
食用菌体内抗氧化酶活性影响因素的研究进展
2015-12-27 00:50崔世瑞冯志勇陈明杰
微生物学杂志 2015年4期
崔世瑞, 冯志勇, 陈明杰, 汪 虹, 陈 辉*
(1.上海市农业科学院食用菌研究所 国家食用菌工程技术中心,上海 201403;2.南京农业大学 生命科学学院,江苏 南京 210095)
食用菌体内抗氧化酶活性影响因素的研究进展
崔世瑞1,2, 冯志勇1, 陈明杰1, 汪 虹1, 陈 辉1*
(1.上海市农业科学院食用菌研究所 国家食用菌工程技术中心,上海 201403;2.南京农业大学 生命科学学院,江苏 南京 210095)
食用菌的菌丝和子实体在正常生长发育中会产生活性氧,但是在环境胁迫下或者贮藏运输中活性氧会大量增加,打破了活性氧的平衡,必须依靠抗氧化酶系对抗其有害作用。介绍了抗氧化酶的种类及其作用,综述了食用菌不同部位、不同发育阶段等自身因素,及温度、金属离子、采后保鲜技术等外界环境因素对抗氧化酶活性的影响,并对食用菌抗氧化酶研究前景进行展望。
食用菌;抗氧化酶;活性变化
活性氧(reactive oxygen species,ROS)是一种化学性质活泼、氧化能力很强的含氧物质的总称,主要包括O2-·、RO-、ROO-、H2O2、ROOH等,活性氧的积累可以导致膜脂过氧化、碱基突变、DNA突变、蛋白质的损伤,对细胞膜引起不可逆的损伤,导致衰老[1-2],需氧生物体在生命活动中,细胞内的线粒体、叶绿体以及与质膜相关的电子传递系统会产生活性氧,但是机体内存在清除自由基和活性氧的机制,主要分为酶促和非酶促清除机制,其中超氧化物歧化酶(superoxide dismutase, SOD)、过氧化氢酶(catalase, CAT)、过氧化物酶(peroxidase, POD)等是酶促清除机制的主要酶,抗坏血酸(ascorbic acid, ASA)和还原性谷胱甘肽(reduced glutathione,GHS)等是非酶清除机制中的主要物质[3],因此机体内活性氧的积累和清除处于动态的平衡中。但是在逆境胁迫下,体内的活性氧数量急剧增加,机体内的抗氧化酶迅速增加产生相应的应激刺激。近几年关于抗氧化酶的报道多集中在玉米、果蔬等植物中[4-6],而在食用菌中相关的报道较少,本文阐述了食用菌菌丝体和子实体内影响抗氧化酶活性的因素及其研究热点。
1 食用菌抗氧化酶的种类及其功能
抗氧化酶类包括对活性氧直接起作用的SOD、CAT、抗坏血酸过氧化物酶(aseorbate pero xidase,APX)以及作用范围较广的POD等。SOD是生物体内特异清除超氧阴离子自由基的酶,是一种含金属的酶,根据金属辅助因子的不同可以分为Cu/Zn-SOD、Fe-SOD、Mn-SOD三种类型[7],它能催化O2-·使其发生歧化反应,生成O2和 H2O2,H2O2又在CAT的作用下生成无毒的H2O 和O2,从而起到抗衰老作用[8]。CAT是一种包含血红素的4聚体酶,在细胞中主要存在于过氧化体中,它与SOD协同作用可清除植物体内潜在危害的O2-·和H2O2,从而最大限度地减少了-OH的形成[9]。POD是一种以铁卟啉为辅基的酶类,其催化的反应除了需要H2O2以外还需要供电子体(AH2) 的存在,只要有足量的供电子体,POD在低浓度H2O2的情况下也可以发挥高效的催化作用[10]。APX在抗坏血酸-谷胱甘肽循环中,以抗坏血酸作为电子受体,发挥清除H2O2的作用[11]。图1为抗氧化酶系统清除ROS的原理图。
