
重组乳酸克鲁维酵母产磷脂酶A2发酵条件的优化
2015-12-26 09:43李由然顾正华丁重阳石贵阳陈国安杨盛荣
食品科学 2015年21期
王 辉,张 梁,*,李由然,顾正华,丁重阳,石贵阳,陈国安,杨盛荣
(1.江南大学 工业生物技术教育部重点实验室,江苏 无锡 214122;2.江南大学 粮食发酵工艺与技术国家工程实验室,江苏 无锡 214122;3.无锡江大百泰科技有限公司,江苏 无锡 214122)
重组乳酸克鲁维酵母产磷脂酶A2发酵条件的优化
王 辉1,2,张 梁1,2,*,李由然2,顾正华2,丁重阳2,石贵阳2,陈国安3,杨盛荣3
(1.江南大学 工业生物技术教育部重点实验室,江苏 无锡 214122;2.江南大学 粮食发酵工艺与技术国家工程实验室,江苏 无锡 214122;3.无锡江大百泰科技有限公司,江苏 无锡 214122)
利用重组乳酸克鲁维酵母(Kluyveromyces lactis)GG799表达磷脂酶A2,对其产酶发酵条件进行研究。采用单因素试验和正交试验对培养基及培养条件进行优化,确定了重组菌产酶的最佳发酵条件。结果表明:最优培养基组成为葡萄糖30 g/L、酵母粉20 g/L、蛋白胨30 g/L、Kh2PO43 g/L;最优培养条件为:发酵温度30 ℃、接种量2%(V/V)、初始ph 7.0、装液量90 mL/250 mL三角瓶、摇床转速220 r/min,在此条件下发酵培养,酶活力由(1.87±0.12)U提高到(5.35±0.27)U。
乳酸克鲁维酵母;磷脂酶A2;发酵
磷脂酶A2(phospholipase A2,PLA2,EC 3.1.1.4)广泛存在于各种生物组织中,它可催化磷脂甘油分子二位酰基水解生成溶血卵磷脂和游离脂肪酸,而溶血卵磷脂比磷脂具有更高的亲水性[1-2]。磷脂酶A2可以广泛应用于食品、化妆品及医药等领域中,如面包制作、蛋黄改性以及植物油脱胶等[3-4]。早期,磷脂酶A2可以从一些哺乳动物的胰腺组织中提取,但是来源少,提取过程复杂且成本较高[5]。
目前,对于磷脂酶A2的研究主要集中在哺乳动物磷脂酶A2基因(pla2)的重组表达方面[6-7],但存在易形成包涵体,表达量低以及宿主无法完成复杂的哺乳动物重组蛋白后加工等问题[3,8-10],同时,宿主菌的遗传背景是否安全也影响着重组磷脂酶A2在食品等领域的应用。另外,据报道,由于磷脂酶A2对磷脂的水解作用,因此,可能会对大肠杆菌等宿主菌有毒害作用[6,11]。乳酸克鲁维酵母(Kluyveromyces lactis)是美国食品药品管理局(Food and Drug Administration,FDA)认定为GRAS(generally regarded as safe)的微生物,具有良好的外源蛋白分泌性能以及高密度发酵的特点,相对于其他表达宿主菌,对于外源蛋白的重组表达具有很大的优势[12]。近年来,乳酸克鲁维酵母在食品领域已成功用于凝乳酶的重组表达[13],在医药领域也已成功实现了人血清白蛋白、白细胞介素的重组表达[14-15],因此,乳酸克鲁维酵母在实现外源蛋白的表达及应用等方面具有巨大的潜力。
