
Seed Morphology and Seedling Variation of Four Ornamental Lupin Pedigrees
2015-12-13 07:57ZhuGAOXiaolingWANGTengyunLIUSongjinGAOQiongYUFaxinYU
Zhu GAO, Xiaoling WANG, Tengyun LIU, Songjin GAO, Qiong YU, Faxin YU*
1. Jiangxi Engineering Research Center for Eco-remedication of Heavy Metal Pollution /Institute of Biological Resources, Jiangxi Academy of Sciences, Nanchang 330029, China;
2. Jiangxi Radio and TV University, Shangrao 331000, China
Plant germplasm as the material basis for plant utilization, improvement and breeding is the most precious natural wealth. Abundant species, their genes and the environments constitute the biodiversity,which plays an important role in maintaining ecological balance, environment stability and sustainable development. The utilization of germplasm resources can be divided into direct,indirect and potential ways.Genetic diversity is the core and the material basis of biodiversity. Plant variations are mainly resulted from genetic variation between individuals, the interaction between the individual genotype and the environment, as well as environmental variation[1].Intraspecific genetic variations mainly occur at three levels:variation of provenances, variation among the populations of the same provenance,variation among the individuals within the same population and variation within individuals[2]. Plant morphological characteristics are controlled by genetic basis, structural genes, regulator genes and environment. The seed characteristics of different populations differ greatly because of long-term effects of natural conditions and geographical barriers[3].Provenance variation is the basis of plant breeding,and ecology factor vari-ation is the basis for phenotypic variation. The huge phenotypic plasticity in plants is often considered to be a functional response that maximizes fitness in variable environments[4]. Seed is an important life stage for updating of angiosperms, and is also an important way for them to pass genetic information to future generations[5]. Seed morphology includes seed shape, appendages and surface structure, etc.Seed morphological characteristics like seed weight, shape and appendages reflect its adaptability to interferences[6].Seed morphology is considered to be relatively stable within the same plant species or variety, but has some differences among populations and individuals. The average seed size of individuals within populations is the same. The correlation between seed size and number is still controversial, but the reproductive output is size-dependent[7-10]. For annual plants, parents tend to maximize their reproductive output, and the offspring individuals getting higher reproductive output will be more adaptive to environment[7]. Under competitive condition, plant growth slows down and the individual becomes smaller, so the reproductive output is reduced, while increasing reproductive allocation may compensate to some extent for the reduction[10]. Therefore, fruit and seed morphology indices can be used as standards for evaluation of provenance differences and breeding work.Since 2005, we have introduced four ornamental lupin varieties from other countries[11], studied their breeding techniques[12-14]and tolerance mechanisms[15-16]. Finally, four pedigrees with ornamental values,adaptive to Jiangxi climate were selected using traditional breeding methods. In the present study, the seed characteristics of the four lupin pedigrees were compared and their variations were analyzed with an attempt to provide some references for the evaluation of seed phenotypic diversity and breeding research of ornamental lupins.
Materials and Methods
Materials
The excellent plants of the lupin pedigrees selected in 2010 were propagated successively and the seedlings grew well were selected and transplanted, and their seeds were collected in 2012.
Determine the number of samples
One hundred seeds collected from each lupin pedigree were randomly selected and measured for seed length, width and weight. Then, variation coefficients for the three traits among different numbers of seeds were calculated, as shown in Fig.1.The number of samples was determined according to the variation coefficient curves[17-18].Finally,30 multi-colored seeds, 40 pink seeds, 40 red seeds and 40 blue seeds were selected for following tests.
Measurement of seed traits
The length and width of the seeds finally selected in "Determine the number of samples" were measured using an electronic caliper, and seed weight was measured using an electronic balance. Seed specific weight was calculated using the formula as follows: Seed specific weight = Seed weight / (Seed length × Seed width).Three repetitions were set for each measurement.
Measurement of seed germination and seedling traits
Lupin seeds were sown into 32-hole pots on September 30,2013,with one seed in each hole and 60 seeds for each treatment. Three repetitions were set for each treatment. The germinated seeds were counted every day from the start to the end of germination to calculate germination rate.Before they were transplanted into filed 20 d after sowing, height of the seedlings was measured using a ruler,and their leaf and leaflet were counted;meanwhile, the survived seedlings were also counted to calculate seedling survival rate.
