 包括自我参照加工受到影响。因此, 我们预期在对自我参照刺激进行加工时, 对比中间型个体, 外倾性人格特质个体的自我参照加工及程度效应会减弱。
2 方法
2.1 实验目的与实验假设
实验目的:本实验采用三类刺激的oddball实验模式, 以探讨外倾性人格个体自我参照加工程度效应的特性。
实验假设:外倾性人格被试中自我参照加工的程度效应是存在的。但与中间型人格被试相比, 外倾性人格被试诱发了更弱的自我参照加工程度效应。
2.2 被试
采用大五人格量表, 内外倾维度分量表, 从在校本科生中选出18名外倾维度得分最高的被试,即外倾个体(得分:32分至45分, 39 ± 4.12), 其中男生8名, 年龄在19~25岁之间, 平均为21.58岁。再选出20名中间型被试, 即控制组(得分:−5分至5分, 0.43 ± 3.87), 其中男生9名, 年龄在18~24岁之间, 平均为20.72岁。被试共计38人, 均为右利手, 身体健康无神经系统疾病, 没有脑部损伤史,视力正常或校正后正常。实验完成后付给一定报酬。
2.3 实验材料
采用三类刺激的oddball范式, 其中标准刺激是三字短语“红绿蓝”, 比例最大,占到所有trial的70%; 目标刺激是3个汉字“的地得”, 只占6%, 被试的任务是对目标刺激进行按键反应; 偏差刺激是4种自我相关的名字, 各占6%, 其中被试自己的名字与自我高相关, 被试父亲的名字与自我中度相关,被试所在国的国家领导人名字与自我为低度相关,熟悉但不具备归属感的外国国家领导人的名字作为熟悉刺激(例如:奥巴马)。实验后被试采用9点量表对四类偏差刺激的自我相关评定结果分别是8.82 ± 0.52, 7.14 ± 0.43, 5.63 ± 0.67和3.18 ± 0.72。
2.4 实验设计
实验为2人格类型(外倾性, 控制组)×4自我相关类型:(高自我相关、中等自我相关、低自我相关和熟悉刺激) 两因素混合设计, 因变量为ERP数据(P2和N2的波幅与潜伏期, P3的平均波幅)。
2.5 实验程序
被试戴好电极帽后舒适地坐在光线柔和的隔音室里, 双眼距电脑屏幕约为1 m, 水平和垂直视角均在5°以内。在本研究中, 标准刺激呈现560次(70%), 目标刺激呈现48次(6%), 而每类偏差刺激分别呈现48次(6%)。整个实验刺激分为4个区组(block), 每个区组呈现200个刺激序列(trials), 总共800个刺激序列(trials)。在每个刺激序列中, 首先在屏幕中央呈现一个300 ms的十字架, 紧接着随机呈现800~1200 ms的黑屏, 之后呈现500 ms的三类刺激之一, 最后再呈现1000 ms的黑屏。在呈现三类刺激的时, 被试的任务是认真看刺激但只对目标刺激进行按键反应, 且要求又快又准。为了掩盖实验的真实意图, 被试被告知只是一个反应时测验, 实验结束后告诉其反应时结果。在区组之间,被试可休息几分钟(图1)。
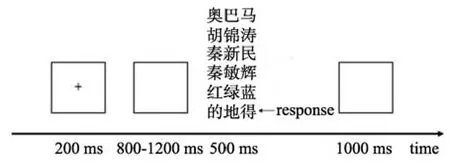
图1 实验中一个试次的实验流程
为了测量刺激的熟悉度和有效性, 要求被试在实验后对四类自我相关刺激的自我相关程度和熟悉度进行评定, 采用自评9点量表。在自我相关程度的评定中, “1”代表完全不相关, “9”代表非常相关。在熟悉度的评定中, “1”代表完全不熟悉, “9”代表非常熟悉。
2.6 脑电记录
使用 NeuroScan ERP记录与分析系统, 按国际10-20系统扩展的64导电极帽记录EEG。在线记录时将参考电极安置于左侧乳突位置, 离线后转为双侧乳突为参考电极, 双眼外侧安置电极记录水平眼电(HEOG), 左眼上下安置电极记录垂直眼电(VEOG)。滤波带通为 0.05~70 Hz, 采样频率为500 Hz/导, 头皮阻抗 < 5 kΩ。
2.7 ERP数据处理与统计
完成连续记录EEG后, 离线(off-line)处理数据,用NeuroScan软件校正VEOG, 并充分排除其他伪迹。本研究仅对偏差刺激呈现后的脑电数据进行分析, 分别对不同相关程度的自我参照刺激进行EEG叠加。波幅大于±80 μV者视为伪迹而自动剔除。分析时程(epoch)为刺激后600 ms, 基线为刺激呈现前200 ms。根据已有研究结果与本研究目的,对P2 (170~270 ms)和N2 (270~370 ms)的波幅和潜伏期以及P300 (400~500 ms)的平均波幅进行统计分析。选择15个电极位置, 进行四因素重复测量方差分析, 4个因素分别为2(被试人格类型:外倾性与控制组)×4 (自我参照刺激类型:自我高相关、中等自我相关、低自我相关和熟悉刺激) ×3(半球:左侧(F3, FC3, C3, CP3, P3), 中线(Fz, FCz, Cz, CPz, Pz),右侧(F4, FC4, C4, CP4, P4))×5 (脑区:额区(F3, Fz, F4),额中区(FC3, FCz, FC4), 中央区(C3, Cz, C4), 中顶区(CP3, CPz, CP4), 顶区(P3, Pz, P4))。方差分析的p
值采用 Greenhouse-Geisser法校正。脑电地形图由64导数据得出, 对其神经机制的探索采用偶极子溯源分析, 找到内隐自我参照加工程度效应所激活的大脑区域。3 结果
3.1 行为结果
实验后的评定表明自我相关刺激存在着一个显著的主效应,F
(3, 57)=38.05,p
< 0.001, η=0.32。事后比较表明高自我相关名字比中等自我相关名字评定得分更高[t
(19)=3.31,p
< 0.05], 而中等自我相关名字比低自我相关名字[t
(19)=3.08,p
<0.05]和熟悉名字[t
(19)=9.5,p
< 0.01]都要高些。此外, 低自我相关名字比熟悉名字的评定得分又要高一些,t
(19)=8.21,p
< 0.01。而四类自我相关刺激在熟悉度方面的评定没有发现显著的差异,F
(3, 57)=1.03,p