本实验室前期已成功实现了原核微生物Lactobacillus casei DSM20011 pla2基因在乳酸克鲁维酵母GG799中的分泌表达。因此,本实验在此基础上对乳酸克鲁维酵母重组表达磷脂酶A2的发酵条件进行优化研究,以提高重组酶产量及活性,降低生产成本,为工业化生产提供一定的理论依据。
1 材料与方法
1.1 材料与试剂
1.1.1 菌株
乳酸克鲁维酵母(Kluyveromyces lactis)重组菌株GG799/pKLAC1-pla2由粮食发酵工艺与技术国家工程实验室前期构建并保藏。
1.1.2 培养基
固体培养基组成:葡萄糖20 g/L、蛋白胨20 g/L、酵母粉10 g/L、琼脂20 g/L;种子液培养基组成:葡萄糖20 g/L、蛋白胨20 g/L、酵母粉10 g/L,ph 7.0;发酵培养基组成:葡萄糖30 g/L、酵母粉20 g/L、蛋白胨30 g/L、Kh2PO43 g/L,ph 7.0。
1.1.3 试剂
卵磷脂 Acros公司;其余试剂均为分析纯。
1.2 仪器与设备
hyL-C组合式摇床 太仓市强乐实验设备有限公司;生化培养箱 上海一恒科技有限公司;电子滴定仪上海羽通仪器仪表厂;水浴恒温振荡器 太仓市医疗器械厂;可见分光光度计 上海美谱达仪器有限公司。
1.3 方法
1.3.1 菌种活化
从甘油保藏管取一定量的菌液,在固体培养基上划线转接,置于30 ℃恒温培养箱培养3 d后,置于4 ℃冰箱中保存备用。
1.3.2 种子液制备
将活化的菌种接入装有50 mL种子培养基的250 mL三角瓶中,在振荡培养箱中30 ℃、250 r/min培养22 h,作为种子液。
1.3.3 菌体生物量测定
生物量以菌体干质量表示,取发酵液于12 000 r/min离心5 min,弃上清液,无菌水重新悬浮菌体,反复3 次,菌体75 ℃烘箱烘至恒质量,称质量。
1.3.4 酶活力的测定
酶活力的测定[16]:采用酸碱滴定法测定磷脂酶A2酶活力,取1 mL酶液加入9 mL用Tris-HCl(50 mmol/L,ph 8.0)配制的含质量分数4%的卵磷脂、体积分数1%的Triton X-100、30 mmol/L CaCl2的反应体系中,37 ℃、200 r/min准确反应15 min,反应完成后加入10 mL无水乙醇终止反应,生成的游离脂肪酸用20 mmol/L的NaOh滴定。酶活力单位(U)定义:1 mL酶液1 min催化卵磷脂生成1 μmol游离脂肪酸定义为一个酶活力单位。
1.3.5 单因素试验
1.3.5.1 不同碳源对菌体生长及产酶的影响
在初始发酵培养基的基础上,分别以乳糖、半乳糖、蔗糖、葡萄糖、甘油、麦芽糖和糊精为唯一碳源,添加量均为20 g/L,以酵母粉和蛋白胨为氮源,添加量分别为10 g/L和20 g/L,于ph 7.0、30 ℃、200 r/min摇瓶培养72 h,测定菌体生物量及发酵上清磷脂酶A2酶活性。
1.3.5.2 碳源添加量对菌体生长及产酶的影响
以酵母粉、蛋白胨为氮源,添加量分别为10 g/L和20 g/L,葡萄糖为碳源,添加量分别为10、15、20、25、30、40 g/L,于ph 7.0、30 ℃、200 r/min摇瓶培养72 h,测定菌体生物量及发酵上清磷脂酶A2酶活性。
1.3.5.3 不同氮源对菌体生长及产酶的影响
