
特异性结合革兰氏阴性菌外膜蛋白C的单链抗体核糖体展示文库的构建
2015-12-16 07:57王秀敏管庆丰姚军虎王建华
中国饲料 2015年9期
王 潇 , 王秀敏 , 管庆丰 , 滕 达 , 姚军虎 , 王建华 *
(1.西北农林科技大学动物科技学院,陕西杨凌 712100;2.农业部饲料生物技术重点实验室,北京海淀100081;3.中国农业科学院饲料研究所基因工程研究室,北京海淀 100081)
多种革兰氏阴性菌如大肠杆菌、志贺氏菌及沙门氏菌等均为饲料中常见病原菌,会引发腹泻、败血症等疾病(周晓清,2012)。常规抗菌剂或者抗菌药多具广谱抗菌性,对正常菌群也有很强抑制效果,无差别性杀菌容易破坏肠道微生态环境,使机体遭受二次感染(Mao等,2012)。而靶向抗菌剂能选择性杀灭目标菌,对正常细菌无作用或作用较小(霍丽堵和凌均棨,2009)。特异性靶向定位区对靶向抗菌剂的靶向效应具有结构决定作用。单链抗体可变区片段 (scFv)是轻链 (VL)和重链(VH)可变区通过(Gly4Ser)3 linker连接而成的具有抗原亲和力的抗体片段,是靶向抗菌剂特异性靶向定位区的理想候选物之一(Pan等,2012;Ahmad 等,2012)。
外膜蛋白C(OmpC)在多种革兰氏阴性菌膜上均有分布,具有运输疏水性分子等生理机能,为耐药性关联蛋白,且能引发机体强烈的体液免疫反应并产生特异性抗体 (Liu等,2012a、b)。 核糖体展示技术是将抗体库体外转录翻译并在完成后阻止mRNA及蛋白脱落,形成蛋白质、核糖体与mRNA分子聚合物,进而实现亲和筛选获得与靶抗原具有高亲和性的单链抗体序列的过程(Plückthun,2012)。 近年来核糖体展示技术由于克服了基因库的转化丢失及胞内难以表达毒性蛋白的限制,作为一种筛选容量大、多样性好的方式,已成功用于抗体、酶及其他蛋白的筛选(Sun等,2012)。在核糖体展示中,构建高质量、大容量scFv库是保证筛选出高亲和力抗体可变区序列的关键。scFv具有分子量小、组织穿透力强,且含有决定抗体与抗原特异性的结构,在抗原物质检测和药物设计方面具有极高的参考价值。本课题组前期工作已完成大肠杆菌OmpC的原核重组表达(待发表),本研究将其作为抗原蛋白开展了相关工作,PCR构建核糖体展示抗体库,筛得抗OmpC蛋白的特异性scFv,旨在为构建饲料有害革兰氏阴性菌的预警检测及靶向抗菌剂和关联药物的靶向定位区设计提供参考。
1 材料与方法
1.1 材料与试剂 纯化的重组大肠杆菌外膜蛋白OmpC(C端带有6×His标签)由本实验室制备;Taq DNA聚合酶、EasyPfu高保真DNA聚合酶、DNA marker,均购自全式金生物科技有限公司;TRIZOL、逆转录PCR试剂盒,均购自TaKaRa公司;PCR产物纯化和胶回收试剂盒,均购自天根生化科技有限公司。引物由上海生工公司合成;DNA测序服务由上海生工公司提供。6~8周龄雌性BALB/c小鼠购自北京维通利华实验动物技术有限公司。
1.2 试验方法
1.2.1 大肠杆菌外膜蛋白OmpC免疫小鼠和骨髓细胞制备 将OmpC蛋白 (纯度为96%)免疫3只6~8周龄雌性BALB/c小鼠。初次免疫佐剂为完全弗氏佐剂,皮下注射;初次免疫3周后进行二免,佐剂为不完全弗氏佐剂,腹腔注射;2周后进行三免,免疫方式同二免。蛋白免疫剂量为25 μg/只[(25 μg 蛋白溶于 25 μL 磷酸盐缓冲液(PBS),加入25 μL免疫佐剂及50 μL PBS,移液器吹打乳化后 100 μL/只免疫)]。
免疫后6 d,脱颈处死小鼠。将小鼠胫骨和股骨上的肌肉和脂肪去除,并用外科手术剪减掉两端,用20 mL PBS冲洗,收集于50 mL离心管中。收集的骨髓组织用70 μm滤膜过滤,离心10 min(4 ℃,335 g),弃上清,将沉淀重悬于 3 mL 红细胞裂解液中,25℃轻柔摇动5 min。向细胞重悬液中加入 20 mL PBS,4℃、335 g离心 10 min, 弃上清,将沉淀重悬于1 mL PBS中。将上一步得到的重悬液于4℃、930 g离心5 min,获得小鼠骨髓细胞。
