
泽兰实蝇雌成虫的寄主选择行为
2016-01-27 13:17吴国星桂富荣
生物安全学报 2015年1期
马 沙, 吴国星, 谭 清, 桂富荣, 叶 敏, 高 熹*
1云南农业大学植物保护学院,云南 昆明 650201; 2云南省昭通市昭阳区植保植检站,云南 昭通 657000
泽兰实蝇雌成虫的寄主选择行为
马沙1,2, 吴国星1, 谭清1, 桂富荣1, 叶敏1, 高熹1*
1云南农业大学植物保护学院,云南 昆明 650201;2云南省昭通市昭阳区植保植检站,云南 昭通 657000
摘要:【背景】泽兰实蝇是紫茎泽兰的专性寄生天敌,已成为控制紫茎泽兰的重要因子。泽兰实蝇的寄主选择力对其控制紫茎泽兰至关重要,但相关报道还很少。【方法】采用“Y”形嗅觉仪在实验室内测定泽兰实蝇雌成虫对寄主植物——紫茎泽兰及其他植物的选择行为,以及紫茎泽兰受重金属胁迫后泽兰实蝇对其选择的变化。【结果】不同日龄的泽兰实蝇雌成虫对紫茎泽兰的选择性不具有显著差异;交配与否对雌成虫的寄主选择没有显著影响;雌成虫对紫茎泽兰的选择显著高于黄蒿、水葫芦和蓝花鼠尾草等非寄主植物;雌成虫对花期及机械损伤的紫茎泽兰选择更显著;紫茎泽兰经重金属镉(Cd)、铅(Pb)和锌(Zn)单一及复合处理后,泽兰实蝇雌成虫对其选择性显著降低。【结论与意义】泽兰实蝇雌成虫的寄主选择性随植物种类、寄主植物(紫茎泽兰)的生育期、健康状况及重金属胁迫而变化,这有助于更好地利用泽兰实蝇控制紫茎泽兰。
关键词:泽兰实蝇; 紫茎泽兰; 寄主选择
泽兰实蝇ProcecidocharesutilisStone是紫茎泽兰EupatoriumadenophorumSpreng的专性寄生天敌,多个国家曾先后利用该虫控制紫茎泽兰的蔓延(Dodd,1961; Rahman & Agarwal,1991)。我国于1984年首次从西藏聂拉木县将泽兰实蝇引进云南部分地区,随后其被引入四川、贵州等西南地区并形成自然种群(陈升碧和关德盛,1994; 唐川江和周俗,2003; 魏艺等,1989)。