以葡萄糖为碳源,添加量为20 g/L ,分别以鱼粉蛋白胨、酵母粉、蛋白胨、蛋白胨+酵母粉(2∶1,m/m)、牛肉浸膏、硝酸钠和氯化铵为氮源,添加量为30 g/L,于ph 7.0、30 ℃、200 r/min摇瓶培养72 h,测定菌体生物量及发酵上清磷脂酶A2酶活性。
1.3.5.4 氮源质量配比对菌体生长及产酶的影响
以葡萄糖为碳源,添加量为20 g/L,以酵母粉和蛋白胨为复合氮源,添加量为30 g/L,添加质量比例分别为1∶1、1∶2、1∶3、1∶4、2∶1、2∶3,于ph 7.0、30 ℃、200 r/min摇瓶培养72 h,测定菌体生物量及发酵上清磷脂酶A2酶活性。
1.3.5.5 无机盐对菌体生长及产酶的影响
以酵母粉和蛋白胨为复合氮源,添加量为30 g/L,添加质量比例为1∶2,分别以Kh2PO4、MgCl2、MnCl2、ZnCl2、FeCl2、NaCl、NiCl2和CaCl2为唯一无机盐,于ph 7.0、30 ℃、200 r/min摇瓶培养72 h,测定菌体生物量及发酵上清磷脂酶A2酶活性。
1.3.5.6 Kh2PO4添加量对菌体生长及产酶的影响
以Kh2PO4为无机盐,添加量分别为0.1、1.0、3.0、5.0、8.0、10.0 g/L,于ph 7.0、30 ℃、200 r/min摇瓶培养72 h,测定菌体生物量及发酵上清磷脂酶A2酶活性。
1.3.5.7 培养温度对菌体生长及产酶的影响
在已优化的培养基成分基础上,250 mL摇瓶装50 mL培养基,培养温度分别为24、27、30、33 ℃,于ph 7.0、200 r/min摇瓶培养72 h,测定菌体生物量及发酵上清磷脂酶A2酶活性。
1.3.5.8 接种量对菌体生长及产酶的影响
培养温度30 ℃,接种量(V/V)分别为1%、2%、3%、4%、5%,于ph 7.0、200 r/min摇瓶培养72 h,测定菌体生物量及发酵上清磷脂酶A2酶活性。
1.3.5.9 初始ph值对菌体生长及产酶的影响
接种量2%,调节培养基初始ph值分别为5.0、5.5、6.0、6.5、7.0、8.0,于30 ℃、200 r/min摇瓶培养72 h,测定菌体生物量及发酵上清磷脂酶A2酶活性。
1.3.5.10 装液量对菌体生长及产酶的影响
调整250 mL摇瓶中培养基装液量分别为40、50、60、70、80 mL,于ph 7.0、30 ℃、200 r/min摇瓶培养72 h,测定菌体生物量及发酵上清磷脂酶A2酶活性。
1.3.5.11 摇床转速对菌体生长及产酶的影响
装液量为70 mL,调节摇床转速分别为140、160、180、200、220、240 r/min,于ph 7.0、30 ℃、摇瓶培养72 h,测定菌体生物量及发酵上清磷脂酶A2酶活性。
1.3.6 正交试验设计
在单因素试验的基础上,以葡萄糖添加量、酵母粉添加量、蛋白胨添加量、Kh2PO4添加量、接种量和装液量为自变量,以PLA2酶活力为指标,进行L18(37)正交试验,其中设置一个空列项。
2 结果与分析
2.1 单因素试验结果
2.1.1 不同碳源对菌体生长及产酶的影响

图1 不同碳源对重组菌生长及产酶的影响Fig.1 Effect of different carbon sources on cell growth and PLA2activity