> 0.05。3.2 ERPs结果
如图2和图3, 在四类刺激中都发现了N1, P2,N2 和P3 成分。重复测量方差分析表明, N1在潜伏期和波幅上都没有发现显著的差异。
3.2.1 P2 (170~270 ms)
在P2的波幅上, 重复测量方差分析发现, 自我参照刺激类型的主效应显著,F
(3, 108)=4.04,p
<0.01, η=0.1。事后比较发现高自我相关的刺激比其他刺激诱发了更大的P2波幅, allF
s (1, 36) >3.59, allp
s < 0.05。重复测量方差分析发现, 自我参照刺激类型与脑区交互作用显著,F
(12, 432)=10.86,p
< 0.001, η=0.23]。简单效应分析表明高自我相关名字所诱发的P2的波幅在额区[allF
s (1, 36) >5.93, allp
s < 0.01], 额中区[allF
s (1, 36) > 7.39, allp
s < 0.001], 中央区[allF
s (1, 36) > 6.5, allp
s < 0.01]比其他刺激更大。其中额中区位置的效应最大。重复测量方差分析还发现, 自我参照刺激类型与半球交互作用显著,F
(6, 216)=5.18,p
< 0.001, η=0.13。简单效应分析表明高自我相关名字比其他刺激在右侧脑区[allF
s (1, 36) > 3.94, allp
s < 0.05]和中线位置[allF
s (1, 36) > 9.44, allp
s < 0.001]所诱发了更大的P2的波幅。其中中线位置的效益最大。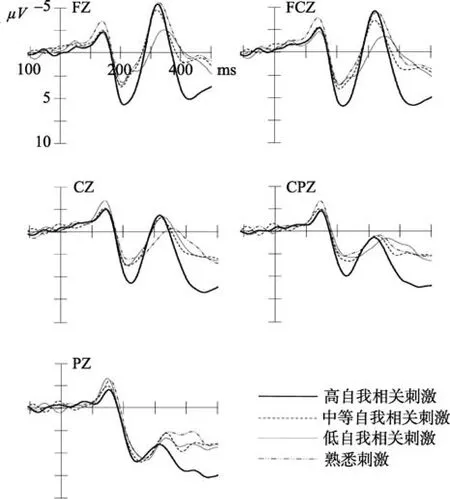
图2 外倾个体4种刺激在Fz、FCz、Cz、CPz和Pz电极点上的ERPs总平均图
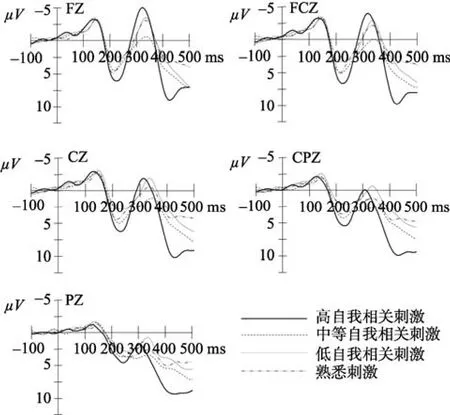
图3 控制组个体四类刺激在Fz、FCz、Cz、CPz和Pz上的ERPs总平均图
在P2的潜伏期上, 重复测量方差分析发现,自我参照刺激类型的主效应显著,F
(3, 108)=3.64,p
< 0.05, η=0.09。事后比较发现高自我相关的刺激比其他刺激诱发了更长的P2潜伏期, allF
s (1, 36) >2.93, allp
s < 0.05。重复测量方差分析发现, 自我参照刺激类型与脑区交互作用显著,F
(12, 432)=3.32,p
< 0.01, η=0.08]。简单效应分析表明高自我相关名字所诱发的P2的潜伏期在额区[allF
s (1, 36) >3.71, allp
s < 0.05], 中央区[allF
s (1, 36) > 3.75, allp
s < 0.05], 中顶区[allF
s (1, 36) > 4.15, allp
s < 0.01]和顶区[allF
s (1, 36) > 3.39, allp
s < 0.05]比其他刺激更长。其中中央区位置的效应最大。3.2.2 N2 (270~370 ms)


3.2.3 P3 (400~500 ms)
3.3 溯源分析结果
将两类被试的ERPs总平均导入具有标准MRI头像的Curry 6.0系统, 用sLORETA方法在三壳球模型中重建各任务在不同时段的头皮活动源。根据实验结果的ERPs成分, 将高自我相关刺激减去熟悉刺激, 中等自我相关刺激减去熟悉刺激和低自我相关刺激减去熟悉刺激, 所得差异波取P2(170~270 ms), N2 (270~370 ms)和P300 (400~500 ms)成分的数据进行定位分析。结果发现外倾特质被试的P300成分的偶极子溯源有较大差异, 在400~500 ms时间段上, 高自我相关刺激减去熟悉刺激差异波和中等自我相关刺激减去熟悉刺激差异波显示显著激活了布鲁德曼35区(BA35)的扣带前回和海马旁回, 低自我相关刺激减去熟悉刺激差异波显示显著激活了布罗德曼35区(BA35)海马旁回(见图4)。控制组被试的P300成分的偶极子溯源较为相似, 在400~500 ms时间段上, 高自我相关刺激减去熟悉刺激差异波显著激活了中央前回(BA6)和后扣带回(BA30), 中等自我相关刺激减去熟悉刺激差异波与低自我相关刺激减去熟悉刺激差异波都显著激活了中央前回(BA6) (见图5)。
4 讨论
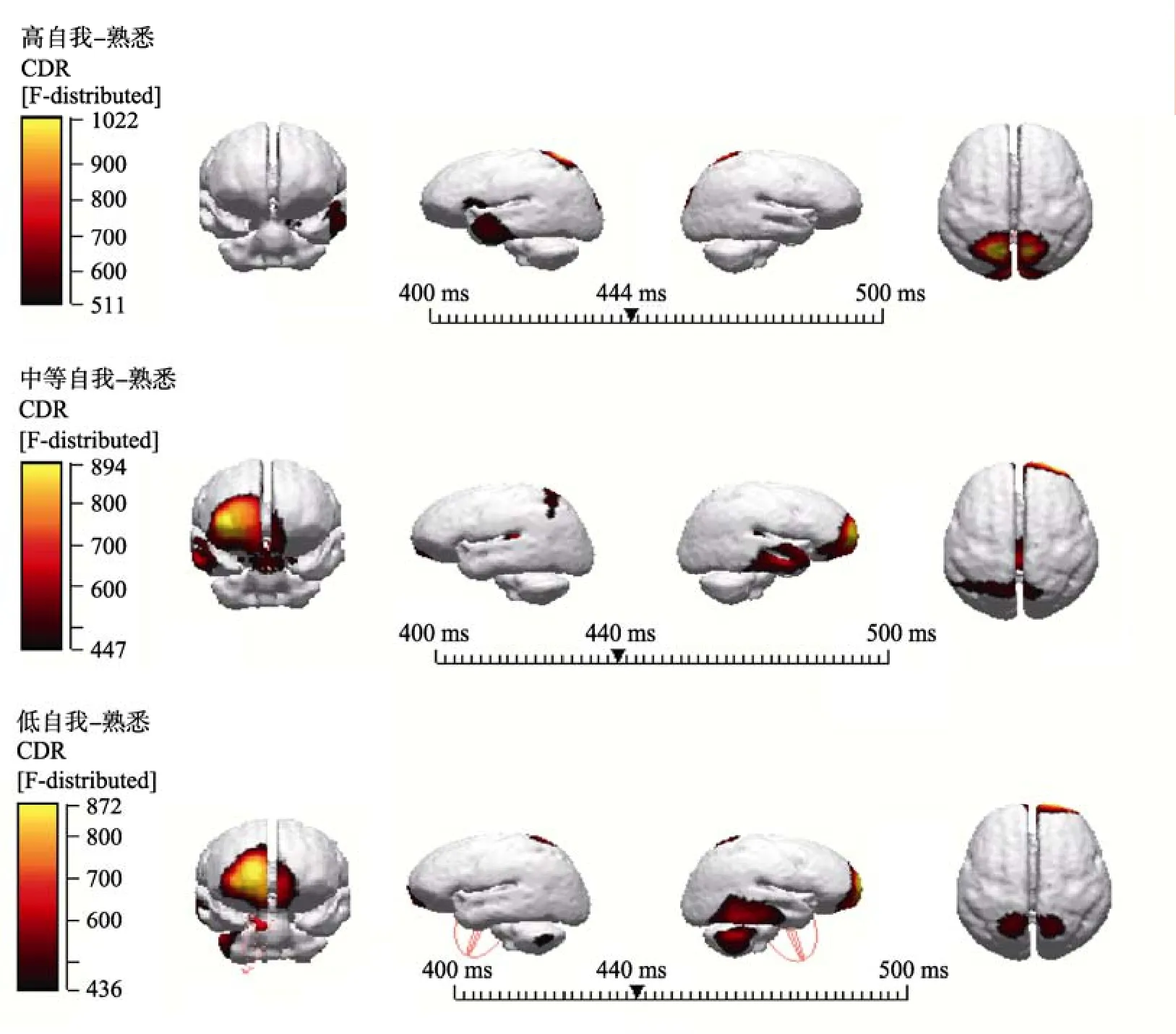
图4 外倾个体三类自我相关刺激与熟悉刺激相减差异波的P300溯源图
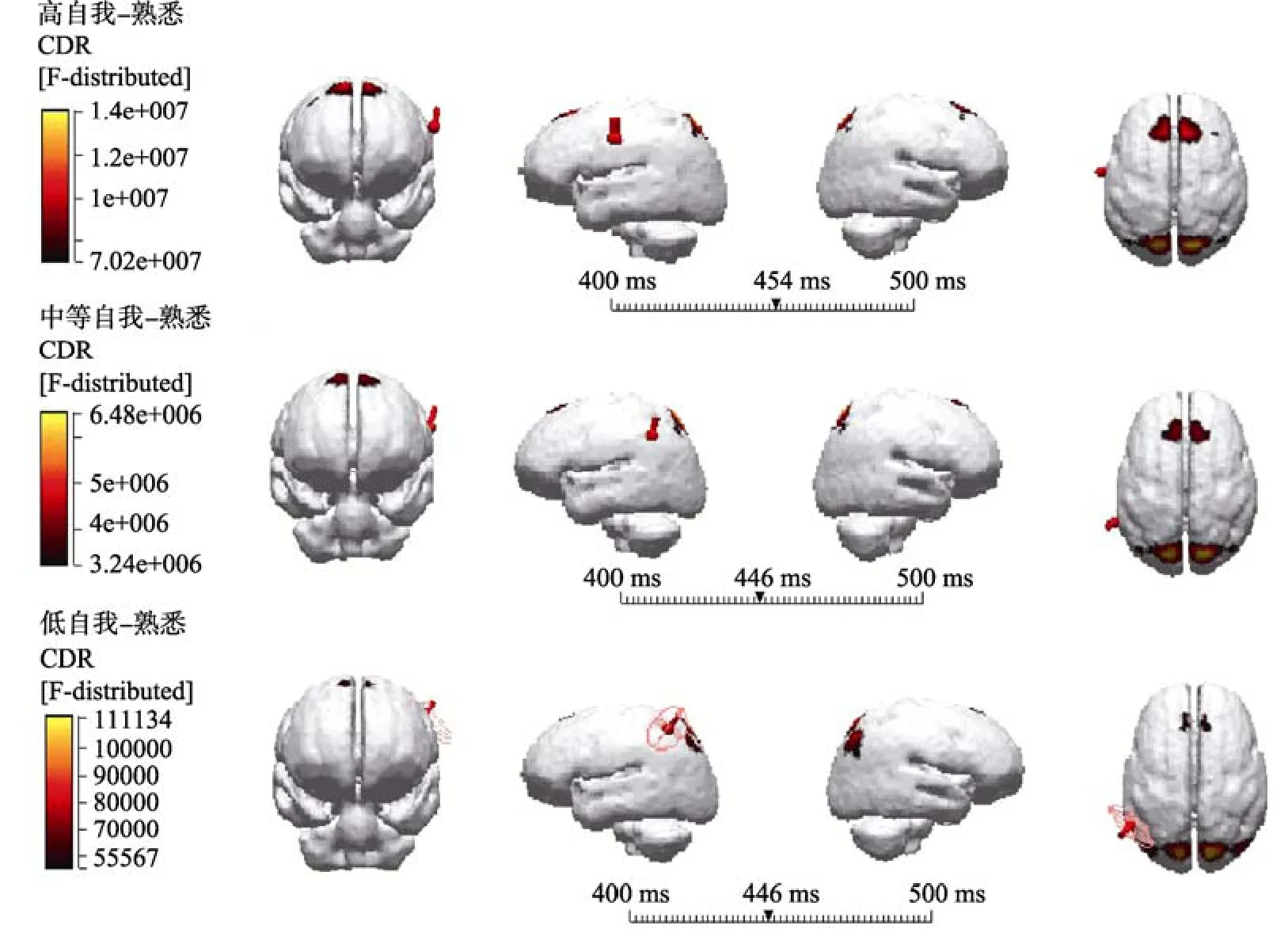
图5 控制组个体三类自我相关刺激与熟悉刺激相减差异波的P300溯源图
在当前的研究中, 研究结果表明四类自我相关刺激在N1的波幅和潜伏期上均没有发现显著的差异, 这可能是由于我们对刺激类型的尺寸大小、长度和复杂性等物理属性上做到了均衡。而在刺激呈现后, 我们在额区电极发现了明显的P2成分, 对比其他刺激, 高自我相关的名字得到了更大的P2波幅。有研究表明, 在额区位置发现的P2成分可能是对刺激特征的快速侦查, 易使人敏感的刺激会得更多的关注(Karayanidis & Michie, 1996; Thorpe,Fize, & Marlot, 1996)。此外, 还有研究表明生物重要性更强的刺激容易得到注意, 激发更大的P2波幅(Carretié, Mercado, Tapia, & Hinojosa, 2001)。还有证据表明, 个体的名字直接代表了他(她)自己,是核心自我之一, 因此在社会文化环境中有着很强的重要性(Chen et al., 2011;Perrin et al., 2005; Zhao, Wu,Zimmer, & Fu, 2011)。因此, 在当前的研究中, 高自我相关的名字诱发了更大的P2波幅, 可能是高自我相关的名字快速的得到了大脑的早期注意, 但是这种注意缺乏高级的认知加工和控制资源的分配(Del Cul, Baillet, & Dehaene, 2007; Hu, Wu, & Fu,2011)。此外, 对比其他的名字, 高自我相关的名字所诱发的P2潜伏期更长, 这与前人的研究保持了一致, 可能是由于自己名字的重要性比较高, 个体在识别高自我相关名字时投入了更多的注意资源,自我卷入程度更高, 从而诱发了更大的P2潜伏期(Fan et al., 2011, 2013; Zhao et al., 2009; Campanella et al.,2002)。中等自我相关名字和低自我相关名字的自我参照加工在这一阶段并没有发现显著的差异, 这可能是由于人脑对不同程度的自我参照刺激的加工和提取发生在稍晚的阶段。