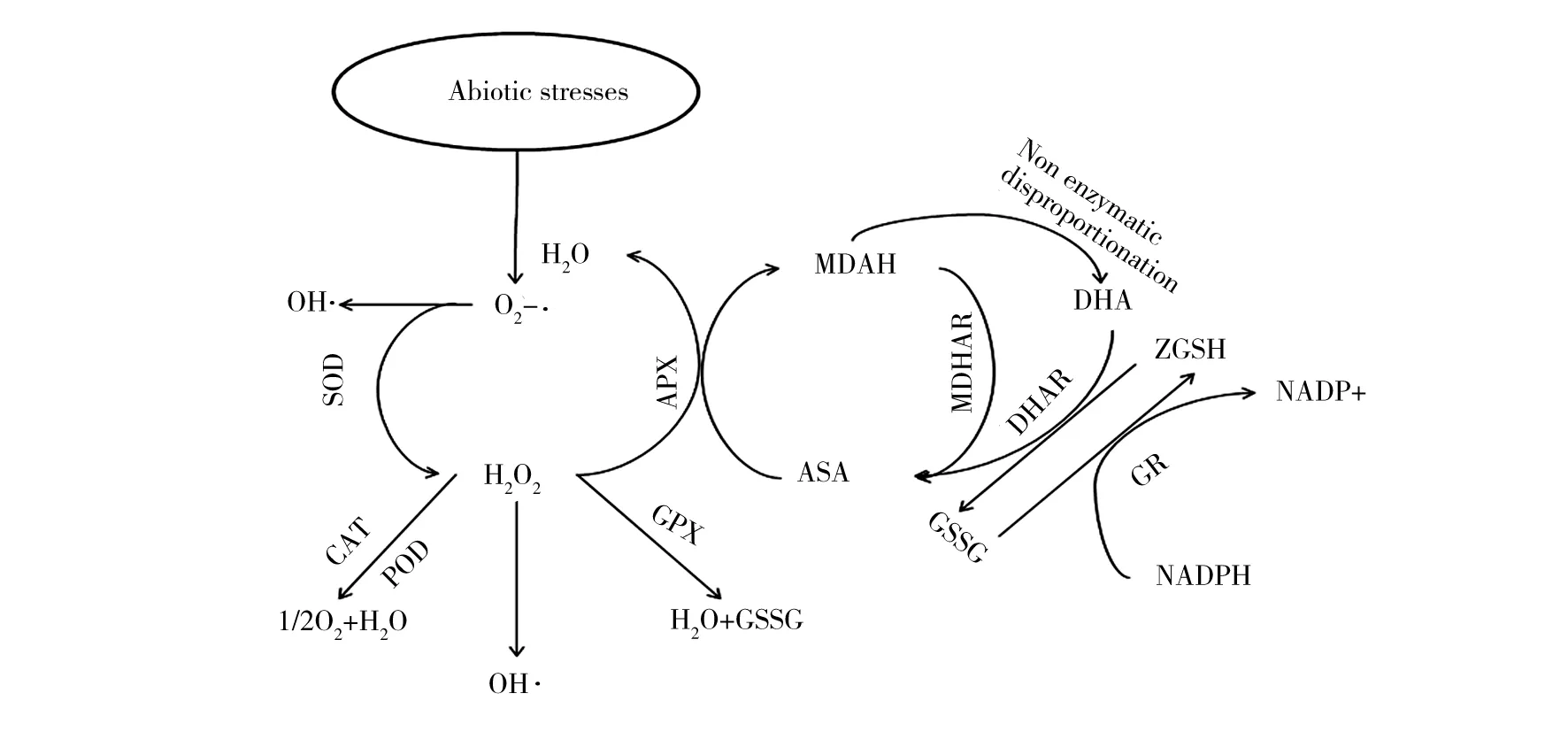
图1 抗氧化系统清除ROS原理图[11]Fig.1 chematic diagram of Antioxidant system to remove ROS[11]
2 食用菌自身因素对于抗氧化酶活性的影响
2.1 不同部位
食用菌子实体不同部位抗氧化酶含量不同,在受到环境胁迫时,不同部位所受到的伤害程度也不一致,从而导致抗氧化酶活性的变化也有所差异。陈明杰等[12]在研究香菇子实体SOD的过程中发现,香菇子实体的不同部位SOD的活性是不一致的,在菌柄和菌盖中SOD的活性基本一致,但是在菌褶中酶活性较高。叶静君等[13]在研究香菇子实体在低温贮藏过程中生理生化变化时发现,菌肉和上柄的维生素C、SOD和CAT的活性高于表皮和下柄,菌肉和上柄组织中O2-·、 H2O2和丙二醛(malondialdehyde,MDA)的含量低于表皮和下柄,菌肉和上柄内酶促和非酶促两类过氧化物防御体系均比表皮和下柄强,抗后熟衰老能力也强,故不易产生褐变现象,这种生理变化趋势与香菇在贮藏过程中表皮和下柄最先出现品质下降的现象相吻合。Conn[14]认为在植物中存在代谢产物区域性分布,即有害的代谢产物分布在特定的器官和组织中,根据上述的分析,在食用菌中也可能存在类似的机制。
2.2 不同的发育阶段
自由基引起的衰老理论在近半个多世纪以来得到了人们的广泛关注,很多研究证实衰老过程中 CAT、SOD、POD 等抗氧化酶的活性均降低,导致自由基代谢产物含量增高,致使线粒体结构损伤,氧化呼吸传递链传递受阻,使自由基的生成量进一步增加,从而加速了衰老进程。董洪新等[15]在研究猪肚菌子实体发育过程中的抗氧化酶活性变化时发现,在猪肚菌子实体生长阶段,CAT 和SOD 活性维持在较高水平,活性氧自由基的水平亦较低,但在衰老阶段,CAT 和SOD 活性明显下降,致使自由基含量提高,并导致细胞膜损伤加剧。陈明杰等[12]研究香菇的原基、菇蕾(柄盖刚分化)、未破菌膜的子实体、菌膜开始张开的子实体、菌膜已张开但未释放孢子的子实体、开伞并已释放孢子的子实体6个不同发育阶段的SOD酶活性的过程中发现在前4个阶段酶活性基本保持一致,第5个阶段酶活明显增加,第6个阶段酶活性又急剧下降。
3 外界环境对于抗氧化酶活性的影响
食用菌在营养生长、生殖生长以及子实体贮存过程中环境因素的剧烈变化,会对其正常的生长发育产生一定的影响,环境因素的剧烈变化可能对食用菌造成氧化胁迫,从而导致体内活性氧含量巨增。但如果环境因素变化在可控范围内,则有利于食用菌正常的生长发育及贮存加工,所以研究环境因素对食用菌抗氧化酶活性的影响是重点之一。