由图1可知,不同碳源对重组菌的产酶和生长影响较大,糊精不利于重组酵母的生长和产酶,这主要与其作为碳源较难被酵母吸收利用有关;当碳源为蔗糖、甘油和麦芽糖时重组酵母虽然生长较好,但产酶能力较乳糖、半乳糖和葡萄糖为碳源时低,这主要是因为碳源不仅会影响乳酸克鲁维酵母的生长代谢,而且会影响该过程蛋白质的分泌,适宜的碳源有利于重组菌外源蛋白的表达[17]。因为葡萄糖为碳源时酶活性最高,故选取葡萄糖为发酵培养基中的碳源。
2.1.2 碳源添加量对菌体生长及产酶的影响

图2 碳源质量浓度对重组菌生长及产酶的影响Fig.2 Effect of carbon source concentration on cell growth and PLA2activity
由图2可知,当碳源添加量为10 g/L时,不利于菌体的生长繁殖,菌体量及产酶量均很低。随着碳源质量浓度的增加,菌体量及产酶量也随之增加,但当碳源质量浓度高于25 g/L时,重组菌的产酶量有所下降,这可能是因为较高质量浓度的碳源对微生物生长有抑制作用,延长发酵周期,不利于发酵产物的积累[18]。当碳源质量浓度为20 g/L时,重组菌产酶量最大,故选取发酵培养基中碳源的质量浓度为20 g/L。
2.1.3 不同氮源对菌体生长及产酶的影响

图3 不同氮源对重组菌生长及产酶的影响Fig.3 Effect of different nitrogen sources on cell growth and PLA2activity
由图3可知,有机氮源对菌体的生长和产酶都较为有利,而重组菌对无机氮源的利用较差,复合氮源条件下,菌株生长和产酶均高于单一氮源,这主要是因为氮源能够影响酵母重组蛋白的表达,而且酵母粉可以提高外源蛋白的分泌量和积累量[19],当蛋白胨和酵母粉同时作为氮源时酶活性最高,因此,选取蛋白胨和酵母粉复合氮源作为发酵培养基的氮源。
2.1.4 复合氮源质量配比对菌体生长及产酶的影响

图4 氮源质量配比对重组菌生长及产酶的影响Fig.4 Effect of nitrogen source concentration on cell growth and PLA2activity
由图4可知,复合氮源酵母粉和蛋白胨以不同的质量比例加入发酵培养基中时,对菌体的生长量影响较小,当m(酵母粉)∶m(蛋白胨)=2∶3时最不利于重组菌产酶,这可能是因为不同的氮源比例可以通过菌体代谢影响发酵培养基的ph值,从而影响代谢产物的形成[20]。当m(酵母粉)∶m(蛋白胨)=1∶2时,重组菌产酶量最高,故选取酵母粉和蛋白胨的添加比例为1∶2。
2.1.5 无机盐对菌体生长及产酶的影响

图5 无机盐对重组菌生长及产酶的影响Fig.5 Effect of different mineral salts on cell growth and PLA2activity
由图5可知,ZnCl2和FeCl2的加入有利于菌体的生长,而Kh2PO4的加入对菌体的生长影响不大,但却对重组菌产酶能力有促进作用,这可能是因为无机盐如MgCl2、Kh2PO4等主要参与一些酶的组成成分,对维持酶的活性等有重要作用[21]。因此,选取Kh2PO4作为发酵培养基中的无机盐。
