
应激颗粒和抗病毒先天性免疫
2015-11-19 07:27董路娜孙英杰胡桂学
中国动物传染病学报 2015年4期
董路娜,孙英杰,郑 航,胡桂学,丁 铲
(1.吉林农业大学动物科学技术学院,长春 130118;2.中国农业科学院上海兽医研究所,上海 200241)
应激颗粒和抗病毒先天性免疫
董路娜1,孙英杰2,郑 航1,胡桂学1,丁 铲2
(1.吉林农业大学动物科学技术学院,长春 130118;2.中国农业科学院上海兽医研究所,上海 200241)
病毒感染哺乳动物细胞过程中,病毒复制产生的基因组或病毒蛋白激活先天性免疫信号通路后,产生干扰素的同时诱导干扰素下游基因表达,促进细胞的抗病毒反应。与此同时,病毒感染宿主细胞还能引起胞质颗粒的聚集,其成份大多为核糖核蛋白,RNA结合蛋白、翻译起始因子等,这个聚集区域被称之为应激颗粒(stress granules,SG)。先天性免疫通路和SG都是宿主细胞对病毒感染的一种抵抗性反应,而与之相对,一些病毒却通过抑制SG和先天性免疫通路的形成来促进自身复制。由于SG中包含RNA结合蛋白以及一系列病毒或宿主的RNA,而先天性免疫中识别病毒RNA的模式识别分子也能特异性结合病毒RNA,那SG如何协同先天性免疫在宿主抗病毒中发挥一定的作用,本文就SG在病毒感染诱导的先天性免疫中的作用进行简要综述。
应激颗粒;病毒;先天性免疫;RNA结合蛋白
病毒感染宿主细胞能够激活宿主的一系列抗病毒反应,除了获得性免疫之外,通过模式识别受体(pattern recognition receptro,PRRs)识别病毒从而诱导产生干扰素是一种重要的先天性抗病毒机制,除此之外,近年来发现的应激颗粒(stress granule,SG)也被越来越多的证明和病毒的复制有着重要的关系,鉴于SG成分及形成机制的特殊性,不难推测其在病毒诱导的先天性免疫中发挥重要的作用。本文着重对病毒感染过程中SG和先天性免疫间关系的最新进展进行综述,可以为深入了解病毒与宿主细胞互作以及控制病毒的感染提供一个新的思路。
1 病毒感染和应激颗粒
1.1 应激颗粒 细胞在受到各种不同程度的损害如热休克、氧化应激、营养饥饿等刺激时,为抵抗这些不良刺激,在胞质中形成核蛋白聚集成的非核膜包裹的聚集区域,称为SG。一般认为SG是细胞停滞翻译的现象,作为防止异常蛋白合成的一种手段。停滞翻译的核蛋白在外界压力消失的情况下恢复转录和翻译,或者被P小体(processing bodies,PB)分解[1,2]。
多种RNA结合蛋白(RNA binding proteins,RBPs)存在于SG中,其中一些经典的蛋白可以用作SG的标记蛋白,通过荧光可检测到。有些蛋白是SG形成和维持其稳定状态的关键因素,如Ras-GAP SH3结合蛋白1(RasGTPase-activating proteinbinding protein 1,G3BP1)是磷酸化依赖性核酸内切酶,T细胞限制性胞内抗原1(T cell restricted intracellularantigen-1,TIA1)和TIA相关蛋白(TIA-related protein,TIAR)合称TIA/TIAR。通过对这些调节因子基因的敲除或者RNA干扰可起到一定的抑制SG形成的作用[3,4]。然而,SG形成的具体分子生物学机制还不清楚。