3.1 温度对抗氧化酶活性的影响
不同的食用菌品种有不同的菌丝生长温度和出菇温度,低于或者高于最适的生长温度,都会对食用菌造成低温或高温胁迫,从而给食用菌的生长发育造成不可逆的影响。魏帅等[16]研究温度因子对阿魏菇菌丝抗氧化酶活性的影响时发现在低温诱导下阿魏菇存在着应答反应,随着诱导温度的降低,SOD活性先下降后上升并最终接近于对照水平。CAT活性虽然变化不稳定,但是一直高于对照,阿魏菇凭借较高的SOD和CAT活性使活性氧处于相对较低的水平来抵御低温伤害。王松华等[17]研究表明,在低温处理前期,草菇菌丝可以通过增强SOD、POD和CAT同工酶的表达而提高活性氧的清除能力,进而增强了抗胁迫的能力。黄桂英[18]研究表明,低温胁迫下真姬菇CAT、POD、SOD的活性都在最初的9 h内上升较快,且均高于对照,之后开始逐渐下降。
3.2 金属离子对抗氧化酶活性的影响
食用菌对于不同的金属元素敏感程度不同,某些金属元素是其正常生长发育的必需元素,其耐受能力相对较强,但是对于一些非生长所需的金属离子胁迫,其往往表现出较强的应激反应。食用菌富集重金属的能力较强,远远超过植物,其富集的金属可通过食物链直接进入人体,危害食用者的健康[19-20]。王松华等[21]的研究表明,随着镉离子浓度的升高,CAT、POD、谷胱甘肽还原酶(glutathione reductase,GR)的酶活性先升高后降低,而SOD、脂氧合酶(lipoxidase,LOX)的酶活性却逐渐升高,导致SOD/CAT的比值显著升高,造成H2O2的积累。王松华等[22]的另一个研究表明,用0~116 mmol/L Cd 处理柱状田头菇菌丝7 d 后,抗氧化酶 SOD、POD、CAT 活性均表现出低浓度 Cd促进酶活性上升, 高浓度抑制酶活性,高浓度同时会诱导LOX同工酶的产生。宋鹏等[23]研究了不同浓度的Cu、Zn、Pb、Cd(氯化盐)分别对双孢蘑菇菌丝进行短时间(1 d)和长时间(8 d)诱导,发现SOD在重金属长时间诱导后活性显著增加,Cd诱导作用最显著;经过长时间诱导,低浓度Cu和高浓度Zn能显著增加CAT酶活。何卓晶等[24]的研究表明,Cu对于蛹虫草菌丝体SOD和CAT的酶活性增强效果显著,Cu的胁迫导致ROS的生成,激发抗氧化酶活性,以抵抗胁迫环境的危害,但是Zn对于SOD的活性影响不大,这与在动植物中施加过量的Zn对其SOD酶活性影响不大的现象一致。江枝和等[25]的研究表明,在相同培养料条件下 Se 浓度为 30 mg/kg 处理的大杯香菇子实体内SOD、POD、CAT 和多酚氧化酶(polyphenol oxidase,PPO)活性分别平均比对照提高了 53.05%、89.21%、6.67%和176.58%,与对照间差异达到极显著水平,SOD、POD、PPO、CAT 活性的提高,可增强体内的抗氧化能力,利于保护生物膜的稳定性,提高抗逆能力。
