
碱木素焦油的抑菌活性及机理
2015-12-24 07:38初雷霞江金茂赵永坡许文耀江茂生
福建农林大学学报(自然科学版) 2015年1期
初雷霞,江金茂,赵永坡,许文耀,江茂生
(1.福建农林大学生命科学学院,福建福州350002;2.福建南纸股份有限公司,福建南平353000;3.福建农林大学植物保护学院,福建福州350002)
从天然产物尤其是植物中开发高效、低毒、环境兼容性好的天然产物杀菌剂是近几年农药研究的热点[1],已得到的有抑菌作用的活性物质如萜类化合物、皂苷类、酚类、生物碱及多肽类物质等[2].但有关天然产物杀菌机理的报道较少[3].
课题组前期对竹材进行热解获得高得率的竹焦油,并且发现竹焦油对植物病原菌物和病原细菌皆有很好的抑制活性[4].鉴于碱木素是造纸工业的主要副产物,其中仅含少量纤维素和半纤维素,价廉且来源极为广泛,本文以碱木素为原料,热解制备出液化产物碱木素焦油,研究其对植物病原菌物的抑制效果和抑制敏感菌的作用机理,为将碱木素焦油工业化开发为一种新型天然产物杀菌剂奠定基础,也为天然产物杀菌剂的作用机理研究提供参考.
1 材料与方法
1.1 供试材料
将制浆废液中的碱木素酸沉、烘干后,参照江茂生等[4]的方法,用自制固定床热解设备进行热解,将收集的馏出液旋转蒸发浓缩,得碱木素焦油,再用丙酮溶解配制成10%的母液,低温贮藏备用.
1.2 供试病原菌物
16种供试植物病原菌物如表1,由福建农林大学植物病理学实验室提供.

表1 供试病原菌物及其受抑制率Table 1 The tested phytopathogenic fungi and its inhibitory ratios
1.3 试验方法
1.3.1 碱木素焦油对供试菌抑制作用的测定 采用菌丝生长抑制法进行测定.配制含1000 μg·mL-1碱木素焦油的PSA平板,以含等量溶剂的PSA作对照;在培养基上接种直径0.5 cm的供试菌菌饼,置于28℃恒温培养箱中培养.每个处理3次重复.待对照菌丝生长至平板直径2/3时,测量各处理的菌丝生长直径,计算抑制率.
1.3.2 碱木素焦油对P.capsici抑制作用的测定 参照文献[5],将碱木素焦油与甲霜灵分别配制成5个系列浓度的带毒培养基,分别接种直径0.5 cm的菌饼,置于28℃恒温培养箱中培养.每个浓度设3个重复.待对照菌丝生长至平板直径的2/3时,测量各处理菌丝生长直径,计算抑制率.
参照文献[6],将碱木素焦油与甲霜灵分别配制成5个系列浓度;制备浓度为1×105个·mL-1P.capsici孢子囊悬浮液;分别取各浓度的药剂与孢子囊悬浮液各1 mL混匀,吸取一滴置于凹玻片中央,28℃保湿培养,6 h后检查孢子囊的萌发情况,计算孢子萌发抑制率.
将抑制率转换为机率值,采用SPSS 19.0软件计算出抑制机率值与药剂浓度对数的毒力回归方程,求出EC50、EC90及其95%置信区间.计算置信区间时,采用黄昌华[7]的建议,将对照平均菌落直径毫米数作为供试生物个体数(n).
1.3.3 碱木素焦油对P.capsici有氧呼吸抑制作用的测定 采用改装的Wasburg微量气体呼吸仪测定不同浓度碱木素焦油处理下P.capsici的氧呼吸速率.将供试菌活化后,取直径为0.5 cm的菌饼置于含药的PS培养基中,利用NaOH吸收P.capsici呼出的CO2,通过数显微压差仪测定P.capsici在28℃恒温条件下的氧呼吸量.每5 min记录1次因呼吸代谢消耗氧气导致的压力减少值,连续记录120 min.以不加药同条件下P.capsici的氧呼吸量为对照.
1.3.4 碱木素焦油对P.capsici细胞膜作用的测定 采用电导率法测定碱木素焦油对P.capsici胞内电解质渗出量的影响.采用PS液体培养基培养P.capsici至对数期,收集菌丝.分别取1 g菌丝,加入到30 mL不同浓度(0、62.5、125、250、500、1000 μg·mL-1)的药剂中并使菌丝分散悬浮,立即测定菌丝悬浮液的电导率随时间的变化情况.
取上述不同浓度碱木素焦油处理P.capsici 24 h后的菌悬液,过滤,采用考马斯亮蓝法[8]测定滤液中的蛋白质含量,采用DNS法[8]测定滤液中的还原糖含量,分析碱木素焦油对P.capsici胞内蛋白质和还原糖渗出量的影响.