Germination rate(%)=Number of germinated seeds/ Number of all seedtested×100%;
Germination rate coefficient(%)=[Σ(t×n)/Σn]×100%,
wherein, t is days of germination,n stands for all the seeds germinated within t days[19];
Germination index=MDG×PV,
wherein,MDG is the mean of germinated seeds per day(the total number of germinated seeds/number germination days); PV is the maximum number of seeds germinated within one day/days from the beginning of seed germination to the day of maximum number of germinated seeds[20-23].
Statistical analysis
SPSS 16.0 was used for statistical analysis. Statistical significance was evaluated using One-Way ANOVA and Duncan’s multiple comparison.Sigmaplot 10.0 software was employed for plotting.
Results and Analysis
Seed variation among different lupin pedigrees
As shown in Table 1, the seed morphological traits differed among the four lupin pedigrees. Their seed length ranged from 4.15 to 4.37 mm,among which,the length of pink seeds was the largest, 1.06 mm more than the mean, and the length of multi-colored seeds was close to that of red seeds.The seed width of the four lupin pedigrees ranged from 2.90 to 3.07 mm, among which pink seeds were the widest, and multi-colored seeds were the narrowest. The 1 000-seed weight of the four pedigrees varied in a range of 14.45 -18.22 g, that of red seeds was the largest, and of the blue seeds was the smallest. The seed specific weight of red seeds was the largest, which was 1.33 g/mm2greater than that of blue seeds which was the smallest. The variation coefficients for the four seed morphological indices among the four lupin pedigrees were listed from large to small as: seed specific weight, 1 000-seed weight,seed width and length,suggesting that the ornamental lupins had abundant variation in seed specific weight and 1 000-seed weight. ANOVA and Duncan’s multiple comparison showed that the seed length and 1 000-seed weight had significant difference (P<0.05), while seed width and specific weight had insignificant difference among the four pedigrees(P>0.05).To sum up, the four pedigrees had variations in seed morphological traits, and the variation in 1 000-seed weight was most significance, followed by that in seed length.
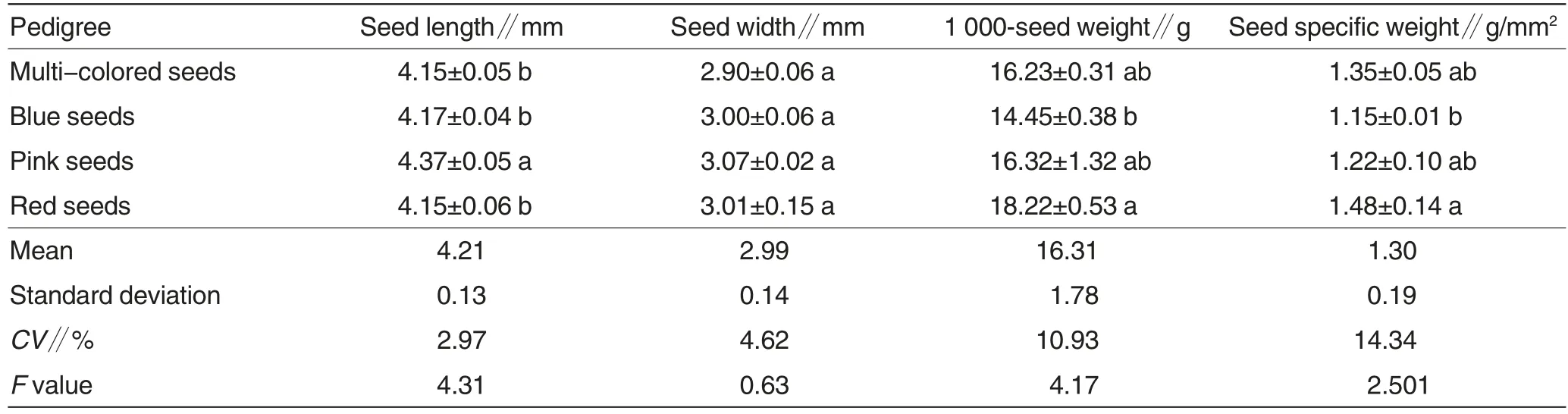
Table 1 Seed properties and multiple comparison of different lupin pedigrees
Seed germination characteristics of different lupin pedigrees
The seed germination characteristics have some differences among different varieties of a plant species.As shown in Fig.2, the seed germination characteristics of the four lupin pedigrees were similar, with slight differences. The seeds of all the four pedigrees began to germinate on the 2ndd, and the germination finished on the 5th d,so it could be concluded that the seed germination period of ornamental lupin was 5 d. The germination rate nearly reached the maximum value on the 7thd, from when several seedlings died, and water should be supplied timely. The germination