
吲哚乙酸对冬虫夏草生物量、胞外多糖产量及其抗氧化性能的影响
2015-11-05 08:33张志红吴素蕊邰丽梅赵天瑞
食品工业科技 2015年12期
张志红,吴素蕊,邰丽梅,樊 建,赵天瑞,*
(1.昆明理工大学云南省食品安全研究院,云南昆明650500;2.中华全国供销合作总社昆明食用菌研究所,云南昆明650223)
吲哚乙酸对冬虫夏草生物量、胞外多糖产量及其抗氧化性能的影响
张志红1,吴素蕊2,邰丽梅2,樊建1,赵天瑞1,*
(1.昆明理工大学云南省食品安全研究院,云南昆明650500;2.中华全国供销合作总社昆明食用菌研究所,云南昆明650223)
通过在液体培养基中添加不同浓度的吲哚乙酸,研究吲哚乙酸对冬虫夏草生物量、胞外多糖产量及其抗氧化性能的影响。结果表明:适宜浓度的吲哚乙酸可以提高冬虫夏草的生物量和胞外多糖产量,并且随吲哚乙酸浓度的增加二者均呈先增后降的变化趋势,当浓度为2.5μg/mL时,生物量和胞外多糖产量均达到最高,分别为18.3g/L和2.9g/L,较不添加吲哚乙酸分别增加了6.3%和15.4%;同时,适宜浓度的吲哚乙酸还可以提高冬虫夏草胞外多糖的抗氧化性能,在1.0μg/mL和2.5μg/mL时,胞外多糖对羟基自由基和DPPH的清除率分别达到最高,为58.9%和56.6%。
冬虫夏草,吲哚乙酸,生物量,胞外多糖,抗氧化
冬虫夏草[Cordyceps sinensis(Berk.)Sacc.]又名虫草,隶属于真菌门(Eumycota)、子囊菌亚门(Ascomycotina)、核菌纲(Pyrenomycetes)、麦角菌目(Clavicipitales)、麦角菌科(Clavicipitaceae)、虫草属(cordyceps)[1]。冬虫夏草是虫草菌寄生于蝙蝠蛾(Hepialus armoricanus)等昆虫幼体内形成的菌物复合体,是一种极其名贵的中草药[2],主要分布在我国青海、西藏、四川、云南等地海拔3000~5100m的高寒草甸区[3]。传统中医认为,冬虫夏草性温和、味甘后微辛,具有止血化痢、补精益髓、益肾、保肺、止痨咳等功效[4]。现代医学研究表明,冬虫夏草具有免疫调节、抗癌、调节内分泌、抗菌、促进造血、抗病毒、护肝以及抗惊厥等生理活性作用[5],具有很高的研究利用价值。
目前,关于植物激素在促进植物细胞生长发育等方面的作用已经有了比较全面的认识[6]。近年来,关于植物激素对于食用菌的作用影响也时见报道,如据上官端琳等[7]报道,适当浓度的α-萘乙酸可以提高黑脉羊肚菌的生物量和胞内多糖产量。据王谦等[8]报道,赤霉素能有效提高金福菇菌丝的生长速度,当浓度为1.0mg/L时,生长速度提高达15.77%。据马荣山等[9]报道,α-萘乙酸能较好地促进草原白磨菌丝体的生长。然而,植物激素类物质对冬虫夏草的作用影响却鲜见报道。吲哚乙酸作为植物激素类物质,不仅能够促进细胞的分裂和伸长,使细胞的生物量增加,还可以调节细胞的代谢活动,促进代谢产物的合成。因此,本实验选取吲哚乙酸为实验材料,研究了吲哚乙酸对冬虫夏草生物量、胞外多糖产量及其抗氧化性能的影响,以期为进一步开发利用虫草资源提供理论与实践依据。
1 材料与方法
1.1材料与仪器
冬虫夏草菌由昆明食用菌研究所提供;葡萄糖、蛋白胨、酵母膏、琼脂、KH2PO4、MgSO4·7H2O、吲哚乙酸(IAA) 均购自昆明鼎国生物技术有限公司;苯酚、乙醇、H2SO4、DPPH、FeSO4·7H2O、水杨酸、H2O2、甲醇等其他试剂均为国产分析纯。