研究发现,泽兰实蝇通过幼虫蛀入紫茎泽兰幼嫩茎枝端部,蛀食并刺激被害部位形成膨大虫瘿,能抑制紫茎泽兰的种子产量、萌发率、分枝数及株高,并能抑制紫茎泽兰的光合作用及生物量的增加及分配(张智英等,1988; 郑征等,1989)。泽兰实蝇是否选择寄生紫茎泽兰,即对寄主的选择力,是其发挥以上控制作用的关键因素。目前国内外的研究多集中于泽兰实蝇寄生后对紫茎泽兰生长发育及控制效果等方面(陈旭东和何大愚,1990; 代聪等,1991; 何大愚等,1987; 李爱芳等,2006; 刘文耀等,1991; 杨光礼和孙元体,1993; 郑征等,1989; Bess & Haramoto,1958),未见有关泽兰实蝇对寄主选择力方面的研究报道。
另据马沙(2013)的初步调查发现,兰坪金顶铅锌矿附近区域泽兰实蝇的寄生率与较远非矿区相比呈现出较大差异,矿区泽兰实蝇的寄生率仅为非矿区的47.5%。经研究,在重金属矿区,紫茎泽兰体内的重金属积累量相对较高(李冰和张朝晖,2008; 汪文云和张朝晖,2008; 王吉秀等,2013; Zuetal.,2005)。分析认为:除了随食物链的传递而到达泽兰实蝇体内的重金属直接对泽兰实蝇产生毒性影响外,重金属还通过胁迫紫茎泽兰间接影响泽兰实蝇对紫茎泽兰的选择水平,然而目前国内外还缺乏相关方面的研究。鉴于此,本文利用“Y”型嗅觉仪,就泽兰实蝇雌成虫对紫茎泽兰及其他植物以及对受重金属胁迫的紫茎泽兰的选择性进行了相关研究。
1材料与方法
1.1 供试虫源
泽兰实蝇采自云南昆明呈贡区带有虫瘿的紫茎泽兰枝条,摘除部分枝叶后将虫瘿放入养虫笼(60 cm×60 cm×60 cm)中,待其自然羽化后,收集羽化的泽兰实蝇并将雌雄成虫分开单独饲养作为供试虫源。
1.2 供试仪器
“Y”型嗅觉仪:“Y”型管基部长10 cm、内径3 cm、两臂长约15 cm、夹角75°,两臂分别依次连有流量计、装植物用的玻璃干燥瓶、活性炭和气泵。
1.3 试验方法
1.3.1嗅觉仪法使用“Y”型嗅觉仪进行测定时,室温控制在(24±2)℃,相对湿度(65±5)%。将供试泽兰实蝇雌成虫放在“Y”型玻璃管的入口处,气流流速为250 mL·min-1,气流进入方式为吸入式。以试虫进入管口开始计时,每虫2 min,当试虫进入“Y”型嗅觉仪的一侧臂超过2/3、停留1 min时,即为选择。