研究结果在额区位置的电极还发现了比较明显的N2成分, 高自我相关的名字比其他名字诱发了更大的N2波幅。近期, 我们实验室有一些研究也发现了类似的效应, 其中一部分研究发现被试自我的名字比其他的名字诱发了更大的N2波幅(Fan et al., 2013; Zhao et al., 2009; 钟毅平等, 2014, 2015),还有研究发现相比熟悉刺激和陌生刺激, 自我参照刺激诱发了更大的N2波幅(Chen et al., 2014; Fan et al.,2011)。前人的研究中也有报告了比较明显的N2成分, 例如有研究表明被试自我的面孔比其他刺激诱发了更大的N2波幅(Herzmann, Schweinberger, Sommer,& Jentzsch, 2004)。N2通常被认为是一种非特殊的成分, 它与注意的转换机制相联系, 随后一般会有P3成分出现(Kiehl, Laurens, Duty, Forster, & Liddle,2001)。在被动接受刺激的情况下, 只有刺激足够突出或者发生了注意的转换, N2b/P3a都会被观察到(Näätänen, Simpson, & Loveless, 1982)。因此, 在本研究中, 高自我相关的名字虽然不是目标刺激, 但是由于其自我相关性更高, 在四类刺激中突出性更强, 很快抓住了人脑的注意。中等自我相关名字和低自我相关名字的自我参照加工在这一阶段并没有发现显著的差异, 这可能是由于在这一阶段人脑对不同程度的自我参照刺激的加工也是粗糙的, 更为精细的认知加工会发生在稍晚的阶段。在实验中,我们还发现了高自我相关名字比其他名字所诱发的N2潜伏期更短, 这可能是由于高自我相关的名字可能具有更好的心理突出性和生理重要性, 大脑识别起来更为容易, 消耗的认知资源更少一些(Campanella et al., 2002)。
更重要的是, 两组被试的高自我相关名字比其他名字诱发了更大的P3, 中等自我相关名字比熟悉名字诱发了更大的P3, 可见两组被试都存在自我参照加工的程度效应。但对比外倾性被试, 控制组被试的名字刺激诱发了更大的P3波幅, 所以控制组被试所诱发的自我参照加工程度效应更大。这些结果较好地验证了研究假设, 对比控制组个体,外倾性个体的自我参照加工的程度效应减弱了。这可能与外倾个体对正性情绪的敏感体验相关, 外倾个体在日常生活中体验到更多的正性情绪。例如,在早期的研究中, Costa和McCrae 发现外倾维度得分较高的个体在日常生活中报告出更多的正性情绪体验, 而且这种相关在一定程度上可预料10年后的正性情绪体验(Costa & McCrae, 1980, 1992)。这些研究表明外倾个体在日常生活中能够体验到更多的正性情绪, 并且很稳定。Larsen等人的研究结果表明, 被试的外倾维度得分能够很好地预测正性情绪的诱发。即在正性情绪诱发的情况下, 外倾性与更大的正性情绪反应相关(Larsen & Ketelaar, 1991)。Derryberry等人的行为研究用反应时证明了外倾性被试对正性刺激的注意解除存在困难, 即外倾性被试的注意从正性刺激出现的位置上移开, 所耗费的时间更长(Derryberry & Reed, 1994)。Canli和Amin等人的研究采用认知实验中的探针刺激范式, 并首次使用fMRI技术从视觉搜索所耗费的注意资源进行验证, 结果发现外倾个体更容易注意正性刺激,回避负性刺激(Amin et al., 2004; Canli et al., 2001)。这些研究表明外倾个体不但与正性情绪相关, 而且在日常生活中趋近正性刺激, 回避负性刺激。此外,还有Yuan等人的研究表明高度外倾个体在内隐情绪任务中, 不仅能够区分正性刺激和中性刺激, 还能进一步区分不同效价强度的正性刺激, 即高度外倾个体的ERP波幅在中性, 中等正性和极端正性三种情况下, 两两差异显著(Yuan, Yang, Meng, Yu, & Li,2008)。这些研究表明对比中间型个体和内倾个体,外倾个体对正性刺激的效价强度变化更加敏感, 具有能识别不同程度正性情绪的能力。总之, 外倾个体由于本身的一些特质, 如乐群, 爱好社交等, 使他们在日常生活中有更多的朋友和更为丰富的休闲生活, 能体验到更多且更为强烈的正性情绪。而这些强烈的正性情绪体验, 可能消耗更多的认知资源, 使得自我参照加工及其程度效应减弱了。
而在额区、额中区和中线, 我们还观察到了刺激条件具有显著差异的P3成分。在本研究中, 我们使用的是三类刺激的oddball, “红绿蓝” 是标准刺激, 比例最大, 占到所有trial的70%, “的地得”是目标刺激, 只占6%, 被试的任务是对目标刺激进行按键反应, 4种自我相关的名字为偏差刺激,各占6%, 比例很小, 所以在实验中观察到的P3,应该是一种新异P3成分。新异P3通常被认为是人脑在晚期阶段对小概率刺激加工的注意偏向(Campanella et al., 2002; Carretié, Hinojosa, Martín-Loeches, Mercado,& Tapia, 2004; Yuan et al., 2008)。对比低自我相关的刺激, 人脑在加工高自我相关的刺激时会投入更多的认知和控制加工资源。因此, 在实验中高自我相关的名字比中等自我相关的名字诱发了更大的P3波幅, 而中等自我相关的名字又比等自我相关的名字和熟悉的名字诱发了更大的P3波幅。这些结果证明了一个显著的自我参照加工的程度效应, 即高自我相关的刺激会比低自我相关的刺激得到更为深入和精细的加工。这些结果可能还表明了, 相比其他ERPs成分, P3成分可能反应了自我信息的总体和精细的加工过程, 能够作为自我参照加工程度效应的有效ERPs指标。