BSC-250型恒温恒湿培养箱上海博讯实业有限公司医疗设备厂;QYC-2102C型光照振荡培养箱宁波江南仪器厂;TU1901型双光束紫外可见分光光度计北京普析通用仪器有限责任公司;TGL-16G型离心机上海安亭科学仪器厂;ES-315型高压灭菌锅TOMY KOGYO CO.LTD;AL204型分析天平梅特勒-托利多仪器(上海)有限公司;ZD-F12型真空冷冻干燥机南京载智自动化设备有限公司;N-1100型旋转蒸发仪上海爱朗仪器有限公司。
1.2实验方法
1.2.1培养基配制PDA培养基:马铃薯20%,葡萄糖2%,琼脂2%,pH自然;液体培养基:葡萄糖4%,酵母膏1%,蛋白胨0.5%,KH2PO40.1%,MgSO4·7H2O 0.05%,pH自然。液体培养基按比例配制好后,按每瓶100mL的装液量分装,于121℃高压灭菌30min。
1.2.2接种与培养菌种活化:将实验所需菌株接种到PDA培养基上,18℃恒温恒湿培养14d,4℃冰箱保藏备用。
发酵培养:为防止高温使IAA失活,因而在接种前将滤菌后的IAA乙醇溶液按实验设定的浓度加入到液体培养基中,每个水平做3个平行实验。将活化好的菌种分别接种到含有不同浓度(0、1.0、2.5、5.0、7.5、10μg/mL)IAA的液体培养基中,接种量为6个带菌琼脂块(约0.1cm2/块)。完成后,将其置于振荡培养箱中,在25℃,180r/min条件下培养7d。
1.2.3生物量的测定发酵结束后,将发酵液离心,弃上清液,得菌丝体,用蒸馏水反复洗涤3~4次,5000r/min离心10min后冻干,测菌丝体干重。
1.2.4胞外多糖的提取参照朱朝阳等[10]的方法稍作修改,提取胞外多糖。发酵液在5000r/min的条件下离心10min,取上清液,并将其真空浓缩至原体积的1/4,加入其3倍体积的95%的乙醇进行醇沉,4℃静置过夜,5000r/min离心10min,弃上清液,再用75%的乙醇洗涤2~3次,5000r/min离心10min,取沉淀冻干,得粗多糖。
1.2.5胞外多糖含量的测定将粗多糖溶解,并稀释至合适的倍数,采用苯酚-硫酸法[11]测定其吸光度,根据标准曲线计算多糖含量。

葡萄糖标准溶液的配制:准确称取干燥的葡萄糖1.0000g置于100mL容量瓶中,用蒸馏水溶解,定容至刻度,配制成10mg/mL的葡萄糖标准溶液,摇匀备用。
葡萄糖标准曲线的绘制:准确吸取葡萄糖标准溶液0、0.2、0.4、0.6、0.8、1.0mL,分别置于20mL的比色管中,加蒸馏水稀释至2.0mL,以蒸馏水为空白对照,分别加入6%的苯酚溶液1.0mL,迅速加入浓硫酸5mL,混匀,沸水浴15min后,用冷水冷却。于490nm波长处,测吸光值A,以葡萄糖浓度为横坐标,吸光值A为纵坐标绘制葡萄糖标准曲线,得回归方程:
y=5.4387x-0.0378(R2=0.9994)
1.2.6胞外多糖抗氧化性能的测定将在不同浓度IAA作用下,发酵所得的冬虫夏草胞外多糖分别配制成2mg/mL的多糖溶液,在测定羟基自由基清除能力和DPPH自由基清除能力时,分别按浓度梯度(0.4、0.8、1.2、1.6、2.0mg/mL)和(0.2、0.4、0.6、0.8、1.0mg/mL)进行稀释,以测定其体外抗氧化性能。