potentials for red seeds and pink seeds were measured on the 3rdd,and those for multi-colored seeds and blue seeds were measured on the 4thd (Fig.2 and Fig.3). Among them, the germination potential for blue seeds was the largest,up to 68.75%,and that for pink seeds was the smallest, the former was over one fold more than the later.Among the four pedigrees, blue seeds had the highest germination rate, varied in a range from 72.92%to 84.72%,but lower than the retention rate indicated by the instruction of parental product (Fig.3).The retention rates of the four pedigrees were all measured before transplanting, and that of blue seeds was the highest, up to 88.10%,and that of red seeds was the smallest, only 74.48%. Variance analysis and Duncan’s multiple comparison on the germination potential, germination rate and retention rate revealed that the four lupin pedigrees had insignificant difference in germination rate(P=0.147 > 0.05), but significant differences (P <0.05) in germination potential (P = 0.001) and retention rate (P =0.047).
Germination rate coefficient and germination index reflects the speed of seed germination. As shown in Fig.4,the germination rate coefficient for red seeds was the highest(235.13%),and that for blue seeds was the lowest(140.86%); the germination index for pink seeds was the highest (14.08%),and that for multi-colored seeds was the lowest (8.47%).Variance analysis and Duncan’s multiple comparison showed that the four lupin pedigrees had significant differences in germination rate coefficient (P = 0.000) and germination index (P = 0.002). Among them, red and pink seeds germinated more quickly, and blue seeds germinated most slowly.
Seedling variation of different lupin pedigrees
The seedling growth traits of the four lupin pedigrees were measured on the 20thd and analyzed for their variations(Table 2).As shown in Table 2, the growth traits differed among the four pedigrees. The average seedling height of the four pedigrees was 3.97 cm, with a range of 0.76. The seedlings of multi-colored and pink pedigrees were higher than those of blue and red pedigrees. The average leaf number of the four pedigrees was 2.57, with a range of 0.34. Red and multi-colored pedigrees had more leaves than blue and pink pedigrees.The average leaflet number of the four pedigrees of the four pedigrees was 6.16, with a range of 0.43. Red and multi-colored pedigrees had more leaflets than blue pedigree. The variation coefficients of the seedling indices were listed here from large to small as plant height, leaf number and leaflet number, and the largest variation coefficient was 6.11% higher than the smallest one. Variation analysis and Duncan’s multiple comparison revealed that the four lupin pedigrees had no significant differences in plant height,leaf number and leaflet number(P>0.05).The results revealed that the seedling traits had some differences among the four ornamental lupin pedigrees, but the differences were not significant.