每只雌蝇仅测试1次,每次测试15头雌成虫,共设3次重复,每测完5头雌成虫立即调换处理与对照的位置,以避免几何位置对泽兰实蝇行为产生影响,每个处理测试完成后更换嗅觉仪。每更换1次处理,用95%乙醇清洗“Y型管”、味源瓶以及连接于其间的硅胶管,吹干后备用(李帅等,2014)。
1.3.2不同日龄泽兰实蝇雌成虫对紫茎泽兰选择的影响选取羽化后不同日龄的雌成虫,分别测定其对紫茎泽兰的选择行为,测定方法同1.3.1。
1.3.3交配行为对泽兰实蝇雌成虫选择紫茎泽兰的影响选择羽化当天已交配的雌成虫,逐日测定其对紫茎泽兰的选择行为,同时与羽化当天未交配的雌成虫进行对比,测定方法同1.3.1。
1.3.4泽兰实蝇雌成虫对寄主植物与非寄主植物的选择性比较黄蒿ArtemisiacarvifoliaBuch.-Ham. ex Roxb.、蓝花鼠尾草SalviafarinaceaBenth和水葫芦Eichhorniacrassipes(Mart.) Solme均采自云南农业大学校内,无虫害和机械损伤。试验设置3种气味源组合:紫茎泽兰vs黄蒿、紫茎泽兰vs蓝花鼠尾草、紫茎泽兰vs水葫芦,观察3日龄未交配泽兰实蝇雌成虫对其的选择行为,测定方法同1.3.1。
1.3.5泽兰实蝇雌成虫对不同健康状况和生育期的紫茎泽兰的选择性比较植株处理:(1)健康植株:没有受到任何机械损伤和虫害的生长健壮的植株;(2)机械损伤植株:供试前24 h,用打孔器(直径3 mm)在供试植株每个叶片上打孔(每片叶1~2个孔)。试验设置2组气味源组合:机械损伤紫茎泽兰vs健康紫茎泽兰、花期紫茎泽兰vs生长期紫茎泽兰,观察3日龄未交配泽兰实蝇雌成虫对其的选择行为,测定方法同1.3.1。
1.3.6铅(Pb)、锌(Zn)和镉(Cd)处理紫茎泽兰后对泽兰实蝇雌成虫选择行为的影响试验前将紫茎泽兰种子播种于温室内,温度(25±2)℃,光照比14L∶10D,相对湿度(75±5)%,每7 d浇水一次,待长至8 cm株高时移栽到塑料花盆中(直径8 cm)。