细胞受到应激之后eIF-2α第51位丝氨酸磷酸化是导致SG形成的主要原因。4种eIF-2α激酶:双链RNA依赖性激酶(protein kinase R,PKR)、总蛋白调节激酶(general control non-derepressible kinase,GCN2)、PKR样内质网激酶(PKR-like endoplasmic reticulum kinase,PERK)、亚铁血红素eIF-2α激酶(heme-regulated eIF-2α kinase,HRI)可以使哺乳动物eIF-2α磷酸化。eIF-2α磷酸化导致eIF2-GTP-tRNAMet三聚体量的减少,而eIF2-GTP-tRNAMet对于将tRNA装配至48S前体复合物的过程至关重要,eIF2-GTP-tRNAMet的不足导致RNA结合蛋白TIA-1和TIAR等代替三聚体结合48S复合物,促进了多核糖体的解聚和SG的形成[2]。
1.2 病毒感染和应激颗粒 病毒尤其是RNA病毒可以引起胞质中SG样小体形成,部分病毒通过激活eIF2α激酶PKR和GCN2诱导SG形成,当细胞检测到细胞质中的病毒RNA时,这两种因子被激活并发挥一定作用。病毒感染宿主细胞诱导SG形成的模式有多种:形成稳定SG、不形成SG或是SG形成后很快被分解等(表1)。
1.2.1 病毒感染诱导暂态SG形成 在小RNA病毒感染宿主细胞过程中,如脊髓灰质炎病毒、柯萨奇病毒和脑心肌炎病毒,SG形成时间短暂与病毒3C蛋白酶水解G3BP1有关[6,7]。另外,丙型肝炎病毒(Hepatitis C virus,HCV)感染能够形成SG,但在感染后期分解。HCV通过其基因组中5'端未翻译区域激活PKR从而诱导SG形成,但压力应激诱导DNA损伤诱导基因34(growth arrest and DNA damage -inducible transcript 34,GADD34)的表达在感染后期介导eIF2α去磷酸化从而抑制SG形成。当恢复压力应激时,一种未知机制促使GADD34表达下调,细胞中eIF2α被磷酸化,这样的环境变化导致SG反复形成与分解[13,14]。
1.2.2 病毒感染诱导稳态SG形成 腺病毒5型感染宿主细胞能够诱导稳定SG形成,有趣的是,缺失E1A蛋白的腺病毒5型感染细胞后。诱导了数轮SG的形成,其具体机制还有待进一步证实[7]。呼吸道合胞体病毒感染全程均能检测到SG的表达,且随着感染时间的延长SG数量增多,将SG形成关键基因G3BP1敲除之后病毒复制能力有一定程度降低[21]
2.3 病毒感染抑制SG形成 门戈病毒和鼠科动物脑脊髓炎病毒都是心肌病毒属的核酸病毒成员,由于病毒非结构蛋白L蛋白抑制SG形成,因此这些病毒感染细胞时不形成SG。L蛋白是通过抑制IRF-3活化来阻止干扰素(interferon,IFN)的产生,但L蛋白抑制SG形成的机制尚不清楚[8]。同样麻疹病毒感染细胞后,细胞中也不产生SG,病毒C蛋白在其中发挥一定的作用,与野生型毒株相比,C蛋白缺失的病毒能明显的诱导SG产生。尽管麻疹病毒C蛋白在感染过程中起多种作用,如调节病毒聚合酶活性、抑制IFN产生,但抑制SG形成的分子机制还有待进一步研究[20]。甲型流感病毒感染过程中,病毒非结构蛋白1(nonstructural protein 1,NS1)通过RNA隔离或其他理化作用阻断PKR激活通路从而抑制eIF2α磷酸化[25]这些研究结果证实SG有一定的抗病毒作用,相应的,病毒也在一定程度上抑制SG的形成。
