
草鱼原肌球蛋白基因的克隆及其组织表达
2015-12-21 08:05孙念陈开建褚武英肖调义李迪
湖南文理学院学报(自然科学版) 2015年2期
孙念, 陈开建, 褚武英, 肖调义, 李迪
草鱼原肌球蛋白基因的克隆及其组织表达
孙念1, 陈开建1, 褚武英2, 肖调义1, 李迪1
(1. 湖南农业大学动物科学学院, 湖南长沙, 410003; 2. 长沙学院生物工程与环境科学系, 湖南长沙, 410003)
为揭示原肌球蛋白基因在草鱼肌肉中的作用, 利用RT-PCR和RACE技术克隆获得了草鱼原肌球蛋白基因cDNA, 并对该基因在普通草鱼和脆肉鲩不同组织中的表达情况进行研究分析。结果表明原肌球蛋白基因cDNA全长序列为1 705 bp, 包含387 bp的5′UTR序列, 1 307 bp的3′UTR序列和855 bp开放阅读框(ORF)。其ORF编码284个氨基酸。系统进化分析表明普通草鱼与斑马鱼、墨西哥脂鲤的原肌球蛋白基因核苷酸同源性分别是93%和87%, 氨基酸同源性分别是96%和93%。在聚类上普通草鱼原肌球蛋白基因与其他鲤科鱼类同源性较高, 表明亲缘关系最近, 与传统分类相一致。Real time-PCR结果表明原肌球蛋白基因在所检测的普通草鱼和脆肉鲩7个组织中均有表达, 原肌球蛋白基因在普通草鱼腹肌中表达最高, 其次为前肠。原肌球蛋白基因在脆肉鲩腹肌中的表达低于普通草鱼, 而脆肉鲩中肌肉、肝脏、肾脏、前肠、后肠中原肌球蛋白基因表达量大于普通草鱼相对应组织, 但差异不显著。
普通草鱼; 原肌球蛋白; 基因表达; 脆肉鲩
作为一类结合蛋白,原肌球蛋白(tropomyosin,TPM)在细肌丝中能与肌动蛋白丝结合并与肌钙蛋白一起参与调节肌球蛋白与肌动蛋白丝对钙离子的相互作用。Perry[1]、杨静娴[2]等发现TPM以大量异构体形式行使着真核细胞内与肌动蛋白细肌丝相关的多种功能, 这是由于TPM作为一类异构体形式广泛存在于真核细胞中。这些TPM异构体共享结构形式, 大部分的同源异构体在分布上具有组织肌丝特异性, 被表达的外显子和其相应的分子区域对其相关肌丝的功能表达具有重要意义。TPM二聚化的螺旋结构使其在甲壳类和软体动物中具有高度保守性[3-5], 也成为肌肉收缩过程中重要的调节蛋白质, 同时又是甲壳类和软体动物类主要的交叉过敏原[6-8]。目前国内关于原肌球蛋白基因的研究报道不多, 李忻怡[9]等研究表明TPM在维持鱼类成体的生命活动过程中发挥重要作用, 其参与文昌鱼胚胎躯体模式的构建, 文昌鱼的肌节、肌肉调节及神经索的发生也与TPM的表达相关。李荔[10]等成功克隆出斑马鱼() TPM基因, 经免疫学鉴定发现其具有免疫过敏原的特性。不仅仅存在于肌肉细胞中, TPM也广泛并少量的存在于非肌肉细胞中, 而不同的细胞类型中同源异构体性质不同, 使得TPM在肌肉和非肌肉组织中功能不同, 包世新[11]等通过研究TPM在膀胱癌中的表达, 发现TPM有望成为一类肿瘤标记物。李晨阳[12]等发现TPM功能研究有助于提示瘢痕挛缩的发生机制。
选取规格较大的普通草鱼()饲喂3个月或半年以上浸泡了的蚕豆后就成了脆肉鲩, 即肉质脆化了的草鱼, 其肉质紧致脆爽, 不易碎。通过构建草鱼cDNA文库, 发现与普通草鱼脆化相关的重要蛋白原肌球蛋白在普通草鱼和脆肉鲩间存在表达差异显著。鉴于TPM在调控骨骼肌细肌丝活动及免疫方面的重要作用和功能, 本文对草鱼TPM基因开展克隆并对其进行表达分析研究, 为进一步对研究脆化草鱼脆化机理奠定基础。
1 材料与方法
1.1 材料
普通草鱼和脆肉鲩从湘乡市养殖场运至湖南农业大学实验室水池暂养, 正常饲喂2周后开始试验。取材时先用丁香油麻醉, 选择同样规格的普通草鱼和脆化草鱼各3尾。取不同组织后迅速丢置液氮中冷藏, 于-80 ℃保存备用。