另外, 前人的研究报告了熟悉刺激比不熟悉的刺激诱发更大的P3波幅(Beauchemin et al., 2006; Bobes,Martín, Olivares, & Valdés-Sosa, 2000; Zhao et al., 2009)。如果刺激的熟悉程度不一样, 可能会对自我相关程度的研究造成污染。而我们的行为学评定结果已经将这种可能排除了, 所有的自我相关刺激和熟悉刺激在熟悉程度上没有显著的差异。因此, 我们在实验中所发现的P3可能无关熟悉度。而且, 它又与自我相关程度效应有着显著的正相关, 自我相关程度越高, P3的波幅越大, 而N2成分不存在这种相关和影响, 所以我们在实验中发现的P3可能不是小概率刺激或者新异刺激的指标, 而是反映了自我相关程度不同刺激的ERPs指标。
更有趣的是, 高自我相关名字比中等自我相关名字和低自我相关名字诱发了更明显的P2、N2和P3, 且这种效应在中线和额区位置最明显。而中等自我相关名字和低自我相关名字诱发的P2、N2和P3波幅在左侧脑区位置更大。这些结果与前人关于个体自我参照加工的右脑优势效应, 集体自我参照加工是左脑优势效应并不完全相同(Fan et al., 2011, 2013;Keenan, Wheeler, Gallup, & Pascual-Leone, 2000; Perrin et al., 2006; Zhao et al., 2009)。这种脑区的优势效应显示出了可能不仅仅体现在个体自我或集体自我刺激的不同, 而是刺激材料与自我相关程度不同可能激活的脑区就会不同(Fan et al., 2013)。而且偶极子溯源发现, 在400~500 ms时间段上, 外倾特质个体的高自我相关刺激减去熟悉刺激差异波和中等自我相关刺激减去熟悉刺激差异波显示显著激活了布鲁德曼35区(BA35)的扣带前回和海马旁回,低自我相关刺激减去熟悉刺激差异波显示显著激活了布罗德曼35区(BA35)海马旁回; 而控制组个体的高自我相关刺激减去熟悉刺激差异波显著激活了中央前回(BA6)和后扣带回(BA30), 中等自我相关刺激减去熟悉刺激差异波与低自我相关刺激减去熟悉刺激差异波都显著激活了中央前回(BA6)。这些研究结果表明刺激的相关程度越高,皮质中线结构(CMS)的激活程度越高, 且外倾特质个体的自我参照加工程度效应的神经机制与控制组个体的并不完全一致。前人的研究表明与非自我参照刺激相比, 自我参照刺激激活了皮质中线结构(CMS)的不同方面, 包括腹内侧前额叶皮质(vmPEC)、前扣带皮层(pACC)、背内侧前额叶皮质(dmPFC)、辅助运动区(Supplementary Motor Area SMA)和楔前叶(precuneus)等(Kjaer, Nowak, & Lou, 2002)。外倾特质个体的自我参照加工的神经机制除了激活了皮质中线结构, 还激活了海马旁回。这些神经基础可能凸显出了外倾特质个体与控制组个体的自我参照加工神经机制的不同之处, 海马旁回的激活可能是由于外倾人格个体的情绪易感性所造成的,外倾人格个体看到与自己相关的名字时搜索、回忆或产生了更多的正性情绪。这也为进一步的研究提供了一个方向, 在未来的研究中可以进一步来探讨不同人格特质个体的自我参照加工神经机制的差异是否是由刺激材料与自我相关程度的不同所引起的。
总的来说, 当前的实验借助高时间分辨率的ERPs技术, 不但重复了经典的自我参照效应, 而且拓展了前人的研究, 揭示了外倾特质被试自我参照加工程度效应的特性。对比控制组个体, 外倾性被试的自我参照加工的程度效应更微弱一些。
Amin, Z., Todd Constable, R., & Canli, T. (2004). Attentional bias for valenced stimuli as a function of personality in the dot-probe task.Journal of Research in Personality, 38
(1),15−23.Beauchemin, M., De Beaumont, L., Vannasing, P., Turcotte, A.,Arcand, C., Belin, P., & Lassonde, M. (2006). Electrophysiological markers of voice familiarity.European Journal of Neuroscience, 23
(11), 3081–3086.Bobes, M. A., Martín, M., Olivares, E., & Valdés-Sosa, M.(2000). Different scalp topography of brain potentials related to expression and identity matching of faces.Cognitive Brain Research, 9
, 249–260.Brédart, S., François, S., & Guimond, S. (2013). The effect of spontaneous self-reference on memory: A replication.L’Année psychologique, 113