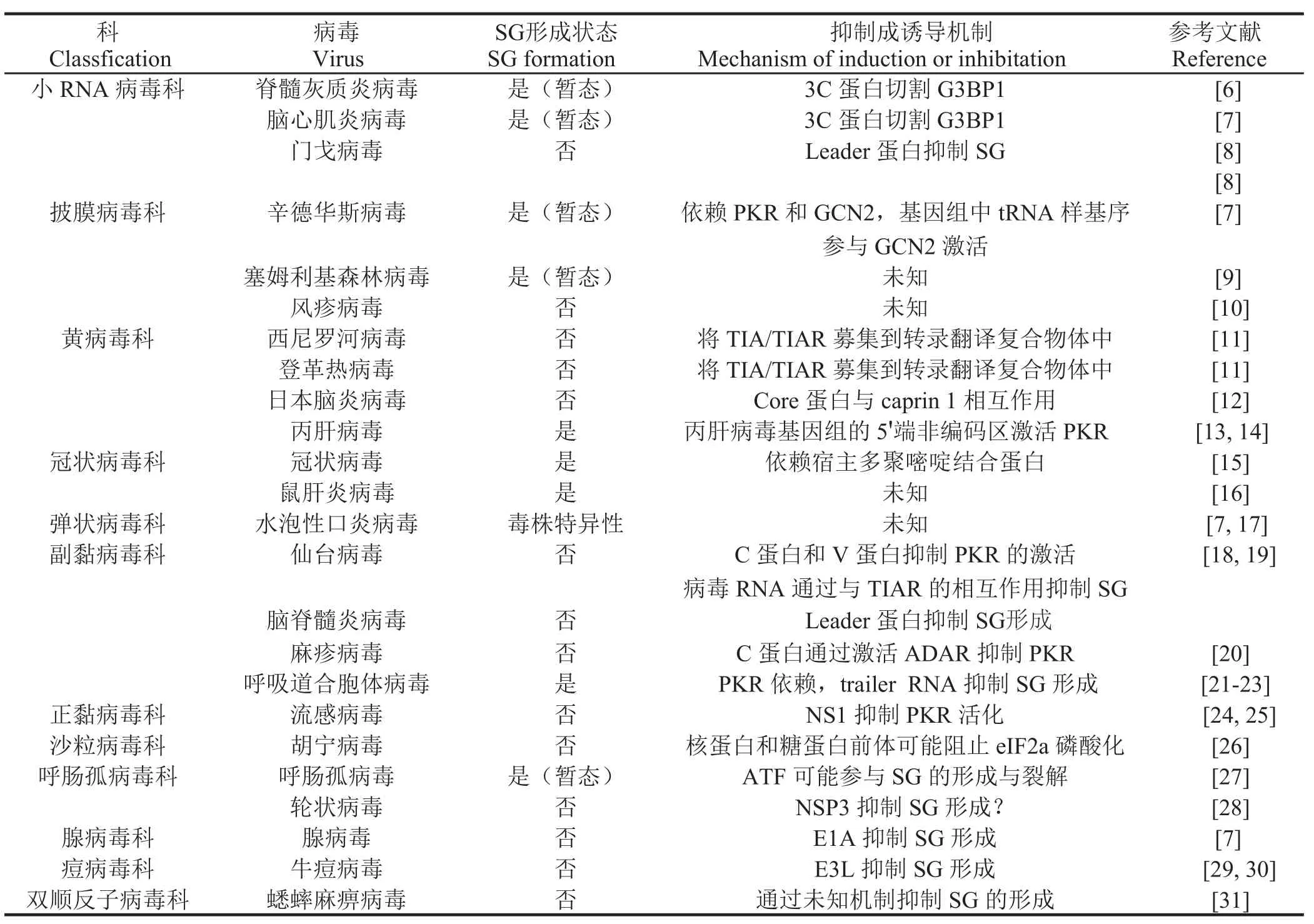
表 1 病毒感染与SG形成Table 1 Viral infection and SG formation
2 应激颗粒和抗病毒先天性免疫
2.1 RLRs通路在SG形成中的作用 对流感病毒的研究结果显示,NS1缺失流感病毒能够诱导细胞产生稳定SG,在SG中能够检测到RIG-I、黑素瘤差异化相关基因5(melanoma differentiation-associated protein5,MDA5)、遗传学和生理实验室蛋白2(laboratory of genetics and Physiology 2,LGP2)等抗病毒蛋白,同时还能检测到病毒RNA,根据敲除和RNA干扰实验证实SG的抗病毒作用,并将其命名为抗病毒SG(antiviral SG,avSG),说明SG能够作为宿主细胞的抗病毒平台识别非自身RNA进而发挥其抗病毒作用[25]。与此矛盾的是,L蛋白失活的门戈病毒感染宿主细胞也能产生SG,SG中能够检测到识别小RNA病毒科病毒的受体MDA5,但是进一步实验结果显示,MDA5介导的干扰素通路既不诱导SG也并非SG形成所必需[36]。最新的研究结果显示,以双链RNA模拟病毒核酸刺激细胞为模型,将RIG-I、MDA5和MAVS分别RNA干扰,发现只有MAVS的敲低影响了细胞内SG的形成,而RIG-I和MDA5的敲低对SG形成并无影响,进一步研究显示MAVS通过其CARD区域与PKR直接相互作用,促使PKR的二聚化激活,从而通过PKR-eIF2α通路诱导SG形成[37]。RIG-I是一种DExD/H box类RNA解旋酶,DHX36作为另一种DExD/H box类RNA解旋酶通过调控双链RNA(dsRNA)依赖的PKR的激活参与RIG-I信号通路的激活,同时也能调控PKR依赖的抗病毒SG(avSG)的形成,协助病毒RNA的特异性识别[38]。
2.2 SG在病毒诱导的先天性免疫中的作用 在肠道病毒感染细胞过程中,SG形成关键基因G3BP1的PxxP结构域对SG的形成,细胞招募PKR至SG中的过程以及PKR介导的eIF2α必需,说明G3BP1能够在转录和翻译水平调控宿主先天性免疫,在细胞应激反应与先天性免疫之间起到了桥梁作用[39]。在流感病毒感染中,通过将SG形成关键基因G3BP1或PKR的RNA干扰导致IFN产量的下降和病毒滴度的上升,说明G3BP1和PKR参与了流感病毒诱导的先天性免疫过程[25]。由于PKR-eIF2α通路是病毒诱导宿主细胞SG产生的经典途径,同时PKR还是重要的干扰素刺激因子,有报道显示PKR还参与了IFN的产生[40]。鉴于PKR分子的这种特殊性,其在SG与抗病毒先天性免疫的联系中可能起到了至关重要的作用(图1)。