1.2.6.1羟基自由基清除能力测定参照Smirnoff[12]的方法稍作修改,在10mL离心管中分别加入9.0mmol/L FeSO4溶液1mL,1mL样品溶液,9.0mmol/L H2O21mL,摇匀后静置10min,加入9.0mmol/L水杨酸-乙醇溶液1mL,摇匀,37℃保温30min,冷却,3000r/min离心10min,取上清液于510nm处,测吸光值Ax。为去除多糖本身对吸光值的影响,以等体积蒸馏水代替水杨酸为样品对照组,测吸光值Ax0;以等体积蒸馏水代替多糖为空白对照组,测吸光值A0。计算公式如下:
1.2.6.2DPPH自由基清除能力测定参照Blois[13]的方法稍作修改,配制2mmol/L DPPH甲醇溶液并避光保存,取1mL待测样液加入2mL上述DPPH溶液,充分混匀,25℃恒温避光反应30min,在517nm处测定吸光值Ax;以等体积的甲醇溶液代替DPPH甲醇溶液作为样品对照组,测吸光值Ax0;以等体积蒸馏水代替样品溶液作为空白对照组,测吸光值A0。计算公式如下:
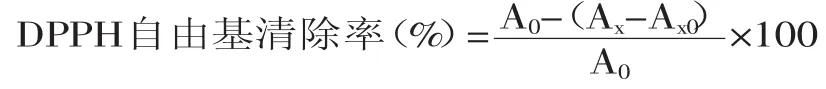
1.3数据处理
采用软件Origin 8.0及相关方法对实验数据进行处理和分析。
2 结果与讨论
2.1不同浓度IAA对冬虫夏草生物量的影响
不同浓度IAA作用下的冬虫夏草生物量结果如图1所示。
由图1可知,在0~10μg/mL范围内,随着IAA浓度的增加,冬虫夏草菌丝量呈现出先增后降的变化趋势,在2.5μg/mL时菌丝体干重达到最大,为18.3g/L,较不添加IAA增加了6.3%;同时,当IAA浓度>5.0μg/mL时,冬虫夏草的生长受到抑制,并随其浓度的增加而明显增强。结果表明,适宜浓度的IAA可以促进冬虫夏草生物量的增加,并随IAA浓度的增加而呈现先增后减的变化趋势,但过高浓度的IAA会对冬虫夏草的生长产生明显的抑制作用。
IAA属于生长素类激素,能够活化细胞质膜上的质子泵,将细胞质中的H+泵到细胞壁内,使细胞壁基质酸化,细胞壁变松弛,可塑性增强,促进细胞的伸长,从而使细胞的体积和重量增加[14]。因此,适宜浓度的IAA可以促进冬虫夏草生物量的增加。然而,不同的生物对生长素类激素具有不同的浓度适应范围,浓度过高会刺激细胞产生乙烯,引起细胞的老化和死亡[15]。所以,过高浓度的IAA会对冬虫夏草的生长产生明显的抑制作用。
2.2不同浓度IAA对冬虫夏草胞外多糖产量的影响
不同浓度IAA作用下得到的胞外粗多糖含量结果如图2所示。
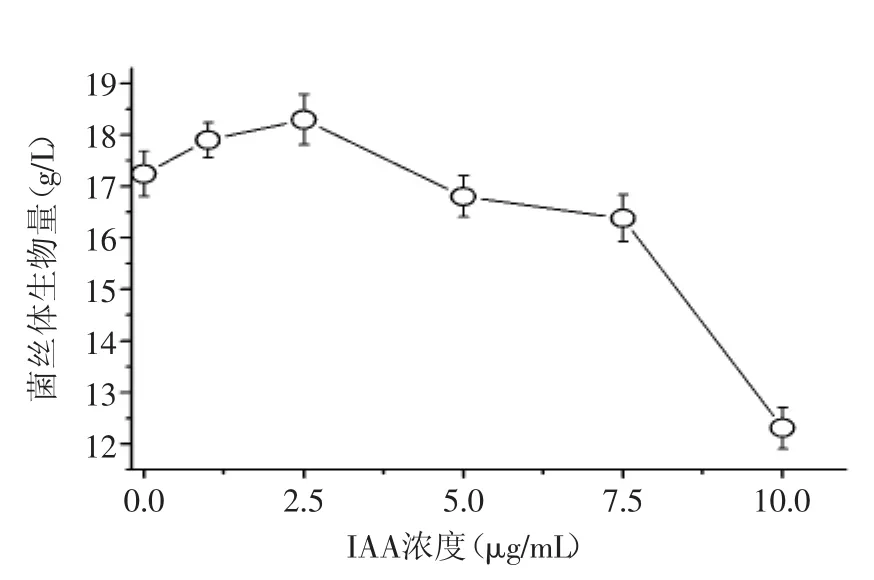
图1 不同IAA浓度对冬虫夏草生物量的影响Fig.1 Effect of different concentration of IAA on mycelial biomass

图2 不同IAA浓度对冬虫夏草胞外多糖产量的影响Fig.2 Effect of different concentration of IAA on EPS production
由图2可知,在0~10μg/mL范围内,随着IAA浓度的增加,冬虫夏草胞外多糖产量呈现出先增后降的变化趋势,在2.5μg/mL时胞外多糖产量达到最大,为2.9g/L,较不添加IAA增加了15.4%;同时,在0~10μg/mL范围内,较不添加IAA的空白组,胞外多糖的产量均有不同程度的提高。结果表明:在0~10μg/mL范围内,IAA可以促进冬虫夏草胞外多糖产量的提高,并且随IAA浓度的增加还会呈现出先增后减的变化趋势。