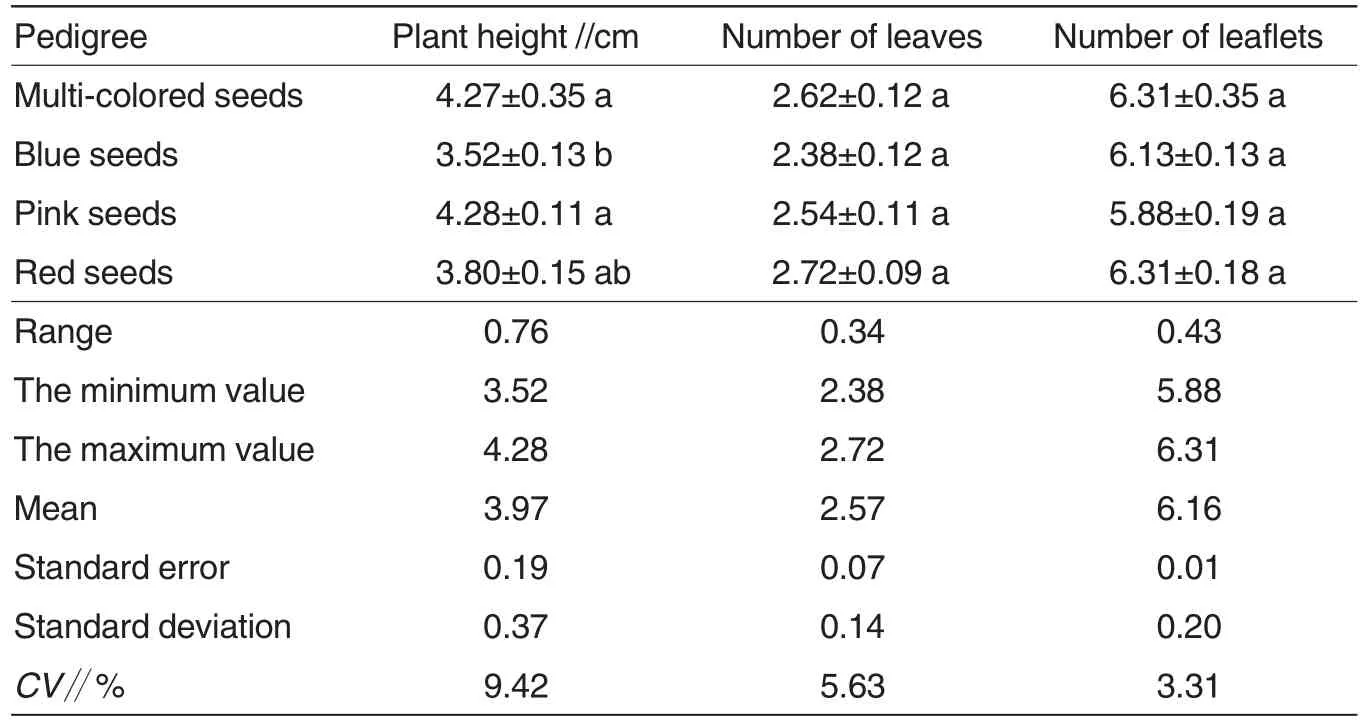
Table 2 The seedling variation of different lupin pedigrees
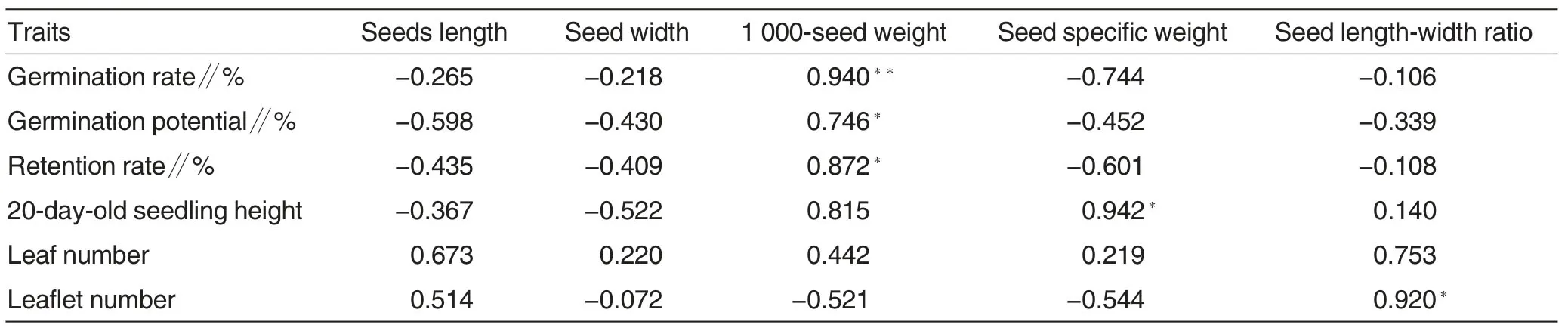
Table 3 Analysis on correlations of seed traits with seed germination and seeding growth in different pedigrees
Correlations of seed shape with seed germination and seedling growth
Seed shape is significantly correlated with seed germination and seedling growth. As shown in Table 3,both seed length and width had a negative correlation with seed germination, indicating that round seeds had advantages in germination, but thecorrelation was not significant. 1 000-seed weight had a significantly positive correlation with seed germination,indicating that heavier seeds had more advantages in germination than the lighter ones. The seed specific weight had a significantly positive correlation with 20-day-old seedling height, indicating that among the seeds of the same size the seedlings of the heavier seeds grew more quickly at late seedling stage. The ratio of seed length to seed width had a significant correlation with the number of seedling leaflets, with the correlation coefficient of 0.920. To sum up, the seed shape had certain correlations with seed germination and seedling growth of ornamental lupin, while full seeds had higher germination rate and speed,and retention rate. The ratio of seed length to seed width had a certain correlation with the number of seedling leaflets.