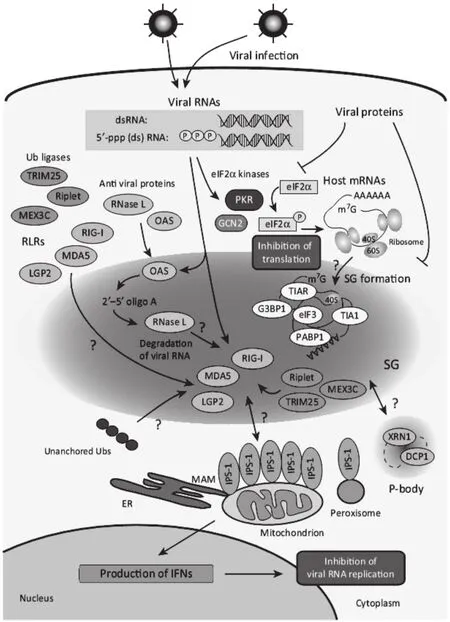
图1 应激颗粒与先天性免疫互作模式图Fig. 1 The model of the interaction between SGs and innate immunity
2.3 SG与ISGs介导的mRNA调控 已有报道显示细胞在受到应激反应时,多种ISGs被招募到SG中,其中包括聚(ADP-核糖)聚合酶(PARP)蛋白、型莫洛尼白血病病毒10同源蛋白(MOV10)、RNA腺苷脱氨酶ADAR1(ADAR1)等,这些蛋白均参与了miRNA介导的靶基因沉默。
2.3.1 PARPs 许多聚(ADP-核糖)聚合酶家族蛋白均是ISGs,其中被鉴定出来最著名的抗病毒蛋白是PARP13,或称之为锌指抗病毒蛋白(zinc-finger antiviral protein,ZAP),由IRF-3介导的I型IFN刺激分泌[41],ZAP有两个剪辑突变体ZAPL(PARP13.1)和ZAPS(PARP13.2)[42]在所有PARP成员中,PARP12和ZAP的两个剪辑突变体ZAPL和ZAPS均被证实位于SG中[43]其中在亚砷酸钠诱导的氧化应激中,PARP12与经典的SG标记物G3BP1、TIAR以及转录起始因子3η共定位,有一些证据证明,PARP12可能是一种SG特异性PARPs发挥对ZAP和Ago2(另一种PARP)的ADP-核糖基化作用。由于许多ISGs发挥抗增殖的作用,因此其在细胞中的本底表达量很低且受到严格调控,据猜测与其他基因相比,更多数量的微RNA(microRNA,miRNA)在正常生理状态下结合在ISGs的3'UTR发挥阻遏作用[44],而病毒感染状态下ISG的mRNA去阻遏从而上调表达抑制病毒复制。ADP核糖在去除RNA诱导沉默复合物(RNA-induced silencing complex,RISC)活性上调ISGs表达过程中发挥重要的作用,ZAPS、ZAPL和PARP12则作为重要的组分参与了应激状态下RISC活性的释放,但其中的具体机制还不清楚。
2.3.2 MOV10 MOV10蛋白是RNA解旋酶超家族1的成员,在C端有保守的解旋酶基序[45]。MOV10作为一种I型IFN刺激因子,能够抑制HCV和逆转录病毒的复制,但是却能够协助丁型肝炎病毒的转录[46,47]。MOV10能够在纯化的Ago1和Ago2复合物中鉴定到,位于PB中,功能上参与了miRNA介导的mRNA沉默[48]。MOV10能通过完美匹配miRNA介导mRNA的切割、通过不完美匹配miRNA介导蛋白翻译的抑制。与此对应的是,APOBEC3G作为另一种ISG同样也位于PB中,通过干扰MOV10和Ago2的互作抑制miRNA功能,导致RISC复合物的异常组装和成熟[49]。概括的说,MOV10与Ago2的互作对于RISC的功能至关重要,但是MOV10对miRNA不同的抑制机制仍然有不同的结论,需要进一步证实。