1.3.5 碱木素焦油对P.capsici胞外纤维素酶及多聚半乳糖醛酸酶活性抑制作用的测定 参照孙文秀[9]的方法稍作改动进行测定.将 P.capsici接种到 Czapek-Dox 液体培养基(KNO32.0 g,KCl 0.5 g,FeSO40.01 g,K2HPO41.0 g,MgSO4·7H2O 0.5 g,维生素 B10.2 g,L-天冬酰胺 0.5 g,果胶 10 g,蒸馏水 1000 mL)中静置(28℃)培养7 d后抽滤得到滤液,离心(6000 r·min-1×30 min),取上清液,即为粗酶液.
分别取1 mL酶液,加入计量碱木素焦油与酶作用5 min;加入2.5 mL缓冲液,45℃水浴下预热10 min;加1 mL 1%CMC-Na,酶解反应120 min;立即加入1.5 mL DNS,100℃煮沸5 min,流水冷却,补加水定容至20 mL;以加同剂量药且不含底物的处理溶液作为参比液,540 nm波长下测定不同浓度药剂处理的酶反应体系的光密度.将酶反应底物换成1%果胶,其他操作步骤相同.根据还原糖标准曲线读出酶反应降解CMC-Na、果胶产生的还原糖量.以45℃、pH=5.0的条件下分解多糖1 h每产生1 mg单糖为一个酶活单位[9],计算碱木素焦油作用后的纤维素酶和多聚半乳糖醛酸酶活性.
2 结果与分析
2.1 碱木素焦油对供试菌的抑制作用
由表1可以看出,碱木素焦油浓度在1000 μg·mL-1时,对16种病原菌物均有一定的抑制作用,抑制率为43.75% -100%.其中,对15种病原菌物的抑制率在60%以上,对 P.capsici、B.paeoniae、C.higginsianum、R.solani、S.rolfsii的抑制率可达 100%;对 G.candidum 的抑制率最低,为 43.75%.可见,碱木素焦油对植物病原菌物的抑制作用具有一定的广谱性.
通过多重比较发现,碱木素焦油对不同属的菌株 P.capsici、B.paeoniae、C.higginsianum、R.solani、S.rolfsii的抑制作用差异不显著(P>0.05);对同一属中,如Botrytis属中的2个菌株,Colletotrichum属中的C.glosoporioides、C.theae-sinensis和 C.higginsianum 3 个菌株,抑制作用的差异也不显著(P >0.05).由此可知,碱木素焦油对植物病原菌物的抑制作用没有明显的属特异性.但由于样本数量有限,要得出其作用规律还需进一步研究.
2.2 碱木素焦油对P.capsici的毒力
甲霜灵是防治由P.capsici引起的植物疫病的传统药剂.测定结果表明,碱木素焦油和甲霜灵对P.capsici菌丝生长的毒力回归方程分别为 y=1.65x+1.58(R2=0.9920)和 y=1.56x+0.97(R2=0.9805);对孢子萌发的毒力回归方程分别为 y=1.80x+0.65(R2=0.9317)和 y=3.07x -4.52(R2=0.9256).由表 2可见,碱木素焦油对菌丝生长和孢子萌发的EC50及EC90均低于甲霜灵,菌丝对碱木素焦油的敏感性比孢子更强.
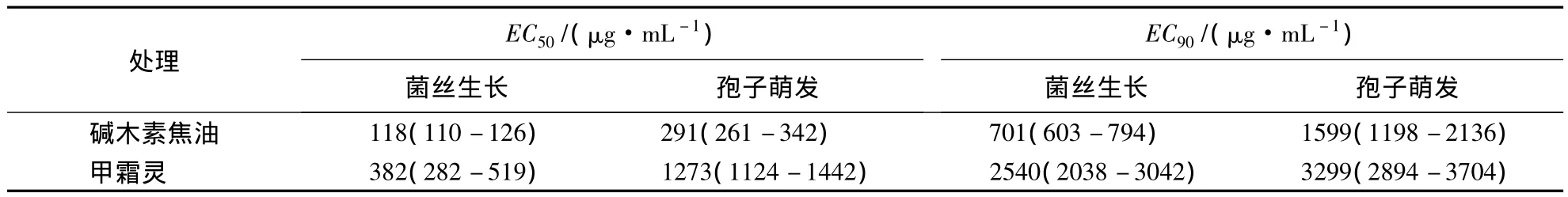
表2 碱木素焦油对P.capsici菌丝及孢子萌发的毒力Table 2 Virulence of alkali lignin tar on mycelium growth and spore germination of P.capsici
2.3 碱木素焦油对P.capsici有氧呼吸的抑制作用
测定结果(图1)表明,当用药浓度为250 μg·mL-1时,与对照相比,P.capsici的有氧呼吸明显受到抑制,且随药剂处理时间的延长作用增强;当用药浓度达到1000 μg·mL-1时,P.capsici的有氧呼吸活动微弱,处理时间达120 min时,氧气的消耗量仍很少,表明P.capsici的有氧呼吸几乎完全被抑制.