1.2 TPM cDNA克隆
1.2.1 总RNA提取
迅速取出冷冻的普通草鱼和脆肉鲩肌肉组织于研钵中, 在液氮下研磨成粉末状。然后按照RNA iso Plus(Takara, Japan)说明书提取总RNA。
1.2.2 TPM cDNA部分序列片段克隆
根据GenBank已有报道的鱼类TPM cDNA保守序列片段设计2对引物, 克隆出2段中间片段。用肌肉中提取的总RNA为模板, 照反转录试剂盒(Clontech, USA)的说明合成第1条cDNA链, 再进行PCR扩增, 其反应体系如表1所示。

表1 反应体系
反应程序为94 ℃预变性5 min; 94 ℃ 30 s, 56 ℃ 30 s, 72 ℃ 2 min, 35个循环。3%琼脂糖凝胶电泳检测PCR扩增产物, 采用天颂生物公司的凝胶回收试剂盒纯化, 连接到pUCm-T载体并转染感受态细胞, 用于克隆测序筛选。
1.2.3 快速扩增TPM4a cDNA末端
根据1.2.2中获得的TPM中间片段和已设计好的特异性引物, 3’和5’RACE按照天颂生物公司试剂盒说明书进行扩增。3’RACE和5’RACE的特异性引物见表2。第1轮PCR反应条件为: 95 ℃3 min进行升温; 94 ℃30 s预变性, 59 ℃40 s退火, 共35个循环;72 ℃10 min延伸。第2轮PCR以第1轮PCR产物稀释100倍为模板, 将TPM-F1引物替换为TPM-F2, TPM-R1引物替换为TPM-R2, 对获得的3’RACE和5’ RACE产物分别转染Top10感受态细胞, 筛选阳性克隆, 对阳性菌落进行测序。

表2 本实验所用引物
1.3 TPM基因序列比对与分子进化树分析
将扩增出的TPM cDNA中间片段和3’, 5’末端序列进行拼接, 得到TPM cDNA全长序列。根据ExPASy(http://www.expasy.ch/ tools/)软件预测分析TPM氨基酸序列理化性质并分析TPM蛋白二级结构。采用DNA star软件对普通草鱼和已知的3个其他鱼类的TPM4a氨基酸序列(表3)进行同源性比对,并用BioEdit软件构建邻接(Neighbour- Joining, NJ)进化树。

表3 用于构建系统进化树的8个鱼类的TPM4a氨基序列登录号
1.4 TPM的组织表达差异
β-Actin作为内参照(引物), 用RT-PCR检测法检测TPM蛋白基因在草鱼不同组织的表达差异。Real-time PCR所用引物见表2。普通草鱼和脆肉鲩鱼各3尾, 麻醉解剖后取背肌、腹肌、尾肌、肝脏、肾脏、前肠和后肠等组织, 分别提取总RNA后反转录获得cDNA模板。用SYBR染料法在RT-PCR仪(ABI Prism 7300)上进行RT-PCR扩增和数据分析。反应体系见表4。

表4 RT-PCR反应体系
每种样品的目的基因和内参照基因均设有3个重复, 同时设置阴性对照。所有加样操作均在超净工作台上进行。反应程序为: 95 ℃ 30 s; 95 ℃ 5 s, 60 ℃ 32 s, 45个循环。扩增曲线和荧光值由ABI Prism 7300 SDS软件分析。用双标准曲线法计算目的基因相对于内参基因的表达量。并用SPSS 17.0进行显著性分析, 用Excel作图。
1.5 统计分析
TPM mRNA的相对表达量采用双标准曲线法分析进行差异性显著检验。最终数据采用±表示,< 0.05时被认为TPM基因在2种鱼中的表达呈显著性差异。
2 结果与分析
2.1 TPM基因的克隆及序列分析
采用同源克隆和RACE技术克隆出了普通草鱼原肌球蛋白基因TPM全长序列。该全长序列为1 705 bp, 包含387 bp 的5’UTR 序列, 1 307 bp的3’UTR序列, 855 bp开放阅读框, 编码284个氨基酸(图1)。