2.3.3 ADAR1 ADAR家族成员能识别特定RNA底物分子中的腺苷(A)并催化其水解脱氨为肌苷(I),A到I的RNA编辑导致细胞将I作为鸟苷(G)来识别,从而导致遗传密码子的改变。其中ADAR1被认为参与病毒基因组的编辑过程,导致了病毒基因组的多样性,也发挥抑制病毒复制的作用[50]。ADAR1分为p110和p150两个亚型,其中p150亚型存在于氧化应激和干扰素诱导的SG中,p 1 5 0的这种定位依赖于其特有的Z-D N A结合结构域[51]。ADAR1通过直接与RNAi通路成员互作或者miRNA的A到I剪辑发挥促进或者抑制miRNA功能,不同的报道之间有一定的冲突,还没有定论。在麻疹病毒(Measles virus,MV)的研究中发现,MV感染后PKR和ADAR1与SG标志物G3BP1共定位,将ADAR1敲除后MV诱导SG增加,将ADAR1的p150亚型进行互补表达能够恢复对SG的抑制,说明ADAR1能够负调控MV诱导的SG[20]。
2.4 SG与其他细胞抗病毒成分 除了以上经典的先天性免疫分子之外,还有其他一些抗病毒因子被证明与病毒诱导的SG密切相关。例如ATP依赖的RNA解旋酶DDX3X,参与了包括先天性免疫在内的许多细胞功能中,在HCV感染过程中发挥正向调控作用,HCV的3'UTR能够与DDX3X和IKK-α互作并共定位至SG中。在HCV感染的细胞中,多数HCV非结构蛋白在SGs和脂滴(lipid droplet,LD)附近与DDX3X共定位,通过将DDX3X和SG关键组分敲低能显著抑制HCV复制,说明DDX3X通过与HCV蛋白和RNA互作、SG和LD表面的动态互作在HCV感染过程中发挥作用[52]。在粘液瘤病毒和牛痘病毒感染过程中,M062蛋白能够通过直接互作抑制抗病毒蛋白SAMD9功能,而缺失M062蛋白的粘液瘤病毒能够诱导抗病毒颗粒的形成,与经典的SG成分有所不同,其形成不依赖于eIF2α而依赖于SAMD9,因此被命名为SAMD9样颗粒[53]。
3 小结
SG作为一种细胞针对应激的一种反应对生物维持自稳态至关重要,SG形成与IFN的产生量有密切的相互关系,许多RLRs和其他抗病毒蛋白以及病毒RNA或蛋白都聚集于SG中,这些证据表明SG很有可能作为一种先天性免疫的“平台”发挥作用,广义上说,是否SG与细胞自噬类似,本身就是动物进化过程中形成的先天性抗病毒机制?是宿主发挥抗病毒反应中重要的一个环节?了解SG和先天性免疫如如何协同抵抗病毒感染对于开发新型抗病毒策略和预防病毒感染有重要的理论意义。
[1]Kedersha N, Chen S, Gilks N, et al. Evidence that ternary complex(eIF2-GTP-tRNA(i)(Met))-deficient preinitiation complexes are core constituents of mammalian stress granules[J]. Mol Biol Cell, 2002, 13(1): 195-210.
[2]Kedersha N, Anderson P. Stress granules: sites of mRNA triage that regulate mRNA stability and translatability[J]. Biochem Soc Trans, 2002, 30(Pt 6): 963-969.