试验在云南农业大学温室内进行,试验用土壤采自云南农业大学校内试验地表层土壤。Pb胁迫处理水平为4000 mg·kg-1,Pb以Pb(NO3)2的形式加入;Zn胁迫处理水平为4000 mg·kg-1,Zn以ZnSO4·7H2O形式加入;Cd胁迫处理水平为1000 mg·kg-1,Cd以CdCl2·2.5H2O形式加入;Pb+Cd复合处理水平为2000 mg·kg-1Pb+500 mg·kg-1Cd;Zn+Cd复合处理水平为2000 mg·kg-1Zn+500 mg·kg-1Cd;Pb+Zn复合处理水平为2000 mg·kg-1Pb+2000 mg·kg-1Zn;Pb+Zn+Cd复合处理水平为2000 mg·kg-1Pb+2000 mg·kg-1Zn+500 mg·kg-1Cd。以上处理分别加入土壤后,拌匀,以不加入重金属的土壤作为空白对照,稳定2周后移栽紫茎泽兰幼苗。
选取大小、高度一致(8 cm)的紫茎泽兰幼苗,移栽到每个浓度处理中,每个处理栽种20盆,每盆3株,每周浇1次水。
紫茎泽兰Pb、Zn、Cd含量测定:生长2个月后,选取大小、高度一致的紫茎泽兰,参照汪文云和张朝晖(2008)、Zuetal.(2005)的方法,先用自来水将紫茎泽兰植物体洗净,再用去离子水漂洗干净后,依次烘干、粉碎、研磨、硝化(HClO4∶HNO3=1∶5,v/v),用TAS-990原子吸收分光光度计测定样品中的Pb、Zn、Cd含量。
泽兰实蝇趋向行为测定:试验设置7组气味源组合:Pb处理紫茎泽兰vs对照、Zn处理紫茎泽兰vs对照、Cd处理紫茎泽兰vs对照、Pb-Cd处理紫茎泽兰vs对照、Zn-Cd处理紫茎泽兰vs对照、Pb-Zn处理紫茎泽兰vs对照、Pb-Zn-Cd处理紫茎泽兰vs对照。测定3日龄未交配雌成虫对以上气味源的选择行为,方法同1.3.1。
1.4 数据处理
利用DPS软件(唐启义和冯明光,2002)对不同发育日龄的各指标进行统计与方差分析,并以Duncan′s新复极差法比较各时间段内指标间的显著性差异。
2结果与分析
2.1 不同日龄泽兰实蝇雌成虫对紫茎泽兰选择的影响
由图1可知,不同日龄雌成虫均更显著趋向于紫茎泽兰的挥发物;且3日龄雌成虫对紫茎泽兰的选择性最好,但不同日龄雌成虫之间的选择性无显著差异(F=2.095,P=0.1563)。