2.4 碱木素焦油对P.capsici细胞膜的作用
在菌丝量一定的情况下,菌丝悬浮液的电导率变化能够体现菌丝细胞内电解质的渗透情况,从而间接反映菌丝细胞膜的透性变化.由图2可见,对照组的电导率缓慢上升,约30 min后达到平稳;500和1000 μg·mL-1碱木素焦油均会快速提高P.capsici菌丝悬浮液的电导率;1000 μg·mL-1剂量下,菌悬液的电导率在前5 min内迅速上升,约10 min后达到平稳.

图1 碱木素焦油对P.capsici有氧呼吸的影响Fig.1 Effects of alkali lignin tar on respiration of P.capsici

图2 碱木素焦油对P.capsici电导率的影响Fig.2 Effects of alkali lignin tar on cells conductivity of P.capsici
由图3-4可以看出,随药剂处理浓度的升高,P.capsici的蛋白质及还原糖的渗透量均出现上升趋势;在处理浓度达到 500 μg·mL-1时,P.capsici的蛋白质渗透量显著提高;当浓度超过 250 μg·mL-1时,P.capsici的还原糖渗透量显著提高.
细胞膜是一种选择透过性膜,能够保持细胞内环境稳定,正常生长情况下,细胞膜内外的物质交换处于一个相对平衡的状态.P.capsici蛋白质及还原糖的渗出趋势与电解质外渗变化结果,共同反映出碱木素焦油能够作用于P.capsici的细胞膜,使P.capsici的细胞膜受损,通透性变大,从而使其胞内蛋白质及还原糖、电解质等物质透出,造成菌丝生长缓慢乃至凋亡.
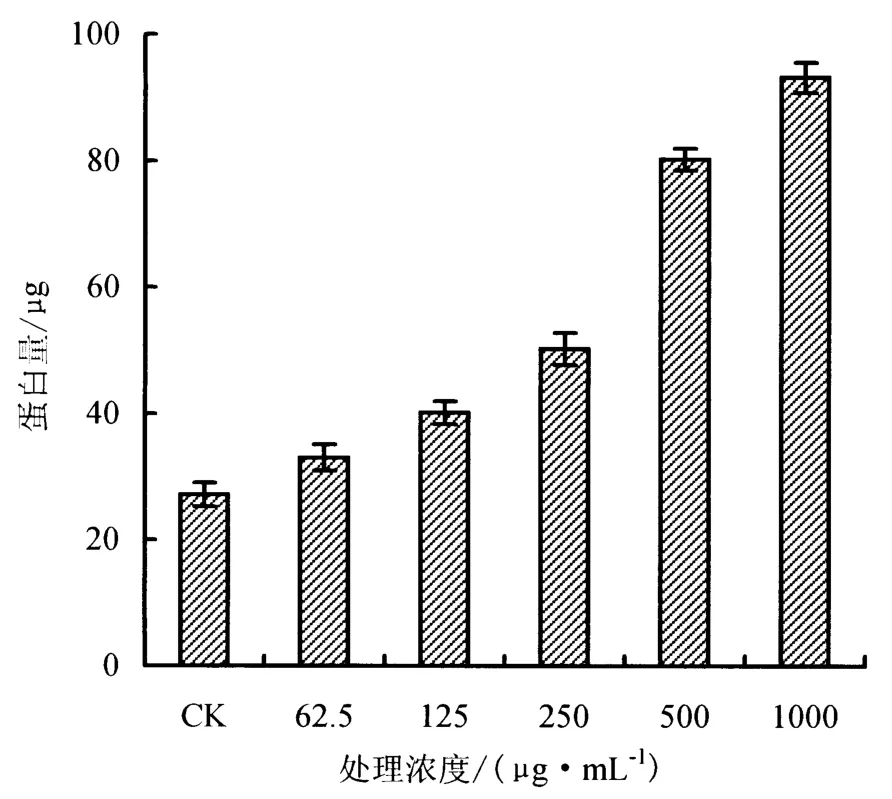
图3 碱木素焦油对P.capsici胞内蛋白渗出量的影响Fig.3 Effects of alkali lignin tar on proteins penetration from cells of P.capsici

图4 碱木素焦油对P.capsici胞内还原糖渗出量的影响Fig.4 Effects of alkali lignin tar on reducing sugar penetration from cells of P.capsici
2.5 碱木素焦油对P.capsici纤维素酶及多聚半乳糖醛酸酶活性的抑制作用
生物质热解焦油中一般含有大量的由木质素分解产生的酚类物质[4],具有很强的使蛋白质和酶变性的性能.由图5-6可以看出,随药剂处理浓度的升高,P.capsici纤维素酶及多聚半乳糖醛酸酶的活性呈现类似的递降趋势.在125 μg·mL-1浓度处理下,碱木素焦油对这2种酶的活性有轻微抑制作用;当浓度高于500 μg·mL-1时,会显著抑制这2种酶的活性,抑制率均达到40%以上,并且随药剂浓度升高抑制作用继续增强.纤维素酶及多聚半乳糖醛酸酶是P.capsici侵染寄主时释放的主要酶[10],其活性受到抑制,可以推测碱木素焦油能够降低P.capsici侵染寄主的能力.