2.1.6 无机盐添加量对菌体生长及产酶的影响
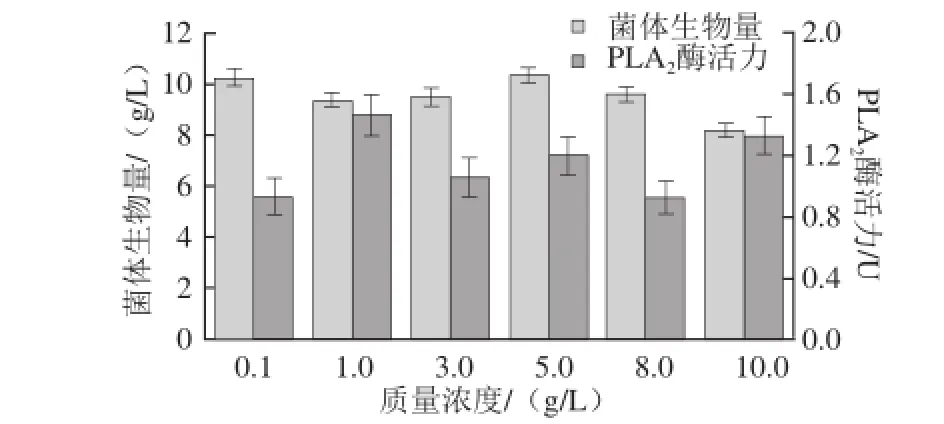
图6 无机盐添加量对重组菌生长及产酶的影响Fig.6 Effect of mineral salt concentration on cell growth and PLA2activity
由图6可知,当Kh2PO4的添加量为5.0 g/L时,菌体生物量最高,说明该添加量下有利于菌体的生长,而添加量为1.0 g/L时,酶活力最高,故发酵培养基中无机盐的添加量为1.0 g/L。
2.1.7 接种量对菌体生长及产酶的影响
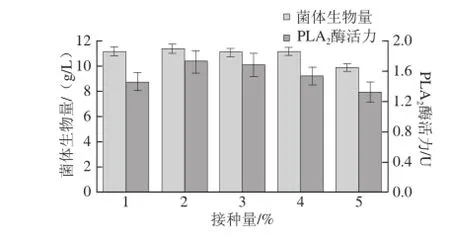
图7 接种量对重组菌生长及产酶的影响Fig.7 Effect of inoculum amount on cell growth and PLA2activity
由图7可知,接种量为1%时,不利于重组菌的产酶,但当接种量达到3%及以上时,重组菌产酶能力下降,这是因为接种量可以影响微生物生长的延迟期,接种量过小,菌体不足,延迟期较长;接种量过大,菌体繁殖快,由于培养基营养物质消耗过快,不利于微生物的持续生长,并进而影响产物的合成[22]。接种量为2%时,酶活力最高,选取发酵培养基的接种量为2%。
2.1.8 培养温度对菌体生长及产酶的影响
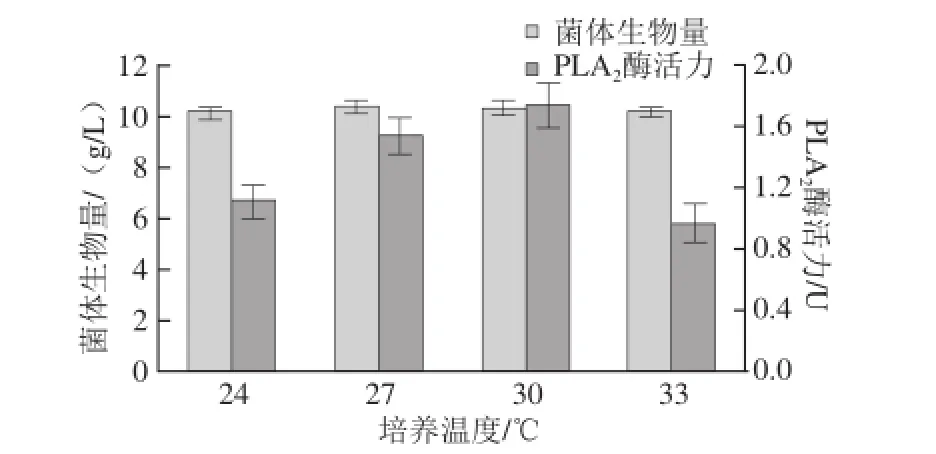
图8 培养温度对重组菌生长及产酶的影响Fig.8 Effect of cultivation temperature on cell growth and PLA2activity
由图8可知,在所选温度条件下,重组菌的生长量基本不受影响,但在24、33 ℃条件下酶活性较低,这可能是因为温度过低会减缓菌株的生长代谢,而过高会影响重组酶的稳定性以及后续的加工过程[23],这些均不利于重组菌产酶。当温度为30 ℃时,酶活力最高,故选取的发酵培养温度为30 ℃。
2.1.9 初始ph值对菌体生长及产酶的影响

图9 初始pH值对重组菌生长及产酶的影响Fig.9 Effect of initial pH on cell growth and PLA2activity
由图9可知,重组菌对ph值的耐受范围较广,在所选ph值范围内菌株生长基本不受影响,当培养基初始ph值为8时,酶活性较其他ph值条件下的低,这可能是因为在酵母发酵培养过程中,不同ph值对其分泌的蛋白酶活性影响不同,培养基中蛋白酶活性高时,可能会对重组酶产生降解,进而影响其活性[24]。ph值为7.0时,有利于产酶,此条件下酶活性最高,因此,调节发酵培养基初始ph值为7.0。
2.1.10 装液量对菌体生长及产酶的影响
由图10可知,装液量对菌株生长及产酶均有影响,且对产酶影响较大。当装液量为40 mL/250 mL摇瓶时,不利于重组菌产酶,随着装液量的增加,酶活力逐渐提高,而装液量增加到80 mL/250 mL摇瓶时,酶活力降低,这可能是因为装液量影响酵母发酵过程的溶氧,从而影响菌体对营养物质的吸收利用以及代谢产物的形成[25]。当装液量为70 mL/250 mL摇瓶时,酶活力达到最高。因此,选取摇瓶装液量为70 mL/250 mL摇瓶。