Conclusion and Discussion
Individual is a unit for plant lives in response to environment, while population and species are the units in plant adaption and evolution[24]. Phenotypic diversity is a result of genetic and environmental diversities,and it is an important clue to investigate genetic variation. The population grown in controlled environment can better reflect the genetic diversity, and reveal genetic law and variation degree of the species[25]. The variation of seed traits exists among and within plant populations and among individuals, and this theory is supported by the studies on Dipteronia dyeriana (Aceraceae),Gordonia acuminate, Cylcobalanopsis glauca and Sinocalycanthus chinensis[26-29]. Mehlman et al.[30]believed that the seed size had some variation among the individuals of a species,while 80%of the variation existed within individuals of S, chinensis, 85% of the variation in cones and seeds existed among and within the individuals of Abies chensiensis[31]. The morphological variation in seeds of Camillia rosthornina populations in Jinyun Mountain mainly occurred within individuals[32]. This study revealed that there were abundant variations among the four ornamental lupin pedigrees;pink seeds had the largest volume(4.37 mm long and 3.07 mm wide)and red seeds had the largest 1 000-seed weight (18.22 g); the variation coefficients for the seed morphological traits among the four pedigrees ranged from 2.97%to 14.34%;seed specific weight and 1 000-seed weight could be used as important indicators for selection in breeding because of their higher variability.
Morphological and phenotypic traits are the most simple and direct ways to detect the genetic variations of plants[33].The seed morphology of selfpollinated plant species changes little because of isolation mechanism between individuals and little gene exchange and their seeds are relatively large, while the seed morphology of cross-pollinated plant species varies greatly due to the abundant gene exchange, and their seeds are relatively small[34]. Sakai[35]believed that inbreeding within the species may help to avoid the competitions between seeds by increasing seed size,and decline in inbreeding proportion may result in small seeds. Ornamental lupins are self-pollinated, their seeds have similar morphology because of little gene exchange.There were slight variation in seed length, and significant variations in seed specific weight and 1 000-seed weight of ornamental lupins,indicating that the variation in seed weight was mainly from the variation of seed width. According to seed size and number,it could be concluded that blue pedigree had the most seeds in a pod[35].
Seed quality reflects the size and fullness of seeds. The full seeds had larger 1 000-seed weight and more abundant nutrients which can promote seed germination. In addition, 1 000-seed weight also had great influence on seedling growth and biomass.The seedlings of larger seeds were taller and had larger biomass[36-37].The seeds of the four ornamental lupin pedigrees started to germinate one day after sowing, and the germination period was 5 d.Germination rate coefficient and germination index of pink and red seeds were higher than those of blue seeds, but blue seeds had the largest germination rate(84.72%).The correlation between seed traits and germination revealed that 1 000-seed weight had significantly positive correlations with seed germination rate,germination potential and seedling retention rate, and full seeds had some advantages in germination.