1.2.2 OmpC免疫小鼠骨髓细胞总RNA的提取及其cDNA逆转录 按TRIZOL法提取细胞总RNA(朱姜,2008)。按照 TaKaRa RNA PCR Kit说明书,利用随机引物Oligo(dT)l5将小鼠骨髓细胞总RNA逆转录为cDNA(孟夏萌,2008)。
1.2.3 元件、引物的设计与合成 元件合成见表1:(1)含有T7启动子、5'端茎环结构与核糖体结合位点序列(Shine Dalgarno,SD)的 scFv 上游元件序列。 (2)含有6His myc标签、gⅢ tether和 3'端茎环结构的scFv下游元件序列。

表1 构建核糖体展示文库合成元件
引物设计见表2:(1)用于扩增抗体库VL和VH 的 引 物 :VHF、VHR、VLF 和VLR (Luo 和 Xia,2012)。 (2)用于扩增带有(Gly4Ser)3 linker的 VH和VL序列的引物:VH+linker R和VL+linker F。重链下游引物及轻链上游引物均带有linker序列,以便通过重叠延伸PCR合成带有linker序列的scFv基因。(3)用于在VL基因下游连接带有6His标签的碱基序列,便于鉴定和纯化的引物:HisRev VLR。(4)重叠延伸获得连接gⅢ tether及最终序列的引物:T7B和T6te。
1.2.4 构建核糖体展示文库 核糖体展示元件构建示意如图1所示:(1)以反转录产物cDNA为模板,以 VHF、VHR,VLF、VLR 为引物分别进行 PCR 扩增 VH、VL 基因;(2)将引物 VHF、VH+linker R,VLF、VL+linker F分别混合,以VH、VL基因为模板,PCR获得VH+linker序列和VL+linker序列,用胶回收试剂盒纯化;(3)将等物质的量浓度的纯化产物VH+linker和VL+linker混合,利用引物VHF和VLR进行重叠延伸PCR,将VH与VL通过linker连接获得scFv基因库,用胶回收试剂盒纯化;(4)以引物VHF和HisRev VLR对scFv基因进行PCR扩增,获得带6His标签的scFv序列,用胶回收试剂盒纯化;(5)将等物质的量浓度的纯化产物scFv(带有6His标签)与gⅢ tether混合,利用引物VHF和T6te进行重叠延伸PCR,获得连接gⅢtether及3'端茎环结构序列;(6)纯化所获序列与T7SD序列等物质的量浓度混合,利用引物T7B和T6te进行重叠延伸PCR反应,最后获得完整序列(图1)。

表2 构建核糖体展示文库引物序列

图1 核糖体展示元件的构建过程
2 结果与分析
2.1 PCR法获得抗体库VH、VL及其连接linker序列 本课题组前期研究表明,大肠杆菌OmpC蛋白免疫BALB/c小鼠后所产生抗体对OmpC蛋白的效价高达1∶240000,对大肠杆菌菌体效价高达1∶27000,说明免疫小鼠能产生高亲和力抗体,且部分抗体可特异性结合于菌体表面蛋白区域,初步实证了scFv作为饲料中革兰氏阴性病原菌检测物质和靶向抗菌药物靶向定位区的可行性(Wang等,待发表)。此外,研究发现小鼠免疫6 d后,会有高浓度骨髓浆细胞产生,并在骨髓中较长时间内保持稳定(Reddy等,2010)。因此,小鼠免疫后第6天提取小鼠骨髓细胞总RNA,通过反转录获得抗体可变区cDNA序列,以此为模板,以 VHF、VHR,VLF、VLR 为引物扩增 VH、VL 序列(图 2)。

图2 PCR扩增获得抗体重链、轻链可变区及其连接linker序列