图5 碱木素焦油对P.capsici纤维素酶活性的影响Fig.5 Effects of alkali lignin tar on cellulase enzyme activity of P.capsici

图6 碱木素焦油对P.capsici多聚半乳糖醛酸酶活性的影响Fig.6 Effects of alkali lignin tar on polygalacturonase enzyme activity of P.capsici
3 结论
本研究表明,碱木素焦油具有比较广谱的抑菌作用,对供试的10个属中的16种植物病原菌物均有不同程度的抑制作用.碱木素焦油对P.capsici菌丝及孢子萌发的EC50及EC90均低于甲霜灵.
碱木素焦油对P.capsici有复合的抑菌作用靶位点.它能够通过抑制P.capsici的有氧呼吸代谢途径导致其能量供应受到限制,使细胞内的各种需能代谢反应无法完成;同时,能够破坏P.capsici的细胞膜,使其通透性变大,损失胞内蛋白质、还原糖、电解质,导致细胞内环境紊乱;此外,能够使纤维素酶及多聚半乳糖醛酸酶活性降低而抑制菌丝生长.因此,以造纸工业的副产物碱木素为原料,热解制备碱木素焦油并开发为农用杀菌剂,对于降低造纸工业污染及促进碱木素的高值化利用具有重要意义.
[1]GURJAR M S,ALI S,AKHTAR M,et al.Efficacy of plant extracts in plant disease management[J].Agricultural Sciences,2012,3(3):425 -433.
[2]YAZDANI D,TAN Y H,ZAINAL ABIDIN M A,et al.A review on bioactive compounds isolated from plants against plant pathogenic fungi[J].Journal of Medicinal Plants Research,2011,5(30):6584 -6589.
[3]刘海燕,高微微,樊瑛.植物源杀菌剂的研究进展[J].中国农学通报,2005,21(4):254-257.
[4]江茂生,李玉玲,许文耀,等.竹焦油对植物病原细菌的抑制作用[J].福建农林大学学报:自然科学版,2012,41(5):464-468.
[5]中华人民共和国农业部.NY/T1156.2-2006农药室内生物测定试验准则杀菌剂第2部分:抑制病原真菌菌丝生长试验平皿法[S].北京:中国农业出版社,2006.
[6]中华人民共和国农业部.NY/T1156.1-2006农药室内生物测定试验准则杀菌剂第1部分:抑制病原真菌孢子萌发试验凹玻片法[S].北京:中国农业出版社,2006.
[7]黄昌华.农药试验中的问题及解决办法[J].农药科学与管理,2001(增刊):35-36.
[8]曹春蕾,崔宝凯,秦问敏.桑木层孔菌液体培养过程中几种胞外酶活性的变化[J].菌物学报,2011,30(2):275-280.
[9]孙文秀.辣椒疫霉致病遗传变异、多聚半乳糖醛酸酶基因克隆及表达特性研究[D].济南:山东农业大学,2007.
[10]DOUAIHER M N,NOWAK E.Correlative analysis of Mycosphaerella graminicola pathogenicity and cell wall-degrading enzymes produced in vitro:the importance of xylanase and polygalacturonase[J].Plant Pathology,2006,56(1):79 - 86.
猜你喜欢
食品与生物技术学报(2022年1期)2023-01-11
冶金动力(2022年5期)2022-11-08
盐科学与化工(2020年9期)2020-09-23
天津科技大学学报(2019年2期)2019-04-22
鞍钢技术(2018年2期)2018-04-13
分析化学(2017年12期)2017-12-25
合成化学(2015年10期)2016-01-17
化工进展(2015年3期)2015-11-11
——α-葡萄糖醛酸酶的研究进展*
新能源进展(2014年5期)2014-07-01
中国造纸(2014年1期)2014-03-01