Seed morphology as one of the important characteristics of genetic variation, not only determines dispersal ability of species, but also affects seed germination and seedling growth.The ratio of seed length to seed width of is closely related to seedling growth of camphor[18]; seed width had significantly positive correlation with the height of annual seedlings of Lindera aggregata[38]; seed morphology 1 000-seed weight had no significant correlations with seedling growth and tuber traits[39];but Quercus seeds had no significant relationship with seedling growth, and the seedlings of heavier seeds grew worse[40]; for Panzhihua kapok, the seedlings of longer seeds grew faster,and the seedling retention rate was higher with larger 1 000-seed weight[41]. All the studies revealed that the correlations between seed traits and seedling growth varied in different species.The seed traits of ornamental lupins had certain correlations with seed germination and seedling growth.Full seeds had higher germination rate and speed, and seedling retention rate. The ratio seed length to seed width of ornamental lupins also had a certain correlation with the number of seedling leaflets.
[1]MAO AH(毛爱华).Genetic variation and selection of provenances, families and clones of Platycladus orientalis (Linn.)Franco in Jiaxian,Henan Province(河南郏县侧柏种源、家系及无性系遗传变异与选择)[D]. Beijing: Beijing Forestry University(北京: 北京林业大学),2009.
[2]WANG QY(王秋玉).Genetic variation of the Provenance for Picea koraiensis(红皮云杉地理种源的遗传变异)[D].Harbin:Northeast Forestry University(哈尔滨:东北林业大学),2003.
[3]GREIPSSON S, DAVY AJ. Seed mass and germination behaviour in populations of the dune building grass Leym usarenarius[J]. Annals of Botany, 1995,76:493-501.
[4]COLEMAN JS, CONNAUG HAY KD,ACKERLY DD. Interpreting phenotypic variation in plants[J]. Trends in Ecology&Evolution,1994,9(5):187-191.
[5]ZHANG DY(张大勇). 植物生活史进化与繁殖生态学[M].Beijing:Science Press(北京:科学出版社),2004.
[6]HENDRY GAE, GRIME JP. Methods in comparative plant ecology: a laboratory manual [M]. London: Chapman & Hall,1993.
[7]FENNER M.Seed ecology [M].London and New York: Chapman and Hall,1985.
[8]SILVERTOWN JW, LOVETT DJ. Introduction to plant population biology[M].Oxford: Blackwell Scientific Publications,1993.
[9]SCHIMD B, BAZZAZ FA, WEINER J.Size dependent of sexual reproduction and of clonal growth in two perennial plants [J]. Canadian Journal of Botany,1995,73:1831-1837.
[10]ZHU ZH (朱志红), WANG G (王刚).Studies on the phenotypic plasticity and reproductive allocation in Avena sativa L.(燕麦表型可塑性与繁殖分配的研究)[J].Journal of Lanzhou University(Natural Sciences)(兰州大学学报:自然科学版),2002,38(1):76-82.
[11]WANG XL (王小玲),ZENG DQ (曾德庆),GAO Z (高柱),et al.Advances of study and technological strategies in China on lupins (羽扇豆研究进展及其在我国发展的技术策略)[J]. Jiangxi Science(江西科学),2007,25(4):442-495.
[12]WANG XL, GAO Z, LIU T, et al. Primary establishment of the tissue culture technique and regeneration system for ornamental Lupinns polyphyllus[J].Journal of Northeast Agricultural University,2009,16(3):7-12.
[13]WANG XL(王小玲),GAO Z(高柱),YU FX (余发新), et al. Study on propagation technology of ornamental lupins(观赏羽扇豆繁殖技术研究)[J].Journal of Anhui Agricultural Sciences (安徽农业科学),2008,36(29):12653-12655.
[14]WANG XL(王小玲),GAO Z(高柱),YU FX (余发新),et al.Study on influence factors of transplanting survival rate on ornamental lupins test-tube plantlets(观赏羽扇豆试管苗移栽成活率的影响因素研究)[J].Northern Horticulture(北方园艺),2011,(14):62-64.
[15]WANG XL(王小玲),GAO Z(高柱),YU FX (余发新),et al.Physiological effect of exogenous salicylic acid on ornamental lupin under high-temperature stress (外源水杨酸对观赏羽扇豆高温胁迫的生理响应)[J].Chinese Agricultural Science Bulletin (中国农学通报),2011,27(25):89-93.