(2), 161–167.Caharel, S., Courtay, N., Bernard, C., Lalonde, R., & Rebai, M.(2005). Familiarity and emotional expression influence an early stage of face processing: An electrophysiological study.Brain and Cognition, 59
(1), 96–100.Campanella, S., Gaspard, C., Debatisse, D., Bruyer, R., Crommelinck,M., & Guerit, J. M. (2002). Discrimination of emotional facial expressions in a visual oddball task: An ERP study.Biological Psychology, 59
(3), 171–186.Canli, T., Zhao, Z., Desmond, J. E., Kang, E., Gross, J., &Gabrieli, J. D. E. (2001). An fMRI study of personality influences on brain reactivity to emotional stimuli.Behavioral Neuroscience, 115
(1), 33–42.Carretié, L., Hinojosa, J. A., Martín-Loeches, M., Mercado, F.,& Tapia, M. (2004). Automatic attention to emotional stimuli: Neural correlates.Human Brain Mapping, 22
(4),290–299.Carretié, L., Mercado, F., Tapia, M., & Hinojosa, J. A. (2001).Emotion, attention and the "negativity bias", studied through event-related potentials.International Journal of Psychophysiology, 41
(1), 75−85.Chen, J., Yuan, J. J., Feng, T. Y., Chen, A. T., Gu, B. B., & Li,H. (2011). Temporal features of the degree effect in selfrelevance: Neural correlates.Biological Psychology, 87
(2),290–295.Chen, Y., Zhong, Y. P., Zhou, H. B., Zhang, S. M., Tan, Q. B.,& Fan, W. (2014). Evidence for implicit self-positivity bias:An event-related brain potential study.Experimental Brain Research, 232
, 985–994.Chen, Y., Zhong, Y. P., Zhou, H. B., Zhou, L. P., & Wang, X. Y.(2012). An ERP study on implicit self-positivity bias effect.Chinese Journal of Clinical Psychology, 20
(3), 297–300.[陈芸, 钟毅平, 周海波, 周路平, 王小艳. (2012). 内隐自我正面偏见效应的ERP研究.中国临床心理学杂志, 20
(3),297–300.]Costa, P. T., & McCrae, R. R. (1980). Influence of extraversion and neuroticism on subjective well-being: Happy and unhappy people.Journal of Personality and Social Psychology,38
(4), 668–678.Costa, P. T., Jr., & McCrae, R. R. (1990). Personality disorders and the five-factor model of personality.Journal of Personality Disorders, 4
(4), 362–371.Costa, P. T., & McCrae, R. R. (1992).Revised NEO personality inventory (NEO PI-R) and NEO five-factor inventory(NEO-FFI)
. Odessa, FL: Psychological Assessment Resources.Del Cul, A., Baillet, S., & Dehaene, S. (2007). Brain dynamics underlying the nonlinear threshold for access to consciousness.PLoS Biology, 5
(10), e260.Derryberry, D., & Reed, M. A. (1994). Temperament and attention: Orienting toward and away from positive and negative signals.Journal of Personality and Social Psychology,66