为保证抗体库多样性,VH上游用简并引物,且VH、VL上下游引物均依据抗体可变区框架区设计(Luo 和 Xia,2012;Reddy 等,2010)。 最终获得VH片段大小约为350 bp,VL片段大小约为700 bp(图 2), 与 Lee等 (2004) 报道相符。(Gly4Ser)3柔韧性很强,不会对抗体可变区二级结构及肽链折叠产生影响,具有维持天然抗体特异性结合性质的作用,其被作为linker广泛用于scFv库的构建。故随后以VH、VL基因为模板,以VHF、VH+linker R,VLF、VL+linker F 为引物,PCR扩增后获得VH+linker和VL+linker序列 (图2)(Luo 和 Xia,2012)。
2.2 重叠延伸 PCR构建 VH+linker+VL(scFv)和VH+linker+VL+6His myc(scFv-6His myc)序列将胶回收纯化的VH+linker和VL+linker序列等物质的量浓度混合,利用引物VHF和VLR进行重叠延伸PCR反应,结果如图3所示。由图3可见,VH与VL基因通过linker连接获得scFv基因,即VH+linker+VL序列,片段大小约为1000 bp。以切胶回收纯化的scFv基因为模板,用VHF和His-Rev VLR引物进行PCR扩增,获得带有6His标签的scFv序列(scFv-6His)。在PCR扩增过程中发现,普通PCR和重叠延伸PCR效率都不高,非特异性扩增现象明显,需要反复多次PCR扩增和琼脂糖凝胶电泳后“切胶-回收-纯化”富集目的序列。这可能与上、下游引物及文库中scFv基因序列中的高GC含量有关(陈锐,2009)。

图3 重叠延伸PCR和PCR获得VH+linker+VL序列和VH+linker+VL+6His序列
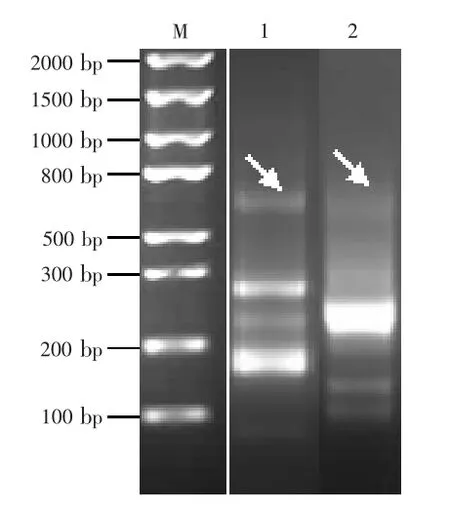
图4 重叠延伸PCR获得VH+linker+VL+6His myc+gⅢ tether+3'loop序列和T7+5'loop+SD+VH+linker+VL+6His myc+gⅢ tether+3'loop序列
2.3 核糖体展示筛选高亲和力scFv序列文库的构建 核糖体展示中,完整的元件构建决定其体外转录翻译效果。首先,为使展示蛋白更好地折叠,并与抗原蛋白结合,翻译完成后完整蛋白需脱离核糖体但不能脱落,故展示蛋白基因C端需融合一段非结构蛋白基因“tether”,由此特殊肽段锚定核糖体。此外,为防止mRNA降解,在3'及5'端添加茎环结构,能够将展示效率提高约15倍(Plückthun,2012)。 将 scFv-6His序列做胶回收纯化后与合成的gⅢtether元件等物质的量浓度混合,利用引物VHF和T6te做重叠延伸PCR,获得scFv+6His myc+gⅢtether+5'loop序列,约1300 bp(图4)。在体外转录过程中还需要强启动子(T7)以高效获得RNA及SD序列以保证翻译过程顺利进行。故将所得序列胶回收纯化后与合成的T7SD元件等物质的量浓度混合,并利用引物T7B和T6te进行重叠延伸PCR反应,结果显示获得完整核糖体展示序列,约1420 bp(图4)。在此过程中非特异性扩增现象更加严重,或与两端茎环结构有关,在重叠延伸PCR连接T7SD和gⅢtether过程中引物设计必须在茎环结构部分,这也可能是造成扩增效率不高的原因之一。在扩增过程中尝试通过改变引物碱基数目改进,但效果不佳。最终采取多次扩增、切胶回收的方式获得抗体库。
3 结论
OmpC蛋白免疫BALB/c小鼠,获得骨髓浆细胞。以其总RNA反转录获得的cDNA为模板,PCR扩增获得 VH和 VL, 并通过(Gly4Ser)3 linker连接两片段,获得scFv库。通过重叠延伸PCR法添加T7启动子、SD、gⅢ tether和两端茎环结构等元件,最终成功构建针对大肠杆菌外膜蛋白C单链抗体核糖体展示文库。为进一步通过核糖体展示法筛选高特异性、高亲和力scFv抗体奠定了基础,为饲料和其他领域革兰氏阴性病原菌检测及靶向抗菌剂和关联药物的靶向定位区的分子设计提供了新思路,也为其他关联领域scFv抗体的制备提供了参考方法和材料基础。
[1]陈锐.核糖体展示人源性scFv文库的构建及筛选抗人LBP抗体的局限性分析:[博士学位论文][D].重庆:第三军医大学,2009.