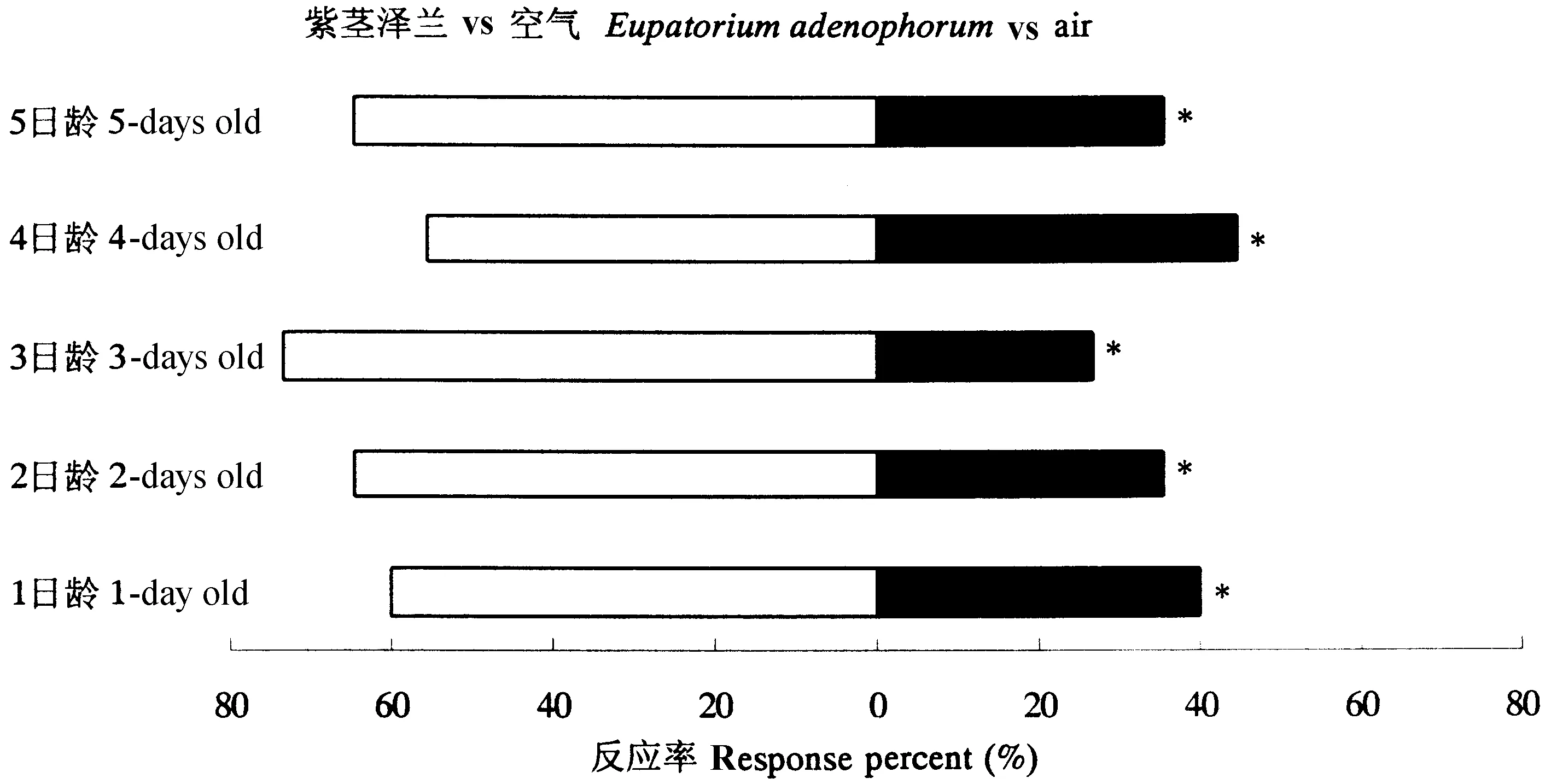
图1 不同日龄未交配雌成虫对紫茎泽兰选择的差异
2.2 交配行为对泽兰实蝇雌成虫选择紫茎泽兰的影响
如图2所示,1~5日龄的成虫,无论交配与否,其对紫茎泽兰的选择均超过对照,且日龄大小和交配行为对其选择紫茎泽兰的反应率无显著影响。
2.3 泽兰实蝇雌成虫对寄主植物与非寄主植物的选择性比较
如图3所示,泽兰实蝇雌成虫对紫茎泽兰的选择率极显著高于水葫芦、黄蒿和蓝花鼠尾草(P<0.01)。
2.4 泽兰实蝇雌成虫对不同健康状况和生育期的紫茎泽兰的选择性比较
如图4所示,泽兰实蝇雌成虫对机械损伤植株的选择率远高于健康植株,二者差异极显著(F=37.5,P=0.0036);雌成虫对花期紫茎泽兰的选择率显著高于生长期(F=13.5,P=0.0213)。

图2 交配行为对泽兰实蝇雌成虫选择紫茎泽兰的影响
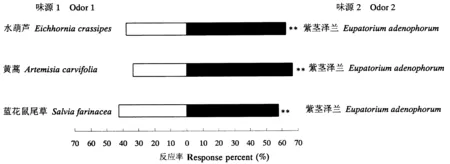
图3 泽兰实蝇雌成虫对寄主植物与非寄主植物的选择差异
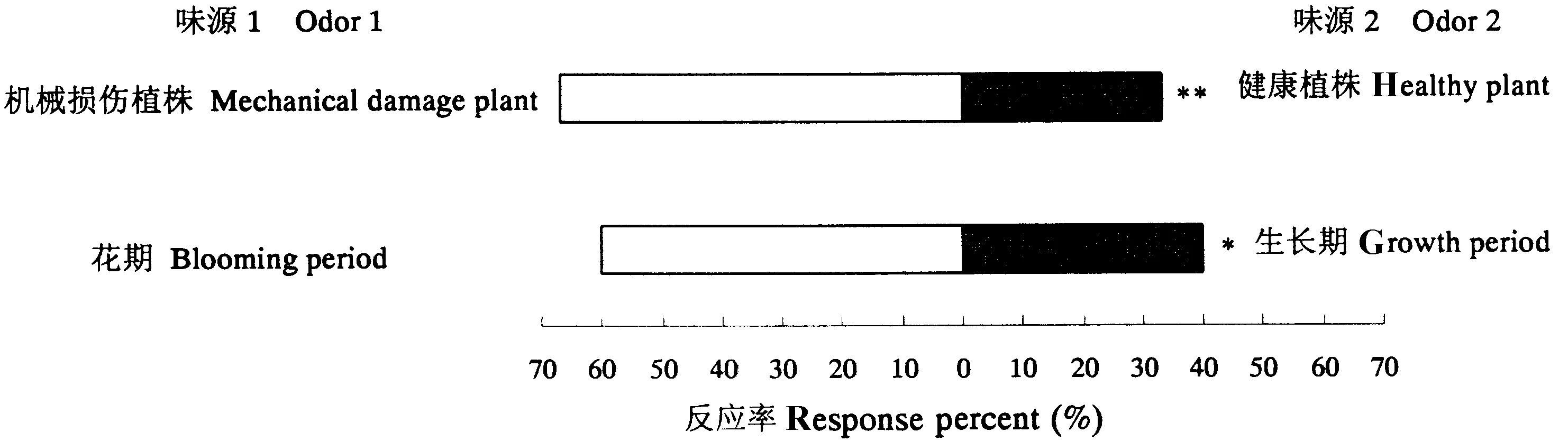
图4 泽兰实蝇对花期及机械损伤紫茎泽兰的选择差异
2.5 Pb、Zn和Cd处理紫茎泽兰后对泽兰实蝇雌成虫选择的影响
当采用Pb、Zn、Cd及这几种重金属复合处理时,紫茎泽兰未表现出受害症状,长势良好。紫茎泽兰对重金属Cd、Pb和Zn具有很强的吸收与忍耐能力,其中以Zn在体内的累积率最高,Pb次之,Cd的累积率最低(表1)。
如图5所示,重金属Pb、Zn和Cd单一处理紫茎泽兰后,泽兰实蝇对其趋向选择性显著低于未经重金属处理的植株;而经Pb-Cd、Zn-Cd、Pb-Zn和Pb-Zn-Cd复合处理紫茎泽兰后,雌蝇对其趋向选择与未经重金属处理的植株相比具极显著差异。
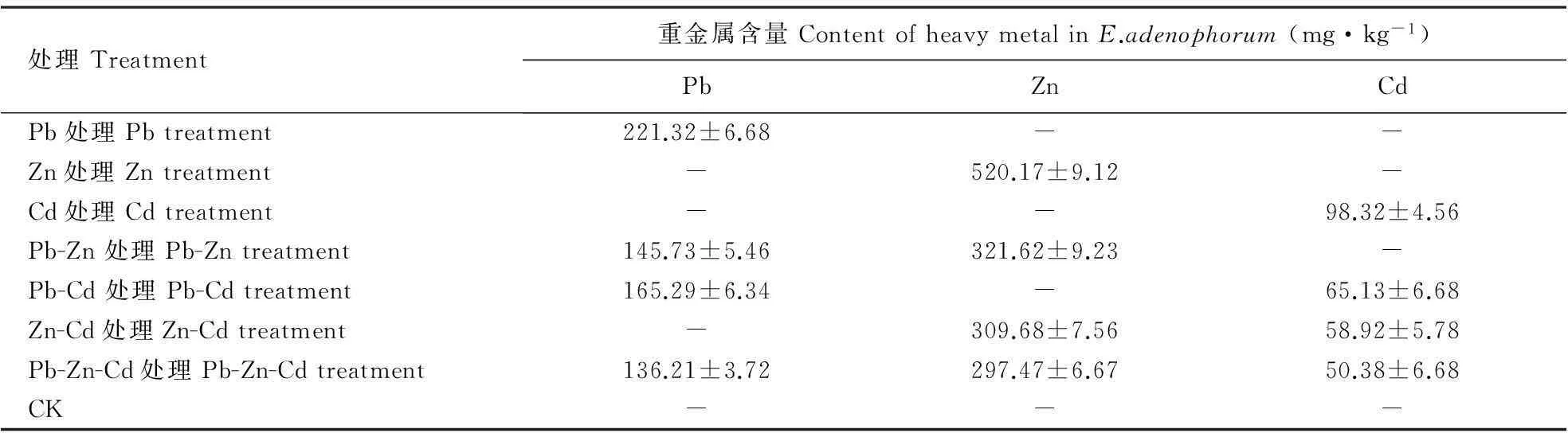
表1 Pb、Zn和Cd在紫茎泽兰中的生物累积
表中数据为平均值±标准误。
The date in the table are means±SE.