(6), 1128–1139.Eysenck, H. J. (1990). Genetic and environmental contributions to individual differences: The three major dimensions of personality.Journal of Personality, 58
(1), 245–261.Fan, W., Chen, J., Wang, X. Y., Cai, R. H., Tan, Q. B., Chen,Y., ... Zhong, Y. P. (2013). Electrophysiological correlation of the degree of self-reference effect.PLoS One, 8
(12),e80289.Fan, W., Zhang, Y. X., Wang, X., Wang, X. Y., Zhang, X. Y., &Zhong, Y. P. (2011). The temporal features of selfreferential processing evoked by national flag.Neuroscience Letters, 505
(3), 233–237.Höller, Y., Kronbichler, M., Bergmann, J., Crone, J. S.,Schmid, E. V., Golaszewski, S., & Ladurner, G. (2011).Inter-individual variability of oscillatory responses to subject's own name: A single-subject analysis.International Journal of Psychophysiology, 80
(3), 227–235.He, Y. Y., Yuan, J. J., Wu, Z. L., & Li, H. (2008). The valence strength of positive stimuli modulates the attention in extraverts study.Acta Psychologica Sinica, 40
(11), 1158– 1164.[何媛媛, 袁加锦, 伍泽莲, 李红. (2008). 正性情绪刺激效价强度的变化对外倾个体注意的调制作用.心理学报,40
(11), 1158–1164.]Herzmann, G., Schweinberger, S. R., Sommer, W., & Jentzsch,I. (2004). What's special about personally familiar faces? A multimodal approach.Psychophysiology, 41
(5), 688–701.Hu, X. Q., Wu, H. Y., & Fu, G. Y. (2011). Temporal course of executive control when lying about self- and other- referential information: An ERP study.Brain Research, 1369
, 149–157.Hu, Y. H., & Huang, M. E. (2006). Diverse consequences of neuroticism and extraversion on down-regulation of negative emotions.Acta Psychologica Sinica, 38
(4), 553–561.[胡艳华, 黄敏儿. (2006). 神经质和外倾的负情绪减弱调节特点.心理学报, 38
(4), 553–561.]Huang, M. E., & Guo, D. J. (2004). The influence of extraversion and neuroticism on emotions.Journal of Psychological Science, 26
(6), 1047–1051.[黄敏儿, 郭德俊. (2004). 外倾和神经质对情绪的影响.心理科学, 26
(6), 1047–1051.]Karayanidis, F., & Michie, P. T. (1996). Frontal processing negativity in a visual selective attention task.Electroencephalography and Clinical Neurophysiology,99
(1), 38–56.Keenan, J. P., Wheeler, M. A., Gallup, G. G., Jr., & Pascual-Leone, A. (2000). Self-recognition and the right prefrontal cortex.Trends in Cognitive Sciences, 4
(9), 338–344.Keyes, H., Brady, N., Reilly, R. B., & Foxe, J. J. (2010). My face or yours? Event-related potential correlates of selfface processing.Brain and Cognition, 72
(2), 244–254.Kiehl, K. A., Laurens, K. R., Duty, T. L., Forster, B. B., &Liddle, P. F. (2001). Neural sources involved in auditory target detection and novelty processing: An event-related fMRI study.Psychophysiology, 38