3.3 采后保鲜技术对抗氧化酶活性的影响
食用菌具有较其他园艺产品高的生理活性,具有很强的呼吸强度和超过90%的含水量[26],品质劣变迅速,容易出现失水、褐变、开伞、腐烂等现象[27],而往往认为机体的衰老机制是抗氧化酶活性降低导致活性氧的不断积累从而使细胞膜膜脂氧化程度增强,使之受到损伤的结果。抗氧化酶活性与逆境下机体受损伤的程度关系十分密切,如果提高逆境中食用菌内源保护酶活性,则可使细胞内代谢活跃,伤害减弱,使食用菌在异常情况下仍有较强的抗逆境能力,因此如何有效地进行食用菌采后的保鲜一直是国内外的研究热点。
辐照贮藏的方法主要是利用60Co等射线照射产品,抑制有害微生物的生长,提高抗氧化酶活性,延长贮藏时间,目前辐射保藏的方法已经广泛应用于果蔬的保鲜[28-29]。同样辐射保鲜的方法也可适用于食用菌保鲜,防止食用菌开伞和褐变,延长其贮藏时间。王耀松[30]在研究蟹味菇采后生理及保鲜技术时发现,不同剂量60Co 辐照处理对蟹味菇子实体相关酶活性变化影响不同,0.8 kGy能保持较高的SOD、CAT活性,减小细胞膜的氧化,减轻褐变程度和破膜率,但是辐射剂量过低或者过量都无法达到较好的保鲜效果。叶蕙等[31]用60Co 射线处理草菇发现在 16 ℃下用带孔自发气调包装贮藏0.8 kGy 辐照处理后的草菇,结果能促进贮藏初期的SOD、CAT的酶活性的增加,抑制膜脂过氧化过程,保持了膜结构的完整性,延缓了草菇的衰老进程,延长了贮藏保鲜期。熊巧玲[32]的研究表明1.2 kGy的60Co的γ辐照能有效延长白灵菇的贮藏保鲜时间,通过该剂量辐照的白灵菇,具有较高的SOD和CAT活性以及较低的PPO及蛋白酶活性,软腐较慢。Gill等[33]也提出以 2 kGy的γ射线处理双孢蘑菇,在 10 ℃、11 d 的贮藏期中,开伞及菌柄伸长方面均受到抑制。Barkai等[34]提出以 0.5 kGy 的γ射线辐照可以在 15 ℃下降低双孢蘑菇50%的呼吸率,但此降低的效果在 5 ℃则不会出现。
气调贮藏最早出现在英国,最先应用于果蔬保鲜,是近期发展起来的一项新技术。它主要是在冷藏的基础上通过调节空气组分比例(降低O2浓度,增加 CO2浓度,以 N2作为平衡气体)来抑制呼吸作用,使机体处于缓慢的代谢活动中,从而起到保鲜的作用[35-36]。LI等[37]的研究表明,利用带有气孔的气调包装来贮藏茶树菇,与对照组相比,SOD、CAT酶活性都是先升高后降低,但是在贮存的后半段,实验组表现出更高的抗氧化酶活性和良好的感官品质。YE等[38]的研究表明,在4 ℃低温下利用气调包装贮藏香菇可以延长香菇的贮藏期至17 d,在气调包装中CO2的浓度对于香菇的贮藏品质也有一定的影响,当CO2的浓度为10%时,能够有效抑制香菇的呼吸速率及MDA含量的积累,延迟软化及褐变出现的时间。Tao等[39]的研究表明,将食用菌的子实体真空冷冻后用气调包装贮藏能够诱导SOD、CAT的酶活性的增加,降低贮存过程中MDA含量的积累,加速了活性氧的清除,对延长食用菌子实体贮存时间有积极的意义。