通过在线网站http://web.expasy.org/protparam/分析原肌球蛋白TPM的氨基酸序列, 该蛋白分子量为32 685.6, 理论等电点pI为4.68, 呈弱酸性。负电荷氨基酸残基总数为80(天冬氨酸+谷氨酸)。正电荷氨基酸残基总数为52(精氨酸+赖氨酸)。分子式为C1390H2319N391O491S10。ProtParam在线预测此蛋白不稳定指数(II)被计算为44.28, 这个蛋白是不稳定的, 脂肪指数是81.90, 平均亲水性是-1.063。由组成分布及含量可知TPM蛋白由20种氨基酸组成。其中谷氨酸含量最高, 达21.5%。其次是赖氨酸和亮氨酸, 含量分别为13.3%和12.0%。所占含量百分比最少的是苯丙氨酸, 只有0.4%。
使用SWISS-MODEL在线程序对普通草鱼TPM的氨基酸序列进行三级结构同源建模, 参与模型构建的氨基酸范围为1~283aa, TPM蛋白序列与所参考生物模板序列比对同源性为80.57%, 模型评估值为0, 说明此蛋白三级模型结构可靠。该模型中总共包括277个α螺旋, 7个无规则卷曲。
2.2 TPM氨基酸序列同源性及分子进化分析
TPM基因cDNA序列于DNAstar软件中寻找开放阅读框并翻译成氨基酸序列。将目标基因氨基酸序列置于NCBI进行核苷酸及氨基酸序列同源性比对, 查找同源蛋白序列。普通草鱼TPM与斑马鱼()、墨西哥脂鲤()的草鱼TPM核苷酸同源性分别是93%和87%, 氨基酸同源性分别是96%和93%。普通草鱼与鲤鱼和鳙鱼的氨基酸同源性分别是92%和98%。

1ACATGGGGGTTTGCAGGCAGCATCAGCTGCATTCTTCTTCTCTC 45CTAAATCTCCTAGCATCCATTAGTCTGACCACACTTTCAGCTGAAGGACCAAGAACTGCA 105CTTCGTTCGTAGCTCCCTTGAATTTCATATTCGACCCAGACACTAGATCAACCTGCAACC 165ATGGAGGCCATCAAGAAGAAGATGCAGATGCTCAAACTGGACAAGGAGAACGCCATCGAC 1MEAIKKKMQMLKLDKENAID 225CGGGCAGAGCGGGCCGAGACGGAACAGAAAGCTGCTGAGGAAAAATGCAAGCAGCTGGAC 21RAERAETEQKAAEEKCKQLD 285GACGAGCTGGTCAGCTTGCAGAAAAAGCTCAAACAAACTGAAGATGAACTGGACAAGTAC 41DELVSLQKKLKQTEDELDKY 345TCAGAGGCCCTGAAAGATGCTCAGGAGAAACTTGAACTGTCTGAGAAAAAAGCTGCTGAC 61SEALKDAQEKLELSEKKAAD 405GCTGAGGGTGATGTGGCGGGCCTGAACCGCAGAATTCAGCTGGTGGAGGAGGAACTGGAT 81AEGDVAGLNRRIQLVEEELD 465CGGGCTCAGGAGAGGCTGGGAACTGCACTGCAAAAACTGGAAGAGGCGGAGAAAGCAGCA 101RAQERLGTALQKLEEAEKAA 525GACGAGAGCGAGAGAGGCATGAAGGTAATTGAGAACCGCGCCATGAAGGACGAAGAGAAG 121DESERGMKVIENRAMKDEEK 585ATGGAGATCCAGGAGATGCAGCTGAAAGAGGCCAAACACATCGCAGAGGAGGCCGACCGC 141MEIQEMQLKEAKHIAEEADR 