图10 装液量对重组菌生长及产酶的影响Fig.10 Effect of medium volume on cell growth and PLA2activity
2.1.11 摇床转速对菌体生长及产酶的影响
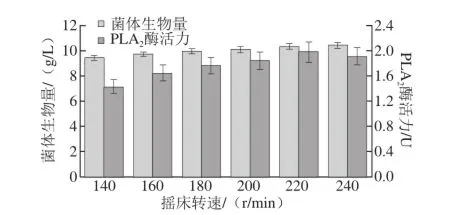
图11 摇床转速对重组菌生长及产酶的影响Fig.11 Effect of shaking speed on cell growth and PLA2activity
由图11可知,当摇床转速为140 r/min时,酶活力较低,随着摇床转速的提高,菌体生物量以及酶活力均逐渐增加,这可能是因为当装液量一定时,转速过低,发酵过程溶氧不足,影响菌体生长以及发酵周期[26],而当摇床转速达到240 r/min时酶活性有所下降,这可能是转速过快加快了发酵过程中营养物质的挥发损失,从而影响了重组菌的产酶。当转速达到220 r/min时,重组菌酶活力最高,故选取摇床转速为220 r/min。
2.2 正交试验分析
根据正交设计原理以及发酵培养基和发酵条件的单因素试验结果,采用L18(37)进行六因素三水平的正交试验,试验结果与方差分析见表1、2。
由表1极差分析可知,各因素对重组菌发酵酶活力的影响大小为:Kh2PO4添加量>葡萄糖添加量>装液量>酵母粉添加量>接种量>蛋白胨添加量,比较每个因素的3 个不同水平,并考虑实际的取值情况,得到最佳组合为A2B3C3D3E2F3。
由表2方差分析可知,葡萄糖及Kh2PO4对重组菌产酶影响显著,因此,由正交试验得到了最佳发酵条件,即各因素添加量为葡萄糖30 g/L、酵母粉20 g/L、蛋白胨30 g/L、Kh2PO43 g/L、接种量2%以及装液量90 mL/ 250 mL摇瓶。在该条件下进行验证实验,重组酶酶活力达到(5.35±0.27) U。

表1 正交试验设计方案及结果Table 1 Orthogonal array design with experimental results

表2 正交试验结果方差分析Table 2 Analysis of variance of the experimental results of orthogonal array design
3 结 论
对重组菌K. lactis GG799/pKLAC1-pla2产磷脂酶A2培养基和培养条件进行了优化。得到的最佳产酶发酵条件为:葡萄糖30 g/L、酵母粉20 g/L、蛋白胨30 g/L、Kh2PO43 g/L、培养温度30 ℃、初始ph 7.0、接种量2%(V/V)、装液量90 mL/250 mL摇瓶、转速220 r/min。优化后酶活力由(1.87±0.12) U提高到(5.35±0.27) U,高于已报道的利用大肠杆菌和曲霉表达系统得到的PLA2酶活性[8,27],为进一步的规模化生产提供理论依据。
参考文献:
[1] BURKE J E, DENNIS E A. Phospholipase A2biochemistry[J]. Cardiovascular Drugs and Therapy, 2008, 23(1): 49-59.