化学保鲜在食用菌釆后贮藏保鲜中也起到很重要的作用,配制一定浓度的化学药液浸泡新鲜子实体,如NaCl、CaCl2,根据不同品种配制成不同浓度的溶液,浸泡或漂洗子实体,可以防止变色、变质,抑制开伞老化,增强保存的时间。林志清等[40]的研究表明,一定浓度的Ca2+处理能够极显著提高香菇采后CAT的活性, 极显著抑制香菇采后POD、PPO和APX的活性。蒋冬花等[41]的研究表明,利用壳聚糖涂抹处理香菇子实体, 能够使SOD活性维持较高的水平,MDA含量显著减少,细胞膜透性变小,显著抑制香菇子实体腐烂、变色和腐败细菌的生长。于新等[42]的研究表明,食品添加剂EDTA-2Na、维生素C、酒石酸、柠檬酸、苹果酸、乳酸等都会一定程度抑制草菇的POD、PPO酶活性,从而减缓草菇贮藏过程中褐变的发生,延长草菇的贮存期。吴靖娜[43]的研究表明,采用魔芋葡苷聚糖、琼脂、海藻酸钠和淀粉4种涂膜剂涂膜鸡腿菇,有助于抑制PPO活力、纤维木质化进程及防止水分的损失,在涂膜剂黏度相同的条件下,海藻酸钠涂膜的保鲜效果最好。
4 展 望
影响食用菌抗氧化酶变化的因素众多,既有食用菌自身的因素,也有外界因素对其的影响。不同部位对于活性氧的耐受力不尽相同,对于活性氧的应答反应也不同。外界的温度、金属离子及不同的食用菌的采后保鲜技术都能够诱导抗氧化酶产生相应的应答刺激,从而维持食用菌正常的生长发育。
近几年来,食用菌的抗氧化研究主要集中在抗氧化酶活性的研究上,利用分子手段使抗氧化酶基因过量表达的报道相对较少,这种研究还主要集中在植物中[44-46]。大量研究发现,提高植物体内抗氧化酶活性和增强抗氧化代谢的水平是提高植物抗逆性的有效途径,通过转基因技术手段将抗氧化相关基因导入植物体内表达,能显著提高转基因植株的部分抗氧化能力和抗逆性[47-48]。抗氧化物的基因操作被认为是培育耐胁迫基因植株富有前途的课题,而这一趋势也将成为今后食用菌抗氧化性的研究热点,也将为食用菌的抗逆性研究提供一条可行的途径。而且,一些重要食用菌基因组测序的完成[49-50],为分子手段转基因方面的研究提供重要的信息支持。一方面,可以将基因构建在强启动子下表达,通过提高基因的表达量提高对氧胁迫的耐受能力,同时也不能忽视对抗氧化基因的调控原件的深入分析及各个抗氧化反应之间的抑制作用及代谢产物的积累对于整体抗氧化响应的影响,SOD的高效表达会促进超氧阴离子转化成H2O2,但是过氧化氢酶基因表达并没有相应的提高,容易造成H2O2的积累,对氧胁迫的能力并不会有显著的提高。另一方面,也可以通过改变体内的信号传导途径或提高抗氧化酶类的代谢水平,来提高抵抗氧化胁迫的能力。
[1] Kreslavak V D, Los D, Allakhverdiev S,et al. Signaling role of reactive oxygen species in plants under stress[J].Russian Journal of Plant Physiology, 2012, 59(2):141-154.