[2]霍丽堵,凌均棨.特异性靶向抗菌肽的抗变异链球菌作用[J].国际口腔医学杂志,2009,36(5):550~553.
[3]孟夏萌.运用核糖体展示技术筛选SEB抗体的初步研究:[硕士学位论文][D].天津:天津医科大学,2008.
[4]周晓清.食源性沙门氏菌的PCR快速检测技术研究:[硕士学位论文][D].郑州:河南农业大学,2012.
[5]朱姜.核糖体展示ScFv文库的构建及使用核糖体展示技术筛选壬基酚抗体:[硕士学位论文][D].扬州:扬州大学,2008.
[6]王潇.大肠杆菌组疫苗设计.免疫机理研究及抗体核糖体展示文库构建:[硕士论文][D].杨凌:西北农林科技大学,2015.
[7]Ahmad Z A,Yeap S K,Ali A M,et al.ScFv antibody:principles and clinical application[J].Clinical and Developmental Immunology,2012:980250.
[8]Luo Y,Xia Y.Selection of single-chain variable fragment antibodies against fenitrothion by ribosome display[J].Anal Biochem,2012,421(1):130~137.
[9]Liu Y F,Yan J J,Lei H Y,et al.Loss of outer membrane protein C in Escherichia coli contributes to both antibiotic resistance and escaping antibodydependent bactericidal activity[J].Infect.Immun,2012a,80(5):1815~1822.
[10]Liu C,Chen Z,Tan C,et al.Immunogenic characterization of outer membrane porins OmpC and OmpF of porcine extraintestinal pathogenic Escherichia coli[J].FEMS Microbiol Lett,2012b,337(2):104~111.
[11]Lee M S,Kwon M H,Kim K H,et al.Selection of scFvs specific for HBV DNA polymeraseusing ribosome display[J].J.Immunological Methods,2004, 284(1):147~157.
[12]Plückthun A.Ribosome display:a perspective[J].Methods Mol Biol,2012,805:3~28.
[13]Mao R Y,Teng D,Wang X M,et al.Design,expression and characterization of a novel targetedplectasin against methicillin-resistantStaphylococcus aureus[J].Appl Microbiol Biotechnol,2012,97(9):3991~4002.
[14]Pan Y,Mao W,Liu X,et al.Selection of single chain variable fragments specific for the human-inducible costimulator using ribosome display[J].Appl Biochem Biotechnol,2012,168(5):967~979.
[15]Reddy S T,Ge X,Miklos A E,et al.Monoclonal antibodies isolated without screening by analyzing the variable-gene repertoire of plasma cells[J].Nat Biotechnol,2010,28(9):965~969.
[16]Sun Y,Ning B,Liu M,et al.Selection of diethylstilbestrol-specific singlechain antibodies from a non-immunized mouse ribosome display library[J].PLoS One,2012,7(3):33186.
[17]Wang X,Guan Q F,Wang X M,et al.Paving the way to construct a new vaccine against Escherichia coli from its recombinant outer membrane protein C via model mouse,Process Biochemistry,Manuscript Number:
猜你喜欢
内蒙古民族大学学报(自然科学版)(2022年2期)2022-11-22
现代仪器与医疗(2022年4期)2022-10-08
猪业科学(2021年3期)2021-05-21
中国生殖健康(2020年2期)2021-01-18
幽默大师(2020年10期)2020-11-10
肿瘤防治研究(2020年5期)2020-07-09
中华诗词(2019年1期)2019-11-14
生物学教学(2019年9期)2019-09-23
猪业科学(2018年4期)2018-05-19
中国康复理论与实践(2015年10期)2015-12-24