[2] 李永端, 谷克仁. 溶血磷脂的性能与应用[J]. 粮油加工, 2007(4): 55-57.
[3] de MARIA L, VIND J, OXENBOLL K M, et al. Phospholipases and their industrial applications[J]. Applied Microbiology and Biotechnology, 2007, 74(2): 290-300.
[4] 周家春. 溶血磷脂开发应用[J]. 粮食与油脂, 2002(3): 33-34.
[5] 梁丽, 常明, 刘睿杰, 等. 磷脂酶研究进展[J]. 食品工业科技, 2013, 34(4): 393-396.
[6] SETO M, OGAWA T, KODAMA K, et al. A novel recombinant system for functional expression of myonecrotic snake phospholipase A2in Escherichia coli using a new fusion affinity tag[J]. Protein Expression and Purification, 2008, 58(2): 194-202.
[7] 邓晨旭, 江连洲, 陈璐璐, 等. 猪磷脂酶A2基因在黑曲霉中的表达[J].生物技术, 2014, 24(3): 35-39.
[8] JOVEL S R, KUMAGAI T, DANSHIITSOODOL N, et al. Purification and characterization of the second Streptomyces phospholipase A2refolded from an inclusion body[J]. Protein Expression and Purification, 2006, 50(1): 82-88.
[9] 李维琳. 紫红链霉菌2917磷脂酶A2基因的克隆及序列分析[J]. 武汉大学学报: 理学版, 2002, 48(4): 500-506.
[10] GELLISSEN G, HOLLENBERG C P. Application of yeasts in gene expression studies: a comparison of Saccharomyces cerevisiae, Hansenula polymorpha and Kluyveromyces lactis-a review[J]. Gene, 1997, 190(1): 87-97.
[11] HONG S, HORIUCHI H, OHTA A. Identification and molecular cloning of a gene encoding phospholipase A2(plaA) from Aspergillus nidulans[J]. Biochimica et Biophysica Acta, 2005, 1735(3): 222-229.
[12] van OOYEN A J, DEKKER P, HUANG M, et al. Heterologous protein production in the yeast Kluyveromyces lactis[J]. FEMS yeast Research, 2006, 6(3): 381-392.
[13] van DEN BERG J A, van der LAKEN K J, van OOYEN A J, et al. Kluyveromyces as a host for heterologous gene expression: expression and secretion of prochymosin[J]. Nature Biotechnology, 1990, 8(2): 135-139.
[14] FLEER R, CHEN X, AMELLAL N, et al. High-level secretion of correctly processed recombinant human interleukin-1 beta in Kluyveromyces lactis[J]. Gene, 1991, 107(2): 285-295.
[15] LODI T, NEGLIA B, DONNINI C. Secretion of human serum albumin by Kluyveromyces lactis overexpressing KlPDI1 and KlERO1[J]. Applied and Environment Microbiology, 2005, 71(8): 4359-4363.
[16] YANG Weili, PENG Lisheng, ZHONG Xiaofen, et al. Functional expression and characterization of a recombinant phospholipase A2from sea snake Lapemis hardwickii as a soluble protein in E. coli[J]. Toxicon, 2003, 41(6): 713-721.
[17] MADINGER C L, SHARMA S S, ANTON B P, et al. The effect of carbon source on the secretome of Kluyveromyces lactis[J]. Proteomics, 2009, 9(20): 4744-4754.
[18] 缪静, 杨在东, 冯志彬, 等. 碳源对γ-聚谷氨酸发酵的影响[J]. 中国酿造, 2010, 29(3): 70-72.
[19] SREEKRISHNA K, BRANKAMP R G, KROPP K E, et al. Strategies for optimal synthesis and secretion of heterologous proteins in the methylotrophic yeast Pichia pastoris[J]. Gene, 1997, 190(1): 55-62.
[20] MACAULEY-PATRICK S, FAZENDA M L, MENEIL B, et al. Heterologous protein production using the Pichia pastoris expression system[J]. Yeast, 2005, 22(4): 249-270.