[2] VicenteA, MartinezG,Chaves A, et al. Effect of heattreatment on strawberry fruit damage and oxidative metabolism during storage[J].Postharvest Biology and Technology, 2006, 40(2):116-122.
[3] Quan L J, Zhang B, Shi W, et al. Hydrogen peroxide in plants: a versatile molecule of the reactive oxygen species network[J]. Journal of Integrative Plant Biology, 2008, 50(1):2-18.
[4] 薛洪宝, 常华兰, 焦艳娜, 等. Cd胁迫对发芽玉米抗氧化酶体系的影响分析[J]. 上海环境科学, 2011, 30(4):151-157.
[5] 裴斌, 张光灿, 张淑勇, 等. 土壤干旱胁迫对沙棘叶片光合作用和抗氧化酶活性的影响[J].生态学报,2013, 33(5):1386-1396.
[7] Mittler R. Oxidative stress, antioxidants and stress tolerance[J]. Trends in Plant Science, 2002, 7(9):405-410
[8] 张欣. 超氧化物歧化酶(SOD)及其研究进展[J].内蒙古石油化工, 2010, 16(4):14-15.
[9] 南芝润, 范月仙. 植物过氧化氢酶的研究进展[J]. 安徽农学通报, 2008,14(5):27-29.
[10]Schuller D, Ban N, HuysteevanR.The crystal structure of peanut peroxidase[J]. Structure, 1996, 4(2):311-321.
[11]Gill SS, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stresstolerance in crop plants[J]. Plant Physiology and Biochemistry, 2010, 48(12):909-930.
[12]陈明杰, 潘迎捷, 汪昭月, 等.香菇子实体中超氧化物歧化酶的活性变化初探[J]. 食用菌, 1989, (6):12-13.
[13]叶静君, 励建荣, 韩晓祥,等.香菇低温贮藏中不同部位的生理生化变化[J]. 中国食品学报, 2012,12(3):137-144.
[14]ConnE. Chemical conjugation and compartmentation: Plant adaptation to toxic natural products [C]//Key T L et al. Cellular and molecular biology of plant stress. New York: Alan Press, 1985,351-366.
[15]董洪新, 程显好,朱林,等.猪肚菇子实体发育过程中抗氧化酶活性的变化[J].食品科学, 2010, 31(24):469-472.
[16]魏帅,白羽嘉,张丽,等.温度因子对阿魏菇菌丝抗氧化酶活性的影响[J].保鲜与加工, 2012, 12(4):10-13.
[17]王松华, 张华, 陈庆榆, 等. 低温胁迫对草菇菌丝抗氧化酶表达的影响[J]. 热带作物学报, 2009, 30(5):593-597.
[18]黄桂英. 真姬菇低温胁迫下菌丝体酶活变化及差异蛋白质组学研究[D].福州: 福建农林大学, 2008.
[19]黄擎, 李维, 郭相, 等. 重金属在食用菌中的富集研究进展[J].中国食用菌, 2014, 33(2):4-6.
[20]Krupa P, Kozdroj J. Accumulation of heavy metals by ectomycorrhizal fungi colonizing birch trees growing in an industrial desert soil[J]. World Journal of Microbiology & Biotechnology, 2004, 20(4):427-430.
[21]王松华, 张华, 崔元戎,等.镉对灵芝菌丝抗氧化系统的影响[J].应用生态学报, 2008, 19(6):1355-1361.