645AAATATGAAGAGGTGGCCCGTAAACTGGTGATTCTCGAGGGTGAGCTGGAGCGAGCTGAG 161KYEEVARKLVILEGELERAE 705GAGAGAGCCGAAGTCGCTGAATGCAAAGCTAGTGATTTGGAGGAGGAATTGAAAAACGTT 181ERAEVAECKASDLEEELKNV 765ACCAACAACCTTAAATCCCTAGAGGCTCAGGCAGAGAAATACTCGGAAAAAGAAGACAAG 201TNNLKSLEAQAEKYSEKEDK 825TATGAAGAGGAAATCAAAGTTCTTAGTGACAAGCTGAAGGAGGCCGAGACCCGTGCAGAG 221YEEEIKVLSDKLKEAETRAE 885TTTGCAGAGAGGTCGGTGGCCAAACTGGAAAAATCTATCGATGACCTGGAAGAAAAATTA 241FAERSVAKLEKSRDDLEEKL 945TCTTCTGCAAAAGAAGAAAACCTCGGAATGCATCAAGTGCTGGATCAGACACTTCAGGAG 261SSAKEENLGMHQVLDQTLQE 1005CTGAACAGCTTATAAGAGACGATGAGAGAGAACCACGTCCATTTTTACAACATTCCACAA 281L N S L * 1065ATACTCTCCTTAATTCCAACTCGCAAGCTATTGAGCCCTATTTTCTCACAGAAAAAAAAA 1125TAATTTAATTTTTTGGTCTGAGGTATTTCTTTTGCGCTTTTATCCAAGTTGGCTCGACAT 1185CGCTTCTTTCAAAAGTTAGCCAAACAACCCGAGGTGCTAATTTCCCCACTGATACAACGT 1245TATAAGTAGCTTAACACATGACTTACCACATGGAAAGTGAATATGTTTGTGTGCCTCAGC 1305ATTTCAAGTTTTGATATTAATGTATCCCGGTCAATTAAAGACGTCAATGAAGGCGTTTTC 1365AGTGCCGTGTTTTTCATAAGTACATCATGCGCTCCATTTACTGGCCACATGATGCATTGC 1425AGTACATTACTTTATTGATAATGCGATGGTATTTTGGACATGCACCATAGTACCCCGGTA 1485GCATTTGTCAAAGTGCCTGTGTGTTTTTGTGGAACCACCACGAAATGGGACCAAACAAGG 1545ACCAAAGAAATATTAGCTTTTCCCTTTTTTCTGTGAATGCGTTGTCGCTTTTTTTCTGTC 1605AAATGACTATTTCAAAATGCCTCAAATGTTTTTTTTCCATTTTCTGTGTAATAAATTTTT 1665ATAAAAAGTCAGAAAAAAAAAAAAAAAAAAAAAAAAAAAAA
用BioEdit软件构建草鱼TPM基因的系统进化树(图2)。设置重复抽样次数为1 000,从进化树可以看出,亚马逊花鱂(-)TPM基因与剑尾鱼()TPM基因首先聚类, 然后与尼罗罗非鱼()TPM基因与伯氏朴丽鱼()TPM基因聚类的分支聚类, 再与红丽鱼()TPM基因聚类, 再与草鱼TPM基因与斑马鱼TPM基因聚类后的分支聚类, 最后与墨西哥脂鲤TPM基因聚类。系统发育树结果表明, TPM基因的进化与物种的进化方向一致。
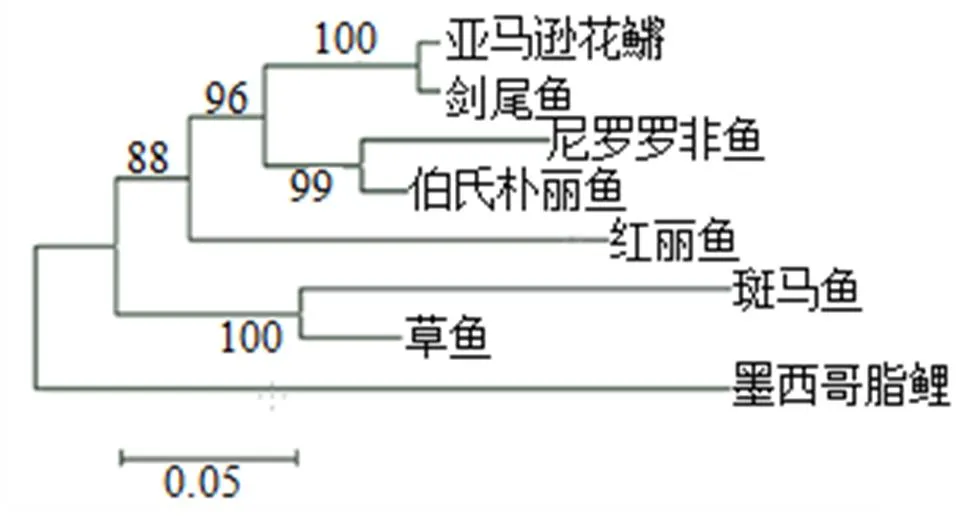
图2 TPM氨基酸序列的NJ系统进化树
2.3 普通草鱼和脆肉鲩不同组织TPM表达差异