(1), 133–142.Kjaer, T. W., Nowak, M., & Lou, H. C. (2002). Reflective selfawareness and conscious states: PET evidence for a common midline parietofrontal core.NeuroImage, 17
(2), 1080–1086.Larsen, R. J., & Ketelaar, T. (1991). Personality and susceptibility to positive and negative emotional states.Journal of Personality and Social Psychology, 61
(1), 132– 140.Li, Z. Q., Wang, L., Zhang, H. C., & Liu, H. C. (2010). Personality traits and subjective well-being: The mediating role of emotion regulation.Journal of Psychological Science, 33
(1),165–167.[李中权, 王力, 张厚粲, 柳恒超. (2010). 人格特质与主观幸福感: 情绪调节的中介作用.心理科学, 33
(1), 165– 167.]Lou, Y. X., Cai, A. Y., Yang, J. M., & Yuan, J. J. (2014). The impact of introversion-extraversion on emotion regulations and the neurophysiological underpinnings.Advances in Psychological Science, 22
(12), 1855–1866.[娄熠雪, 蔡阿燕, 杨洁敏, 袁加锦. (2014). 内-外倾人格对情绪调节的影响及神经机制.心理科学进展, 22
(12),1855–1866.]Ma, Y., & Han, S. H. (2012). Functional dissociation of the left and right fusiform gyrus in self-face recognition.Human Brain Mapping, 33
(10), 2255–2267.Näätänen, R., Simpson, M., & Loveless, N. E. (1982). Stimulus deviance and evoked potentials.Biological Psychology,14
(1-2), 53–98.Perrin, F., Maquet, P., Peigneux, P., Ruby, P., Degueldre, C.,Balteau, E., ... Laureys, S. (2005). Neural mechanisms involved in the detection of our first name: A combined ERPs and PET study.Neuropsychologia, 43
(1), 12–19.Perrin, F., Schnakers, C., Schabus, M., Degueldre, C., Goldman,S., Brédart, S., ... Luxen, A. (2006). Brain response to one's own name in vegetative state, minimally conscious state,and locked-in syndrome.Archives of Neurology, 63
(4),562–569.Rogers, T. B., Kuiper, N. A., & Kirker, W. S. (1977). Self- reference and the encoding of personal information.Journal of Personality and Social Psychology, 35
(9), 677– 688.Sui, J., Zhu, Y., & Han, S. H. (2006). Self-face recognition in attended and unattended conditions: An event-related brain potential study.NeuroReport, 17
(4), 423–427.Tacikowski, P., Jednoróg, K., Marchewka, A., & Nowicka, A.(2011). How multiple repetitions influence the processing of self-, famous and unknown names and faces: An ERP study.International Journal of Psychophysiology, 79
(2),219–230.Tateuchi, T., Itoh, K., & Nakada, T. (2012). Neural mechanisms underlying the orienting response to subject's own name:An event-related potential study.Psychophysiology, 49