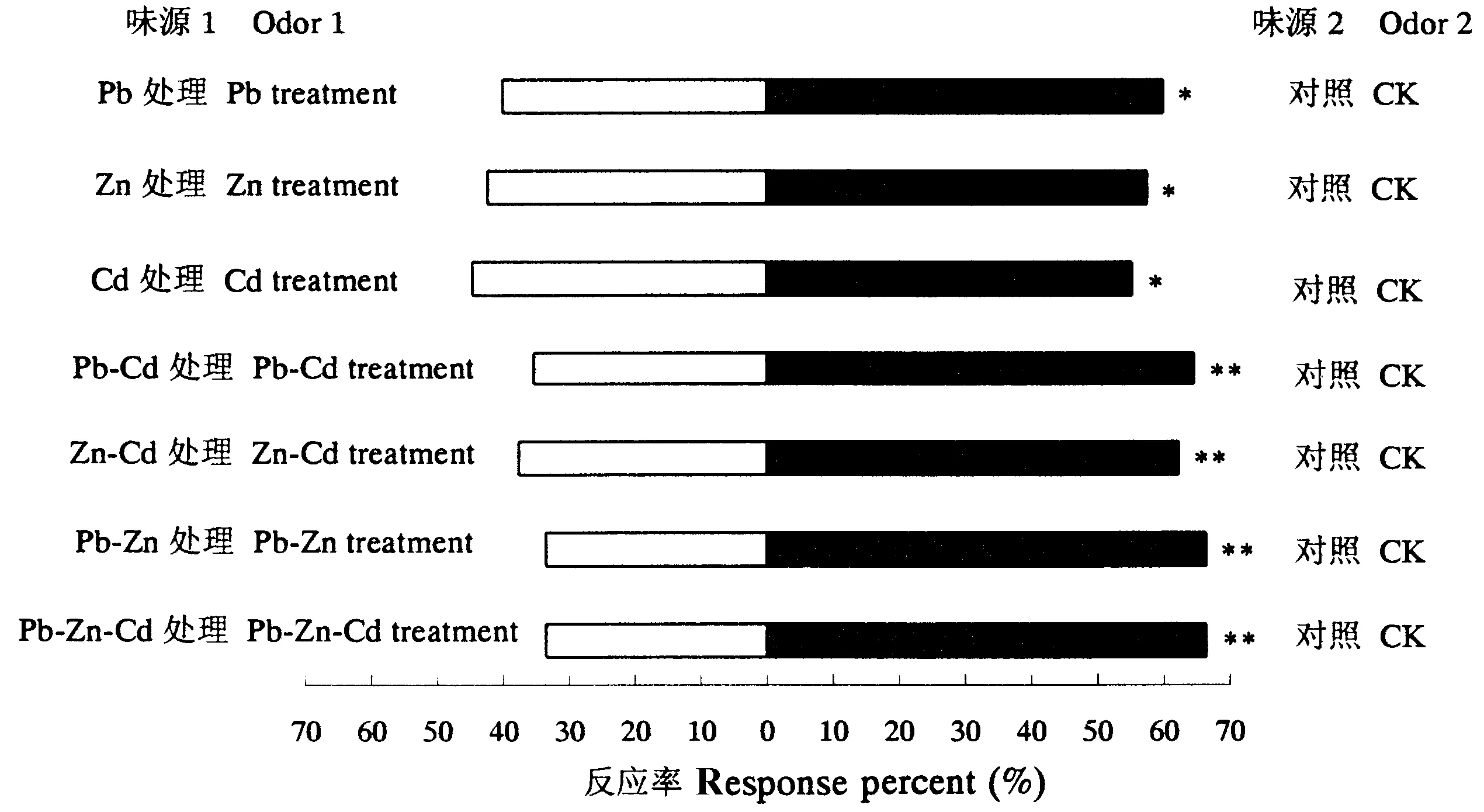
图5 Pb、Zn和Cd处理紫茎泽兰后泽兰实蝇对其选择的差异
3讨论
研究结果表明,羽化后不同日龄的雌成虫在选择紫茎泽兰上无显著差异,且交配与否也对其选择寄主无影响,这可能是因为泽兰实蝇雌成虫在初羽化时即达到性成熟,因此在成虫阶段对寄主的选择较一致。泽兰实蝇专性寄生,对其他植物不敏感,雌成虫对紫茎泽兰的选择率极显著高于其生境周围的杂草如水葫芦、黄蒿和蓝花鼠尾草,这更验证了泽兰实蝇食性的专一。
寄主植物挥发物是植物—昆虫间化学通讯的媒介,诱导昆虫产生多种行为,对植食性昆虫产卵、取食等活动中的寄主选择行为产生重要影响,是植食性昆虫进行寄主定位的利他素(杜家纬,2001; 康乐和Hopkins,2004)。植食性昆虫在寻找寄主阶段,主要通过嗅觉感受器对寄主植物特异性的气味进行识别而选择植物。例如,十字花科植物挥发物烯丙基异硫氰酸酯引诱许多取食十字花科植物的昆虫(娄永根和程家安,1997),马铃薯气味能够引诱马铃薯甲虫LeptinotarsadecemlineataSay(Visser & Ave,1978),苹果气味能够引诱苹果实蝇RhagoletispomonellaWalsh(Fein & Reissig,1982)等。从植食性昆虫的寄主植物中寻找害虫取食及产卵的它感信息物,并运用其来诱杀害虫,已有许多成功范例(Bauretal.,1993; Linetal.,1992; Su,1993)。本研究表明,泽兰实蝇对花期和有机械损伤的植株更具有趋性,因此花期和机械损伤后的植株挥发物有待进一步研究,其中的某些有效成分能开发成提高泽兰实蝇寄生率的引诱物。
紫茎泽兰是一种适生性很强的杂草,在矿区分布极为广泛。在室内用重金属胁迫紫茎泽兰后发现,泽兰实蝇对受单一重金属胁迫的紫茎泽兰的选择水平显著下降,对2种或2种以上重金属复合处理的紫茎泽兰的选择极显著下降,经初步分析,应是寄主挥发物在组成或是比例上发生改变而引起的,为此还待进一步对重金属处理前后的挥发物进行成分鉴定和定量分析。
参考文献
陈升碧, 关德盛. 1994. 泽兰实蝇生物学特性观察及生物防治. 西南农业学报, 7(4): 98-102.
陈旭东, 何大愚. 1990. 泽兰实蝇对紫茎泽兰的控制作用及其评价研究. 杂草学报, 4(3): 1-6.