[22]王松华, 张华, 傅曼琴,等.柱状田头菇菌丝对镉胁迫的抗氧化响应[J].应用生态学报,2007,18(8):1813-1818.
[23]宋鹏, 唐敦义, 邓家玉,等.重金属对双孢菇菌丝合成巯基化合物和抗氧化酶的影响[J]. 四川大学学报(自然科学版), 2010, 41(6):1407-1412.
[24]何卓晶, 王尊生, 李素霞,等.Cu2+、Zn2+胁迫对蛹虫草抗氧化酶活性及脂氧化水平的影响[J]. 药物生物技术,2006,13(1):36-39.
[25]江枝和, 翁伯琪, 雷锦桂,等.添加外源硒对大杯香菇子实体的保护酶系及膜质过氧化酶的影响[J].菌物学报, 2009, 28(4):612-615.
[26]Leixuri A, JesusMF, Catherine BR, et al. Modellingbrowning and brown spotting of mushroom(Agaricusbisporus)stored in controlled environmental conditions using image analysis[J].Journal of Food Engineering, 2009, 91(2):280-286.
[27]Guillaume C, Schwab I, Gastaldi E, et al.Biobased packaging for improving preservation of fresh common mushrooms (AgaricusbisporusL.)[J].Innovative Food Science & Emerging Technologies, 2010, 11(4): 690-696.
[28]Afify A.E.M, BeltagiHse, Aly AA, et al.Antioxidant enzyme activities and lipid peroxidation as biomarker compounds for potato tuber stored by gamma radiation[J].Asian Pacific Journal of Tropical Biomedicine, 2012, 2(3):S1548-S1555.
[29]Hameed A, Shah TM, Atta BM, et al. Gamma irradiation effects on seed germination and growth, protein content, peroxidase and protease activity, lipid peroxidation in Desi and Kabuli Chickpea[J]. Pakistan Journal of Botany, 2008, 40(3):1033-1041.
[30]王耀松.蟹味菇采后生理及贮藏保鲜技术研究[D].杨凌: 西北农林科技大学, 2007.
[31]叶蕙, 陈建勋, 余让才,等.γ辐照对草菇保鲜及其生理机制的研究[J].核农学报, 2000, 14(1):24-28.
[32]熊巧玲. 白灵菇采后生理生化及保鲜技术研究[D].武汉: 华中农业大学, 2007.
[33]Gill WJ, Nichol RC, MARKAKIS P. Irradiation of cultured mushrooms[J].Food Technology, 1969, 23(2):385-388.
[34]Barkai G,Padora B.Mushroom. In Postharvest Biotechnology of Vegetables[M]. USA:CRC Press, 1984, 147-160.
[35]Li TH, Zhang M, Wang SJ. Effects of temperature onAgrocybechaxinguqualitystored in modifiedatmosphere packages with silicon gum film windows[J].LWT-Food Science and Technology, 2008, 41(6):965-973.
[36]Stewart OJ, RaghavanAGSV, GOLDEN KD, et al.MA storage of Cavendish bananas using silicone membrane anddiffusion channel systems[J].Postharvest Biology and Technology, 2005, 35(3):309-317.
[37]Li TH, Zhang M. Effects of modified atmosphere package (MAP) with a silicon gumfilm window and storage temperature on the quality and antioxidant system of storedAgrocybechaxingu[J].LWT-Food Science and Technology, 2010, 43(7):1113-1120.
[38]Ye JJ, Li JR, Han XX, et al.Effects of Active Modified Atmosphere Packaging on Postharvest Quality of Shiitake Mushrooms (Lentinulaedodes) Stored at Cold Storage[J].Journal of Integrative Agriculture, 2012, 11(3):474-482.
[39]Tao F, ZhangM. Effect of vacuum cooling on physiological changes in the antioxidant system of mushroom under different storage conditions[J].Journal of Food Engineering,2007, 79(4):1302-1309.
[40]林志清, 李金雨, 黄维南,等.钙处理对香菇采后氧化酶活性的影响[J].福建农业学报, 2001, 16(3):56-60.