(6),786–791.Thorpe, S., Fize, D., & Marlot, C. (1996). Speed of processing in the human visual system.Nature, 381
(6582), 520–522.Watson, L. A., Dritschel, B., Obonsawin, M. C., & Jentzsch, I.(2007). Seeing yourself in a positive light: Brain correlates of the self-positivity bias.Brain Research, 1152
, 106–110.Yuan, J. J., Yang, J. M., Meng, X. X., Yu, F. Q., & Li, H.(2008). The valence strength of negative stimuli modulates visual novelty processing: Electrophysiological evidence from an event-related potential study.Neuroscience, 157
(3),524–531.Zhao, K., Wu, Q., Zimmer, H. D., & Fu, X. L. (2011).Electrophysiological correlates of visually processing subject's own name.Neuroscience Letters, 491
(2), 143–147.Zhao, K., Yuan, J. J., Zhong, Y. P., Peng, Y. S., Chen, J., Zhou,L. P., ... Ding, D. Q. (2009). Event-related potential correlates of the collective self-relevant effect.Neuroscience Letters, 464
(1), 57–61.Zhong, Y. P., Cai, R. H., Fan, W., Yang, Z. L., Yu, T., & Wu, Y.(2015). The degree of self-reference effect of introverted individuals.Journal of Psychological Science, 38
(3), 672–679.[钟毅平, 蔡荣华, 范伟, 杨子鹿, 喻涛, 吴云. (2015). 内倾特质个体的自我参照加工程度效应.心理科学, 38
(3),672–679.]Zhong, Y. P., Fan, W., Cai, R. H., Tan, Q. B., Xiao, L. H., Zhan,Y. L., ... Qin, M. H. (2014). The influence of positive emotion on the degree effect in self-referential processes:Evidence from ERPs.Acta Psychologica Sinica, 46
(3),341–352.[钟毅平, 范伟, 蔡荣华, 谭千保, 肖丽辉, 占友龙, ... 秦敏辉. (2014). 正性情绪诱导下的自我参照加工: 来自ERPS的证据.心理学报, 46
(3), 341–352.]Zhong, Y. P., Fan, W., Zhou, L. P., Xiao, L. H., Wang, X. Y.,Zhang, X. Y., ... Yan, Z. X. (2012). The spatiotemporal features of self-referential processing: Evidence of ERPs.Journal of Psychological Science, 35
(5), 1039–1046.[钟毅平, 范伟, 周路平, 肖丽辉, 王小艳, 张笑仪, ... 颜志雄. (2012). 音乐情境中自我参照加工的时间特征:来自ERPs的证据.心理科学, 35
(5), 1039–1046.]Zhou, A. B., Li, S. F., Shi, Z., Liu, P. R., Xia, R. X., Xu, K.P., ... Ren, D. Y. (2013). Searching the self: Encoding self-relevant possessive pronoun and Theta activity.Acta Psychologica Sinica, 45
(7), 790–796.[周爱保, 李世峰, 史战, 刘沛汝, 夏瑞雪, 徐科朋, ... 任德云. (2013). 寻找自我: 自我相关物主代词的编码与theta节律的活动.心理学报, 45
(7), 790–796.]猜你喜欢
医学食疗与健康(2022年4期)2022-05-13 16:54:39
癫痫与神经电生理学杂志(2022年6期)2022-02-10 13:55:46
意林(2021年2期)2021-02-08 08:32:47
中华养生保健(2020年7期)2020-11-16 01:13:30
价值工程(2018年25期)2018-09-26 10:14:36
试题与研究·教学论坛(2017年14期)2017-04-10 20:04:12
西南交通大学学报(2016年4期)2016-06-15 20:29:36
华北电力大学学报(自然科学版)(2016年3期)2016-04-25 05:06:56
思想政治教育研究(2014年6期)2015-04-23 23:59:45
断块油气田(2014年6期)2014-03-11 15:33:53