[16]WANG XL(王小玲),GAO Z(高柱),YU FX(余发新),et al.Changes in activities of two protective enzymes and MDA content in leaf of ornamental lupins at different growth stages (观赏羽扇豆叶片不同生育期二种保护酶活性和丙二醛含量变化规律)[J].Northern Horticulture(北方园艺),2010,(10):40-43.
[17]SNEDECOR GW,COCHRAN WG,Statistical Methods [M].Ames Iowa:The Iowa State University Press,19807.
[18]YAO XH(姚小华).Study on the genetic variation and selection of camphor (樟树遗传变异与选择的研究)[D].Changsha: Central South University of Forestry and Technology(长沙: 中南林学院),2002.
[19]BOSCAGLI A,SETTE B.Seed germination enhancement in Satureja montana L.ssp.Montana[J].Seed Science and Technology,2001,29:347-355.
[20]BONGERS FPE, POPMA J. Leaf dynamics of seeding of rain forest species in relation to canpony gaps[J].Oecologia,1990,82:122-127.
[21]ABDUL-BAKI AA, ANDERSON JD.Relationship between decarboxylation of glutamic acid and vigor in soybean seed[J].Crop Science,1973,13:222-226.
[22]MIHIC M, MODI P. Metabolic syndrome risk factors for atherosclerosis and diabetes[J]. Curr Diabetes Rev,2008,4(2):122-128.
[23]SANTOS DL, PALMEIRA CM, SEICA R, et al. Diabetes and mitochondrial oxidative stress: a study using heart mitochondria from the diabetic Goto-Kakizaki rat [J]. Mol Cell Biochem,2003,246(1/2):985-995.
[24]GU WC (顾万春). Statistical genetics( 统计遗传学)[M]. Beijing: Science Press(北京: 科学出版社),2004.
[25]YAN AM(阎爱民),CHEN WX(陈文新).Phenotypic feature diversity of Rhizobia isolated from Medicago sp, Melilotus sp. and Caragana sp.(苜蓿、 草木樨、 锦鸡儿根瘤菌的表型多样性分析)[J].Biodiversity Science (生物多样性),1999,7(2):112-118.
[26]LI S(李珊), CAI YL(蔡宇良), XU L(徐莉), et al. Morphological differentiation of samaras and seeds of Dipteronia dyeriana (Aceraceae)(云南金钱槭果实、种子形态分化研究)[J].Plant Diversity and Resources (云南植物研究),2003,25(5):589-595.
[27]DENG HP (邓洪平), HE P (何平),ZHONG ZC (钟章成).The studying of seeds morphological differentiation among populations of Gordonia acuminata from different geographical provenances and successiving communities (不同地理种源及演替群落的四川大头茶居群种子形态分化研究)[J].Journal of Southwest China Normal University(Natural Science Edition)(西南师范大学学报: 自然科学版), 1999,24(2):207-213.
[28]CAI YL (蔡永立),WANG XH (王希华),SONG YC (宋永昌). Variation of fruit size and its shape of Cylcobalanopsis glauca in the eastern subtropical zone,China (中国东部亚热带青冈果实形态变异的研究)[J]. Acta Ecologica Sinica(生态学报),1999,19(4):581-586.
[29]CAI YL(蔡琰琳),JIN ZX(金则新).Morphological variation of fruits and seeds in endangered plant Sinocalycanthus chinensis(濒危植物夏腊梅果实、种子形态变异研究)[J]. Journal of Northwest Forestry University (西北林学院学报),2008,23(3):44-49.
[30]MEHLMAN DW. Seed size and seed packaging variation in Baptisia lanceolata(Fabaceae) [J]. American Journal of Botany,1993,80(7):735-742.
[31]SUN YL (孙玉玲), LI QM (李庆梅),YANG JY (杨敬元),et al.Morphological variation in cones and seeds in Abies chensiensis (秦岭冷杉球果与种子的形态变异)[J].Acta Ecologica Sinica(生态学报),2005,25(1):176-181.
[32]CAO GX (操国兴),ZHONG CZ (钟章成),XIE DL (谢德体),et al.A preliminary study on the morphological variation of seeds of Camillia rosthornina populations in mt.Jinyun (缙云山川鄂连蕊茶种子形态变异的初步研究)[J].Journal of Southwest University (西南农业大学学报),2003,25(2):105-107.