代聪, 魏艺, 何大愚. 1991. 泽兰实蝇控制紫茎泽兰试验研究. 杂草学报, 5(1): 24-29.
二是重大水利项目建设推进有力。南水北调中线主体工程邯石段年内基本完工,配套工程全面开工建设,确保2014年汛后与主体工程同步建成。引黄入冀补淀工程项目建议书已批复,力争明年开工建设。承德双峰寺水库枢纽工程先后组织了监理和大坝主体工程招标投标,主体工程已全面开工建设。
杜家纬. 2001. 植物—昆虫间的化学通讯及其行为控制. 植物生理学报, 27(3): 193-200.
何大愚, 刘伦辉, 荆桂芬, 魏艺. 1987. 泽兰实蝇的安全性试验. 中国生物防治, 3(1): 1-3.
康乐, Hopkins T L. 2004. 黑蝗初孵蝗蝻对植物气味和植物挥发性化合物的行为和嗅觉反应. 科学通报, 49(1): 81-85.
李爱芳, 高贤明, 党伟光, 黄荣祥, 邓祖平, 唐和春. 2006. 泽兰实蝇寄生状况及其对紫茎泽兰生长与生殖的影响. 植物生态学报, 30(3): 496-503.
李冰, 张朝晖. 2008. 烂泥沟金矿区紫茎泽兰对重金属的富集特性及生态修复分析. 黄金, 29(8): 47-50.
李帅, 陈文龙, 金道超, 杨洪. 2014. 不同水稻挥发物对稻虱红螯蜂雌蜂的引诱作用. 植物保护学报, 41(2): 203-209.
刘文耀, 刘伦辉, 和爰军. 1991. 泽兰实蝇对紫茎泽兰生长发育及生物量分配影响的研究. 生态学报, 11(3): 291-293.
娄永根, 程家安. 1997. 植物—植食性昆虫—天敌三营养层次的相互作用及其研研究方法. 应用生态学报, 8(3): 325-331.
马沙. 2013. 镉、铅和锌在土壤—紫茎泽兰—泽兰实蝇系统中的累积及对泽兰实蝇的毒性研究. 昆明: 云南农业大学.
唐启义, 冯明光. 2002. 实用统计分析及其DPS数据统计分析系统. 北京: 科学出版社.
汪文云, 张朝晖. 2008. 贵州水银洞紫茎泽兰重金属元素测定与分析. 植物研究, 28(6): 760-763.
王吉秀, 高熹, 马沙, 吴国星. 2013. Cd、Pb和Zn在土壤—紫茎泽兰—泽兰实蝇系统中的生物富集效应研究. 中国生态农业学报, 21(7): 877-882.
魏艺, 张智英, 何大愚. 1989. 泽兰实蝇人工繁殖技术. 生物防治通报, 5(1): 41-42.