根据普通草鱼TPM基因cDNA全长设计了荧光定量引物TPM-F和TPM-R, 以β-actin为内参基因, 使用荧光定量PCR检测了TPM基因在普通草鱼和脆肉鲩不同组织中的表达情况。结果显示, TPM基因在所检测的普通草鱼和脆肉鲩7个组织中均有表达, TPM基因在普通草鱼腹肌表达最高和其次为前肠中表达。TPM基因在脆肉鲩腹肌中的表达低于普通草鱼。而脆肉鲩中肌肉、肝脏、肾脏、前肠、后肠中TPM表达量大于普通草鱼肌肉、肝脏、肾脏、前肠、后肠组织, 且差异显著(< 0.05)(图3)。

图3 不同组织TPM mRNA相对表达丰度(平均值±标准差, n = 3), 字母不同表示有显著性差异(P < 0.05)。
3 讨论
Tropomyosin是一类分布广泛、进化保守的蛋白, 在肌肉形成和收缩过程中具有重要的调节作用, 目前在人、斑马鱼、鲟鱼、七鳃鳗、大麻哈鱼、金枪鱼、白姑鱼、斑点石斑鱼、鳕鱼等生物中有报道[13-17]。研究显示分子量为32~40 ku的原肌球蛋白基因为多数甲壳类及软体动物的主要过敏源。目前主要把原肌球蛋白当做一种过敏原来研究[18-26], 到目前为止草鱼的原肌球蛋白还没见报道。对于其在草鱼中是否有免疫作用还需进一步研究。本研究中草鱼肌肉转录组数据测得的草鱼TPM cDNA长度为1 705 bp, 包含387 bp 的5’UTR 序列, 1 307 bp的3’UTR 序列, 855 bp开放阅读框, 编码284个氨基酸。对草鱼TPM的氨基酸序列进行三级结构同源建模, 该模型中总共包括277个α螺旋, 7个无规则卷曲, 目标蛋白序列与参考模板序列比对同源性为80.57%, 可推断该蛋白结构较稳固, 其蛋白热稳定性高。国内外的报道表明, TPM蛋白序列有高度的保守性, 是包括甲壳类在内的无脊椎动物的主要过敏原, 由于TPM是肌原纤维结构蛋白越来越多的报道表明, 基于转录组数据库及RACE技术, 首次在草鱼体内克隆到草鱼TPM基因cDNA。生物信息学分析表明, 草鱼TPM与斑马鱼、墨西哥脂鲤的草鱼TPM核苷酸同源性分别是93%和87%, 氨基酸同源性分别是96%和93%。草鱼TPM与鲤鱼和鳙鱼的氨基酸同源性分别是92%和98%。在聚类上也是草鱼TPM基因与斑马鱼TPM基因聚类在一起后, 再与墨西哥脂鲤TPM基因聚类, 反映了在一同比较和聚类的鱼类中, 草鱼、斑马鱼和墨西哥脂鲤的亲缘关系近, 这都与3种鱼在分类地位同属鲤形目一致的关系, 也同样印证了本试验的聚类、与核苷酸同源性及与氨基酸同源性分析的正确性。
TPM基因调控骨骼肌细肌丝活动及免疫方面的重要作用和功能。原肌球蛋白作为一种重要的蛋白成分几乎存在于所有的动物组织中。TPM基因在所有试验的7个组织中均有较高的表达。TPM基因在普通草鱼腹肌表达最高, 其次为在前肠中表达。TPM基因在脆肉鲩中腹肌中的表达低于普通草鱼。而脆肉鲩中肌肉、肝脏、肾脏、前肠、后肠中TPM表达量大于其在普通草鱼同类组织中的表达, 且差异显著(< 0.05)。肌肉卫星细胞的增值分化能够改变肌纤维数量和肌纤维的类型, 卫星细胞的多潜能性使其转变为脂肪细胞从而增加肌内脂肪的含量, 而卫星细胞的增值分化对增加动物产量, 改良动物肉品质也有重要影响[27-29], 考虑到原肌球蛋白与肌肉收缩过程中重要的调节蛋白质, 也是真核细胞细肌丝中不可缺少的组成成分, 其与肌肉的生长和运动息息相关, 推测TPM4a基因也可能在草鱼肌肉卫星细胞增殖分化中发挥作用, 因此, 对其开展深入研究对探明草鱼脆化机理具有很重要的意义。
参考文献:
[1] Perry S V. Vertebratetropomyosin:distributionproperties and function [J]. J Muscle Res Cell Motil, 2001, 22(1): 45-49.