[33]ZHU LB(竺利波), GU WC(顾万春), LI B(李斌).Study on phenotypic diversity of population in Cersis chinensis (紫荆群体表型性状多样性研究)[J].Chinese Agricultural Science Bulletin(中国农学通报),2007,23(3):138-145.
[34]MA SB (马绍宾),JIANG HQ (姜汉侨).Study on the seed weight and seed seize variation pattern and their biological significance in Podophylloideae(Berberidaceae)(小檗科鬼臼亚科种子大小变异式样及其生物学意义)[J].Acta Botanica Boreali-Occidentalia Sinica(西北植物学报), 1999, 19(4): 715-724.
[35]SAKAI S.Using phylogenies to explain seed size variation among plant [J].Journal of Evolutionary Biology, 1996,9(2):243-252.
[36]MA SB (马绍宾),JIANG HQ (姜汉侨),HUANG HY (黄衡宇),et al.A primary study on seed production of medicinal plant Sinopodophyllum hexandrum(药物植物桃儿七不同种群种子产量初步研究)[J].Chinese Journal of Applied Ecology (应用生态学报), 2001, 12(3):363-368.
[37]WU GL(武高林),DU GZ(杜国祯).Relationships between seed size and seedling growth strategy of herbaceous plant:A review(植物种子大小与幼苗生长策略研究进展)[J]. Chinese Journal of Applied Ecology (应用生态学报),2008,19(10):191-197.
[38]CHEN LH(陈丽华),JIANG JM(姜景民),LUAN QF (栾启福), et al. Study on seed morphology of Lindera aggregata from 14 areas(乌药种子性状产地表型变异研究) [J]. Journal of Zhejiang Forestry Science and Technology (浙江林业科技),2005,25(1):9-11.
[39]LI GL (栗宏林),ZHANG ZX (张志翔),ZHANG X(张鑫). Study on seed morphology and seedling growth of Jat-ropha curcas L. from different provenances(小桐子不同产地种子性状及苗期生长差异研究)[J]. Journal of Arid Land Resources and Environment (干旱区资源与环境),2010,24 (2): 204-208.
[40]WANG QM (汪企明),LI XC (李晓储),HUANG LB(黄利斌),et al.Variation in seed and seedlings of American Quercus 美国栎属种源引种、 变异研究:种子及苗期生长变异 [J]. Journal of Jiangsu Forestry Science Technology(江苏林业科技),1998,26(1):1-6.
[41]GAO Z (高柱). Studies on key techniques for scale plantation of kapok(木棉产业化栽培关键技术研究)[D].Kunming: Southwest Forestry University(昆明: 西南林业大学),2012.
猜你喜欢
中华医学图书情报杂志(2022年1期)2022-11-18
北京航空航天大学学报(2022年8期)2022-08-31
中国现代医生(2022年21期)2022-08-22
北京航空航天大学学报(2022年5期)2022-06-06
绿色科技(2021年21期)2021-11-26
北京航空航天大学学报(2021年4期)2021-11-24
北京航空航天大学学报(2021年9期)2021-11-02
食品安全导刊(2021年21期)2021-08-30
天津医科大学学报(2021年1期)2021-01-26
三农资讯半月报(2020年2期)2020-03-09
 Agricultural Science & Technology2015年1期
Agricultural Science & Technology2015年1期
- Agricultural Science & Technology的其它文章
- Analysis on Resistance of Rice Cultivar Lianjing 7 to Rice Black-streaked Dwarf Disease
- Effects of Anti-wind Erosion with Peanut Stubbles in Sandy Lands during Fallow Period
- Influences of Various Environmental Factors on the Degradation of Deoxynivalenol in Wheat Grains
- Reduction of Cd,Cu,Ni and Pb Mobility by Active Si
- Effect of Maize Sowing Time and Density on Stem and Leaf Morphological Characters of Soybean in Relay-cropping System
- Effects of Planting Density on Yield and Mechanical Harvesting Loss Rate of Brassica napus L.