杨光礼, 孙元体. 1993. 泽兰实蝇对紫茎泽兰抑制效果的初步观察. 杂草科学, (2): 15.
张智英, 魏艺, 何大愚. 1988. 泽兰实蝇生物学特性的初步研究. 生物防治通报, 4(1): 10-13.
郑征, 唐继武, 刘文耀, 陈旭东. 1989. 泽兰实蝇对紫茎泽兰生长及光合作用影响的研究. 杂草学报, 3(2): 21-23.
Bess H A and Haramoto F H. 1959. Biological control of PamakaniEupatoriumadenophorumin Hawii by a Tephritid gall fly,Procecidocharesutilis. Ⅱ. Population studies of the weed, the fly and the parasites of the fly.Ecology, 40: 244-249.
Baur R, Feeny P and Stadler E. 1993. Oviposition stimulatants for the black swallowtail butterfly: identification of electrophysiologically active compounds in carrot volatiles.JournalofChemicalEcology, 19: 919-937.
Dodd A P. 1961. Biological control ofEupatoriumadenophorumin Queensland.AustralianJournalofScience, 23: 356-365.
Fein B L and Reissig G H I. 1982. Identification of apple volatile attraction to the apple maggotRhagoletispomonella.JournalofChemicalEcology, 8: 1473-1487.
Lin H, Phelan P L and Bartelt R. 1992. Synergism between synthetic food odors and the aggregation pheromone for attractingCarpophiluslugubrisin the field.EnvironmentalEntomology, 21: 156-159.
Rahman O and Agarwal M L. 1991. Biological control of crofton weed (EupatoriumadenophorumSpreng) by a fruit flyProcecidocharesutilisStone in eastern Himalayas.IndianJournalofWeedScience, 22: 98-101.
Su J Q. 1993. Using carrot flower trap and kill cotton bollworm moths.PlantProtection, 19: 23-27.
Visser J H and Ave D A. 1978. General green leaf volatiles in the olfactory orientation of the Colorado beetle,Leptinotarsadecemlineata.JournalofChemicalEcology, 24: 738-749.
Zu Y Q, Li Y, Chen J J, Chen H Y, Qin L and Schvartz C. 2005. Hyperaccumulation of Pb, Zn and Cd in herbaceous grown on lead-zinc mining area in Yunnan, China.EnvironmentInternational, 31: 755-762.
(责任编辑:郭莹)
Host selection behavior of female adults ofProcecidocharesutilis
(Diptera: Tephritidae)
Sha MA1,2, Guo-xing WU1, Qing TAN1, Fu-rong GUI1, Min YE1, Xi GAO1*
1CollegeofPlantProtection,YunnanAgriculturalUniversity,Kunming,Yunnan650201,China;
2ZhaoyangPlantProtectionandQuarantineStation,Zhaotong,Yunnan657000,China
Abstract:【Background】 Procecidochares utilis Stone is an obligatory parasite of Eupatorium adenophorum Spreng, and it has become an important factor in the control of E.adenophorum. The host selection of P.utilis adults is very important to control the growth of E.adenophorum, but few related studies are reported at present. 【Method】 The selection behavior of P.utilis female adults was tested using a Y-tube olfactometer on E.adenophorum, a selection of plants and E.adenophorum plants that were exposed to heavy metals. 【Result】 The selection behavior of P.utilis female adults of different ages on E.adenophorum showed no significant difference. Mating or not had no significant effect on host selection for the female adults. The choice of female adults on E.adenophorum was significantly higher than that of Artemisia carvifolia, Eichhornia crassipes and Salvia farinacea. The female adults preferred E.adenophorum in bloom or with mechanical damage. The selectivity of female adults decreased significantly if E.adenophorum plants were first exposed to Cd, Pb and Zn or a combination of them. 【Conclusion and significance】 The study showed selection behavior of P.utilis varied with plant species and development stage, health or level of contamination of the plants. This information can help establish a theoretical foundation for developing control mechanisms of E.adenophorum by using P.utilis.
Key words:Procecidochares utilis; Eupatorium adenophorum; host selection
DOI:10.3969/j.issn.2095-1787.2015.01.006
通讯作者*(Author for correspondence), E-mail: junhuang1981@aliyun.com
作者简介:黄俊, 男, 助理研究员, 博士。 研究方向: 入侵生物学与园林害虫综合治理
基金项目:浙江省自然科学基金(LQ14C140004)
收稿日期(Received): 2014-12-19接受日期(Accepted): 2015-01-13