[41]蒋冬花, 许朝渊, 张萍华,等.3种保鲜剂对香菇保鲜效果[J]. 食品科学, 2004, 25(9):194-197.
[42]于新, 冯彤, 李远志,等.食品添加剂对草菇PPO和POD活性的影响[J]. 食品工业科技, 2002, 23(5):10-13.
[43]吴靖娜. 鸡腿菇保鲜机理及保鲜技术研究[D].福州: 福建农林大学, 2008.
[44]Breusegem FV, SlootenL L, Stassart JM, et al.Effects of overproduction of tobacco MnSOD in maize chloroplasts on foliar tolerance to cold and oxidativestress[J].Journal of Experimental Botany, 1999, 50(330): 71-78.
[45]Du J, Zhu Z, Li WC. Over-expression of exotic superoxide dismutase gene MnSOD and increase in stress resistance in maize[J]. Journal of Plant Physiology, 2006, 32(1):57-63.
[46]王凤德, 衣艳君, 王海庆,等. 豌豆过氧化氢酶在烟草叶绿体中的过量表达提高了植物的抗逆性[J]. 生态学报, 2011, 31(4):1058-1063.
[47]MittlerR, Zilinakes BA. Purification and characterization of pea cytosolic ascorbateperoxidase[J].Plant Physiology, 1991, 97(3):962-968.
[48]Mittler R, ZilinakesBA.Regulation of pea cytosolic ascorbate peroxidase and other antioxidant enzymes during the progression of drought stress and following recovery from drought [J].Plant Journal, 1994, 5(3):397-405.
[49]Morin E, Kohler A, Baker AR, et al. Genome sequence of the button mushroomAgaricusbisporusreveals mechanisms governing adaption to a humic-rich ecological niche[J].Proceedings of the National Academy of Sciences, 2012, 109(43):17501-17506.
[50]Chen S, Xu J, Liu C, et al.Genome sequence of the model medicinal mushroomGanodermalucidum[J/OL].Nat Commun, 2012, 3:913.DOI:10.1038/ncomms1923.
Advance in Effective Factors of Antioxidant Enzyme Activity in Edible Fungi
CUI Shi-rui1, 2, FENG Zhi-yong1, CHEN Ming-jie1, WANG Hong1, CHEN Hui1
(1.Inst.ofEdibleFungiSAAS,NationalEngin.Res.Ctr.ofEdibleFungi,Shanghai210403; 2.Coll.OfLifeSci. ,NangjingAgric.Uni.,Nanjing210095)
The mycelia and sporogonia of edible fungi can produce active oxygen (AO) during normal growth and development phases. But the AO will increase dramatically when facing with the environmental stress or during storage and transportation, in this way the balance of AO is broken. Antioxidant system then must be activated against its harmful effect. The categories of antioxidant enzymes and their effect were introduced in this article. The own factor such as different parts, different development phases and the external environment factors including temperature, metal ion, and postharvest fresh-keeping technology which influence the activity of antioxidant enzymes were also reviewed, and the research prospects of edible fungi antioxidant enzyme was discussed.
edible fungi; antioxidant enzymes; activity change
国家自然科学基金项目(31401932)
崔世瑞 女,硕士研究生。研究方向为食用菌的遗传与栽培。E-mail:sdcuishirui@126.com
* 通讯作者。男,助理研究员。研究方向为食用菌的遗传与栽培。Tel:021-62200747,E-mail:huichen_js@aliyun.com
2014-09-16;
2014-10-12
Q935
A
1005-7021(2015)04-0087-06
10.3969/j.issn.1005-7021.2015.04.015
猜你喜欢
今日农业(2020年16期)2020-12-14
海峡姐妹(2019年6期)2019-06-26
海峡姐妹(2018年8期)2018-09-08
知识经济·中国直销(2018年6期)2018-06-29
物理化学学报(2017年8期)2017-12-19
现代检验医学杂志(2016年1期)2016-11-12
医学研究杂志(2015年4期)2015-06-10
中华皮肤科杂志(2014年3期)2014-12-19
红蜻蜓·低年级(2014年3期)2014-07-09