[2] 杨静娴. 原肌球蛋白的分子生物学研究进展[J]. 大连医科大学学报, 2004, 26(2): 26-28.
[3] Moraczewska J. Structural determinants of cooperatirity in actomyosin interaction [J]. Acta Biochimica Polonica, 2002, 49(4): 805-812.
[4] Marston S B, Redwood C S. Modulation of thin filament activation by breakdown or isoform switching of thin filament proteins [J]. Physiological and pathological implications, 2003: 805-812.
[5] Chen W Z, wen K K, Ashley E, et al. Different interaction of cardiac, skeletal muscle,and yeast tropomyosins with fluorescent (Pyrene 235)yeast actin [J]. Biophys J, 2006, 90: 1 308-1 318.
[6] Halmepuro L, Salvaggio J E, Lehrer S B. Crawfish and lobster allergens:identification and structural similarities with other crustacea [J]. Inter Archivers of Allergy and Immun, 1987, 84(2): 165-172.
[7] Leung P S, Chow W K, Duffey S, et al. Ige reactivity agaimst across-reactive allergen in crustacea and mollusca:evidence for tropmyosin as the common allergen [J]. Jouranl of Allergy and clinical Immunology, 1996, 98(5): 954-961.
[8] Lehrer S B, Mclants M L. Reactivity of IgE antibodies with crustacea and oyster allergens:evidence for common antigenic structures [J]. Journal of Allegy and Clinical Immunology, 1987, 80: 133-139.
[9] 李忻怡, 林浴霜, 张红卫. 文昌鱼tropomyosin基因的克隆、进化分析及其胚胎发育与成体中的表达模式[J]. 动物学研究, 2012, 33(4): 389-394.
[10] 李荔, 陈樱麟, 闰浩. 斑马鱼原肌球蛋白基因克隆表达及免疫学鉴定[J]. 水产科学, 2013, 32(1): 26-30.
[11] 包世新. 原肌球蛋白及其在膀胱癌中的表达[J]. 现在泌尿外科杂志, 2006, 11(3): 185-186.
[12] 李晨阳. 原肌球蛋白在增生性瘢痕中的表达与作用研究[J]. 华西医学, 2010, 25(7): 1 206-1 208.
[13] 戴建凉. 人原肌球蛋白结合蛋白3(TMOD3)cDNA的克隆及功能初步分析[J]. 复旦学报, 2000, 39(6): 681-685.
[14] 郭春生. 鲟鱼原肌球蛋白的提取、结晶与电镜观察[J]. 哈尔滨师范大学自然科学学报, 1990, 6(3): 91-94.
[15] 李刚辉. 七鳃鳗原肌球蛋白的提取, 纯化, 结晶和某些理化性质[J]. 中国第五届生物化学学术会议论文摘要汇编, 1984, 上册: 53-54.
[16] 郭桂云. 大麻哈鱼原肌球蛋白的提取结晶及电镜观察[J]. 哈尔滨师范大学自然科学学报, 1988, 4(3): 73-76.
[17] 李启沅, 李荔, 刘志刚, 等. 鱼虾贝类的主要过敏原分子免疫学特性研究进展[J]. 水产科学, 2008, 27(3): 154-156.
[18] 王春仁, 何国声. 土耳其东毕吸虫原肌球蛋白基因的原核表达[J]. 中国预防兽医学报, 2006, 28(5): 518-521.
[19] 王春仁, 仇建华. 土耳其斯坦东毕吸虫原肌球蛋白基因的原核表达[J]. 中国兽医科学, 2006, 36(3): 212-215.