[3]Gilks N, Kedersha N, Ayodele M, et al. Stress granule assembly is mediated by prion-like aggregation of TIA-1[J]. Mol Biol Cell, 2004, 15(12): 5383-5398.
[4]Tourriere H, Chebli K, Zekri L, et al. The RasGAP associated endoribonuclease G3BP assembles stress granules[J]. J Cell Biol, 2003, 160(6): 823-831.
[5]Onomoto K, Yoneyama M, Fung G, et al. Antiviral innate immunity and stress granule responses[J]. Trends Immunol,2014, 35(9): 420-428.
[6]White J P, Cardenas A M, Marissen W E, et al. Inhibition of cytoplasmic mRNA stress granule formation by a viral proteinase[J]. Cell Host Microbe, 2007, 2(5): 295-305.
[7]Ng C S, Jogi M, Yoo J S, et al. Encephalomyocarditis virus disrupts stress granules, the critical platform for triggering antiviral innate immune responses[J]. J Virol, 2013, 87(17): 9511-9522.
[8]Borghese F, Michiels T. The leader protein of cardioviruses inhibits stress granule assembly[J]. J Virol, 2011, 85(18): 9614-9622.
[9]Mcinerney G M, Kedersha N L, Kaufman R J, et al. Importance of eIF2alpha phosphorylation and stress granule assembly in alphavirus translation regulation[J]. Mol Biol Cell, 2005, 16(8): 3753-3763.
[10]Matthews J D, Frey T K. Analysis of subcellular G3BP redistribution during rubella virus infection[J]. J Gen Virol,2012, 93(Pt 2): 267-274.
[11]Emara M M, Brinton M A. Interaction of TIA-1/TIAR with West Nile and dengue virus products in infected cells interferes with stress granule formation and processing body assembly[J]. Proc Natl Acad Sci U S A, 2007, 104(21): 9041-9046.
[12]Katoh H, Okamoto T, Fukuhara T, et al. Japanese encephalitis virus core protein inhibits stress granule formation through an interaction with Caprin-1 and facilitates viral propagation[J]. J Virol, 2013,87(1): 489-502.
[13]Toroney R, Nallagatla S R, Boyer J A, et al. Regulation of PKR by HCV IRES RNA: importance of domain II and NS5A[J]. J Mol Biol, 2010, 400(3): 393-412.
[14]Garaigorta U, Heim M H, Boyd B, et al. Hepatitis C virus (HCV) induces formation of stress granules whose proteins regulate HCV RNA replication and virus assembly and egress[J]. J Virol, 2012, 86(20): 11043-11056.
[15]Sola I, Galan C, Mateos-Gomez P A, et al. The polypyrimidine tract-binding protein affects coronavirus RNA accumulation levels and relocalizes viral RNAs to novel cytoplasmic domains different from replication-transcription sites[J]. J Virol, 2011,85(10): 5136-5149.
[16]Raaben M, Groot Koerkamp M J, Rottier P J, et al. Mouse hepatitis coronavirus replication induces host translational shutoff and mRNA decay, with concomitant formation of stress granules and processing bodies[J]. Cell Microbiol, 2007, 9(9): 2218-2229.
[17]Dinh P X, Beura L K, Das P B, et al. Induction of stress granule-like structures in vesicular stomatitis virus-infected cells[J]. J Virol, 2013, 87(1): 372-383.
[18]Takeuchi K, Komatsu T, Kitagawa Y, et al. Sendai virus C protein plays a role in restricting PKR activation by limiting the generation of intracellular double-stranded RNA[J]. J Virol, 2008, 82(20): 10102-10110.
[19]Iseni F, Garcin D, Nishio M, et al. Sendai virus trailer RNA binds TIAR, a cellular protein involved in virus-induced apoptosis[J]. EMBO J, 2002, 21(19): 5141-5150.
[20]Okonski K M, Samuel C E. Stress granule formation induced by measles virus is protein kinase PKR dependent and impaired by RNA adenosine deaminase ADAR1[J]. J Virol, 2013, 87(2): 756-766.
[21]Lindquist M E, Lifland A W, Utley T J, et al. Respiratory syncytial virus induces host RNA stress granules to facilitate viral replication[J]. J Virol, 2010, 84(23): 12274-12284.