[21] 吕欣, 段作营, 毛忠贵. 氮源与无机盐对高浓度酒精发酵的影响[J].西北农林科技大学学报: 自然科学版, 2003, 31(4): 159-162.
[22] 朱明军, 梁世中. 装液量和接种量对红发夫酵母生长和虾青素积累的影响[J]. 氨基酸和生物资源, 2002, 24(4): 28-31.
[23] HONG F, MEINANDER N Q, JÖNSSON L J. Fermentation strategies for improved heterologous expression of laccase in Pichia pastoris[J]. Biotechnology and Bioengineering, 2002, 79(4): 438-449.
[24] FLORES M V, CUELLAS A, VOGET C E. The proteolytic system of the yeast Kluyveromyces lactis[J]. Yeast, 1999, 15(14): 1437-1448.
[25] MERICO A, CAPITANIO D, VIGENTINI I, et al. How physiological and cultural conditions influence heterologous protein production in Kluyveromyces lactis[J]. Journal of Biotechnology, 2004, 109(1/2): 139-146.
[26] 张建勇, 王晓港, 王水莲, 等. 溶氧对重组毕赤酵母高密度发酵生产腺苷蛋氨酸的影响[J]. 齐鲁药事, 2007, 26(7): 428-430.
[27] SUGIYAMA M, OHTANI K, IZUHARA M, et al. A novel prokaryotic phospholipase A2. Characterization, gene cloning, and solution structure[J]. Journal of Biological Chemistry, 2002, 277(22): 20051-20058.
Optimization of Fermentation Conditions for the Production of Phospholipase A2by Recombinant Kluyveromyces lactis
wANG hui1,2, ZHANG Liang1,2,*, LI Youran2, GU Zhenghua2, DING Zhongyang2, SHI Guiyang2, ChEN Guoan3, YANG Shengrong3
(1. Key Laboratory of Industrial Biotechnology, Ministry of Education, Jiangnan University, Wuxi 214122, China; 2. National Engineering Laboratory for Cereal Fermentation Technology, Jiangnan University, Wuxi 214122, China; 3. Wuxi Jiangda Baitai Technology Co. Ltd., Wuxi 214122, China)
Fermentation conditions for the production of phospholipase A2(PLA2) by recombinant Kluyveromyces lactis GG799 were optimized using single factor and orthogonal design experiments. The optimal fermentation medium was composed of 30 g/L glucose, 20 g/L yeast extract, 30 g/L peptone and 3 g/L KH2PO4. The best fermentation conditions were determined as 30 ℃,2%, 7.0, 90 mL and 220 r/min for temperature, inoculum amount, initial pH, medium volume in a 250 mL flask and shaking speed, respectively, resulting in an increase of PLA2activity from (1.87±0.12) U to (5.35±0.27) U.
Kluyveromyces lactis; phospholipase A2; fermentation
Q939.97
A
1002-6630(2015)21-0111-06
10.7506/spkx1002-6630-201521022
2015-02-13
国家高技术研究发展计划(863计划)项目(2011AA100905);江苏省“六大人才高峰”计划项目(2012-NY-002)
王辉(1988—),男,硕士研究生,研究方向为工业微生物学与酶工程。E-mail:wanghui8753@126.com
*通信作者:张梁(1978—),男,教授,博士,研究方向为酶制剂和生物能源。E-mail:zhangl@jiangnan.edu.cn
猜你喜欢
阅读(快乐英语中年级)(2022年5期)2022-06-03
昆明医科大学学报(2021年2期)2021-03-29
江西水产科技(2019年3期)2019-09-10
天津农业科学(2019年4期)2019-05-31
山东工业技术(2016年15期)2016-12-01
中国酿造(2016年12期)2016-03-01
天津医科大学学报(2015年2期)2015-12-22
应用海洋学学报(2015年3期)2015-11-22
现代检验医学杂志(2015年2期)2015-02-06
食品工业科技(2014年13期)2014-03-11