[20] 梁银龙, 曹敏杰. 蟹类原肌球蛋白基因的克隆与表达[J]. 水产学报, 2009, 33(1): 25-29.
[21] 吴凯威, 刘志刚. 红星梭子蟹变应原原肌球蛋白基因的克隆、表达及其免疫学鉴定[J]. 水生生物学报, 2009, 33(2): 296-303.
[22] 杜欣军, 张伟伟. 凡纳滨对虾原肌球蛋白基因模式与重组表达[J]. 渔业科学进展, 2009, 30(4): 39-43.
[23] 崔静, 李太武. 南极养殖刺参cDNA文库的构建及原肌球蛋白基因的研究[J]. 海洋与湖沼, 2010, 41(6): 851-856.
[24] 吉色曲伍. 疥螨原肌球蛋白基因的克隆、表达与蛋白定位研究[D]. 成都: 四川农业大学, 2011.
[25] 黄素文, 杨文潮. 淡水小龙虾主要过敏原原肌球蛋白基因的克隆、表达及其免疫活性鉴定[J]. 中国免疫学杂志, 2011, 27(7): 642-647.
[26] 夏长革, 崔静. 南极养殖刺参原肌球蛋白基因的原核表达研究[J]. 海洋与湖沼, 2013, 44(1): 206-208.
[27] 尹靖东. 动物肌肉生物学与肉品质科学[M]. 北京: 中国农业大学出版社, 2011.
[28] Chen L, Wu P, Guo X H, et al. Zhang R: a Novel Regulator of MyoD expression in Fast and Slow Muscles of Siniperca chuatsi [J]. Current Molecular Medicine, 2014, 14: 370-375.
[29] Chu Wuying, Li Wenqian, Chen Lin, et al. Comparative studies on muscle cellularity and flesh quality of the two mandarin fish, Siniperca Chuatsi and Siniperca scherzeri [J]. Journal of food, Agriculture and Environment, 2013, 11(4): 345-352.
(责任编校:刘晓霞)
Molecular cloning and expression analysis of tropomyosin gene from
Sun Nian1, Chen Kaijian1, Chu Wuying2, Xiao Tiaoyi1, Li Di1
(1. Institute of Animal Science, Hunan Agricultural University, Changsha 410003, China; 2. BiologicalEngineering and Environmental Science Department,Changsha University, Changsha 410003, China)
To reveal the original role of TPM gene inmuscle, using rapid amplification of cDNA ends (RACE), the full length of TPM cDNA sequence inis cloned, and the expression of different tissues in commonand embrittlement ofare analyzed by Real-time PCR. The sequence analysis shows that the TPM cDNA had 1 705 bp in length with a 3′-untranslated region (1 307 bp), a 5′-untranslated region (387 bp), and a 855 bp open reading frame (ORF). Bioinformatics analysis predicts that the ORF of TPM4a cDNA encoded a protein of 284 amino acid residuals with a structure of protein tertiary model with 277 alpha helix and 7 random curl. The phylogenetic analysis shows that the amino acid sequence of commonTPM possessed 96% and 93% identity with the TPMs ofand, respectively. Real-time PCR results demonstrated that the abdominal muscle of commonhad the highest expression of TPM, and next the foregut. Comparing with embrittlement of, TPM shows lower expression inabdominal muscle of common, while the higher expression of embrittlement of grass carp tissues were muscle, liver, spleen, foregut and hindgut. The results indicate that TPM can be a new candidate target gene for investigating embrittlement mechanism ofembrittlement and improving healthy level of culture.
common; TPM; gene expression; embrittlement of
10.3969/j.issn.1672–6146.2015.02.016
TS 254.1
1672–6146(2015)02–0051–06
肖调义, tyxiao1128@163.com; 孙念, snty9326@163.com; 陈开健, chenkaijian2002@yahoo.com.cn。
2014-12-26
猜你喜欢
环球时报(2022-09-20)2022-09-20
当代水产(2022年7期)2022-09-20
现代食品科技(2022年4期)2022-04-28
当代水产(2022年3期)2022-04-26
今日农业(2020年24期)2020-12-15
广东第二课堂·小学(2018年9期)2018-10-24
食品与机械(2018年12期)2018-03-11
小资CHIC!ELEGANCE(2015年15期)2015-09-01
郑州大学学报(医学版)(2015年2期)2015-02-27
湖南农业(2015年5期)2015-02-26