[22]Lindquist M E, Mainou B A, Dermody T S, et al. Activation of protein kinase R is required for induction of stress granules by respiratory syncytial virus but dispensable for viral replication[J]. Virology, 2011, 413(1): 103-110.
[23]Hanley L L, Mcgivern D R, Teng M N, et al. Roles of the respiratory syncytial virus trailer region: effects of mutations on genome production and stress granule formation[J]. Virology, 2010, 406(2): 241-252.
[24]Khaperskyy D A, Hatchette T F, Mccormick C. Influenza A virus inhibits cytoplasmic stress granule formation[J]. FASEB J, 2012, 26(4): 1629-1639.
[25]Onomoto K, Jogi M, Yoo J S, et al. Critical role of an antiviral stress granule containing RIG-I and PKR in viral detection and innate immunity[J]. PLoS One, 2012, 7(8): e43031.
[26]Baird N L, York J, Nunberg J H. Arenavirus infection induces discrete cytosolic structures for RNA replication[J]. J Virol, 2012, 86(20): 11301-11310.
[27]Qin Q, Hastings C, Miller C L. Mammalian orthoreovirus particles induce and are recruited into stress granules at early times postinfection[J]. J Virol, 2009, 83(21): 11090-110101.
[28]Montero H, Rojas M, Arias C F, et al. Rotavirus infection induces the phosphorylation of eIF2alpha but prevents the formation of stress granules[J]. J Virol, 2008, 82(3): 1496-1504.
[29]Simpson-Holley M, Kedersha N, Dower K, et al. Formation of antiviral cytoplasmic granules during orthopoxvirus infection[J]. J Virol, 2011, 85(4): 1581-1593.
[30]Katsafanas G C, Moss B. Colocalization of transcription and translation within cytoplasmic poxvirus factories coordinates viral expression and subjugates host functions[J]. Cell Host Microbe, 2007, 2(4): 221-228.
[31]Khong A, Jan E. Modulation of stress granules and P bodies during dicistrovirus infection[J]. J Virol, 2011, 85(4): 1439-1551.
[32]Sadler A J, Williams B R. Interferon-inducible antiviral effectors[J]. Nat Rev Immunol, 2008, 8(7): 559-568.
[33]Pichlmair A, Reis E, Sousa C. Innate recognition of viruses[J]. Immunity, 2007, 27(3): 370-383.
[34]Honda K, Takaoka A, Taniguchi T. Type I interferon[corrected] gene induction by the interferon regulatory factor family of transcription factors[J]. Immunity, 2006, 25(3): 349-360.
[35]Schoggins J W, Wilson S J, Panis M, et al. A diverse range of gene products are effectors of the type I interferon antiviral response[J]. Nature, 2011, 472(7344): 481-485.
[36]Langereis M A, Feng Q, Van Kuppeveld F J. MDA5 localizes to stress granules, but this localization is not required for the induction of type I interferon[J]. J Virol, 2013, 87(11): 6314-6325.
[37]Zhang P, Li Y, Xia J, et al. IPS-1 plays an essential role in dsRNA-induced stress granule formation by interacting with PKR and promoting its activation[J]. J Cell Sci, 2014, 127(Pt 11): 2471-2482.
[38]Yoo J S, Takahasi K, Ng C S, et al. DHX36 enhances RIG-I signaling by facilitating PKR-mediated antiviral stress granule formation[J]. PLoS Pathog, 2014, 10(3): e1004012.
[39]Reineke L C, Lloyd R E. The stress granule protein G3BP1 recruits protein kinase R to promote multiple innate immune antiviral responses[J]. J Virol, 2015, 89(5): 2575-2589.
[40]Zhang S, Sun Y, Chen H, et al. Activation of the PKR/eIF-2alpha signaling cascade inhibits replication of Newcastle disease virus[J]. Virol J, 2014, 11: 62.
[41]Wang N, Dong Q, Li J, et al. Viral induction of the zinc finger antiviral protein is IRF3-dependent but NF-kappaB-independent[J]. J Biol Chem, 2010, 285(9): 6080-6090.
[42]Kerns J A, Emerman M, Malik H S. Positive selection and increased antiviral activity associated with the PARP-containing isoform of human zinc-finger antiviral protein[J]. PLoS Genet, 2008, 4(1): e21.
[43]Leung A K, Vyas S, Rood J E, et al. Poly(ADP-ribose)regulates stress responses and microRNA activity in the cytoplasm[J]. Mol Cell, 2011, 42(4): 489-99.
[44]Seo G J, Kincaid R P, Phanaksri T, et al. Reciprocal inhibition between intracellular antiviral signaling and the RNAi machinery in mammalian cells[J]. Cell Host Microbe, 2013,14(4): 435-445.
[45]Fairman-Williams M E, Guenther U P, Jankowsky E. SF1 and SF2 helicases: family matters[J]. Curr Opin Struct Biol,2010, 20(3): 313-324.
[46]Wang X, Han Y, Dang Y, et al. Moloney leukemia virus 10(MOV10) protein inhibits retrovirus replication[J]. J Biol Chem, 2010, 285(19): 14346-14355.
[47]Haussecker D, Cao D, Huang Y, et al. Capped small RNAs and MOV10 in human hepatitis delta virus replication[J]. Nat Struct Mol Biol, 2008, 15(7): 714-721.
[48]Meister G, Landthaler M, Peters L, et al. Identification of novel argonaute-associated proteins[J]. Curr Biol, 2005,15(23): 2149-2155.
[49]Liu C, Zhang X, Huang F, et al. APOBEC3G inhibits microRNA-mediated repression of translation by interfering with the interaction between Argonaute-2 and MOV10[J]. J Biol Chem, 2012, 287(35): 29373-29383.
[50]Ko N L, Birlouez E, Wain-Hobson S, et al. Hyperediting of human T-cell leukemia virus type 2 and simian T-cell leukemia virus type 3 by the dsRNA adenosine deaminase ADAR-1[J]. J Gen Virol, 2012, 93(Pt 12): 2646-2651.
[51]Ng S K, Weissbach R, Ronson G E, et al. Proteins that contain a functional Z-DNA-binding domain localize to cytoplasmic stress granules[J]. Nucleic Acids Res, 2013,41(21): 9786-9799.
[52]Pene V, Li Q, Sodroski C, et al. Dynamic interaction of stress granules, DDX3X, and IKK-alpha mediates multiple functions in Hepatitis C virus infection[J]. J Virol, 2015,89(10): 5462-5477.
[53]Liu J, Mcfadden G. SAMD9 is an innate antiviral host factor with stress response properties that can be antagonized by poxviruses[J]. J Virol, 2015, 89(3): 1925-1931.
STRESS GRANULE AND ANTIVIRAL INNATE IMMUNITY
DONG Lu-na1, SUN Ying-jie2, ZHENG Hang1, HU Gui-xue1, DING Chan2
(1. College of Animal Science and Technology, Jilin Agricultural University, Changchun 130118, China; 2.Shanghai Veterinary Research Institute, CAAS, Shanghai 200241, China)
In the course of virus infection, virus genome and protein produced by virus replication initiate innate immunity signaling pathway, leading to interferon secretion and interferon stimulate gene expression and ultimately establish an antiviral state. Meanwhile,virus infection also induced stress granule (SG) containing ribonucleoprotein, RNA binding protein and eukaryotic initiated factors, etc. Innate immunity and SG are both cellular antiviral response. Correspondingly, virus promotes its own replication through inhibit SG formation and innate immunity. SG contains RNA binding protein and a series of virus or host RNA, while the molecular associated to innate immunity such as pattern recognition receptor also specifically recognize virus RNA, indicating their close relationship. This review will focus on the strategies how SG and innate immunity collaboratively exert antiviral function.
Stress granule; virus; innate immunity; RNA binding protein
S 852.42
A
1674-6422(2015)04-0065-07
2015-05-04
国家自然科学基金青年基金(31400144);上海市青年科技英才杨帆计划(14YF1408000); 中国博士后科学基金面上一等资助(2014M550107)
董路娜,女,硕士研究生,预防兽医学专业
丁铲,E-mail: shoveldeen@shvri.ac.cn;胡桂学,E -mail : huguixue901103@163.com
猜你喜欢
中老年保健(2022年1期)2022-08-17
肝博士(2022年3期)2022-06-30
今日农业(2021年21期)2021-11-26
中老年保健(2021年12期)2021-08-24
昆明医科大学学报(2021年8期)2021-08-13
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
小读者之友(2020年4期)2020-05-15
猪业科学(2018年8期)2018-09-28
小星星·阅读100分(高年级)(2015年11期)2015-11-28