一方面,IAA可以通过Jak2、IRS、PI3K通道作用于脂肪细胞,促进葡萄糖转运,加速葡萄糖向糖原的转化[16],从而提高了冬虫夏草多糖的产量;另一方面,IAA可以使细胞壁软化和松弛,体积增大,细胞的可塑性和渗透性增强,导致了冬虫夏草胞外多糖的分泌量增加[14,17-18]。同时,过高浓度的IAA会刺激细胞产生乙烯,使细胞老化死亡[15],使多糖的产量降低,胞外多糖分泌量相应减少,因而冬虫夏草胞外多糖的产量会随IAA浓度的增加而呈现先增后降的趋势。
2.3IAA对冬虫夏草胞外多糖羟基自由基清除能力的影响
不同浓度IAA作用下,发酵所得的冬虫夏草胞外多糖羟自由基清除能力结果如图3所示。

图3 冬虫夏草胞外多糖的羟基自由基清除能力Fig.3 The scavenging effect on Hydroxyl radical of EPS from Cordyceps sinensis
由图3可知,不同浓度IAA作用下得到的冬虫夏草胞外多糖对羟基自由基的清除能力均随多糖浓度的增加而增强;在相同多糖浓度下,随着IAA浓度的增加,胞外多糖对羟基自由基的清除能力呈下降趋势,但其清除能力较不添加IAA的空白组更强。当IAA浓度为1.0μg/mL时,冬虫夏草胞外多糖对羟基自由基的清除能力最强,在多糖浓度为2.0mg/mL时清除率可达58.9%。结果表明,IAA可以提高冬虫夏草胞外多糖的羟基自由基清除能力,但其清除能力随IAA浓度的增加而呈现下降趋势。
2.4IAA对冬虫夏草胞外多糖DPPH清除能力的影响
不同浓度IAA作用下,发酵所得的冬虫夏草胞外多糖DPPH清除能力结果如图4所示。

图4 冬虫夏草胞外多糖的DPPH清除能力Fig.4 The scavenging effect on DPPH of EPS from Cordyceps sinensis
由图4可知,不同浓度IAA作用下得到的冬虫夏草胞外多糖对DPPH的清除能力均随多糖浓度的增加而增强;在相同多糖浓度下,随着IAA浓度的增加,胞外多糖对DPPH的清除能力呈现出先增后降的变化趋势。当IAA浓度为2.5μg/mL时,冬虫夏草胞外多糖对DPPH的清除能力最强,在多糖浓度为1.0mg/mL时清除率可达56.6%。结果表明,添加适当浓度的IAA可以提高冬虫夏草胞外多糖的DPPH清除能力,并且随IAA浓度的增加还会呈现出先增后降的变化趋势。
3 结论
适宜浓度的IAA可以提高冬虫夏草的生物量和胞外多糖产量,并且随着IAA浓度的增加还会呈现先增后降的变化趋势,但过高浓度的IAA会对冬虫夏草的生长产生明显的抑制作用。其中,当IAA浓度为2.5μg/mL时,冬虫夏草的生物量和胞外多糖产量均达到最高,分别为18.3g/L和2.9g/L,较不添加IAA分别增加了6.3%和15.4%。
IAA可以提高冬虫夏草胞外多糖的抗氧化能力。随着IAA浓度的增加,冬虫夏草胞外多糖对羟基自由基的清除能力呈下降趋势,而对于DPPH的清除能力却呈先增后降的变化趋势。当IAA浓度为1.0μg/mL和2.5μg/mL时,胞外多糖对羟基自由基和DPPH的清除率分别达到最高,为58.9%和56.6%。冬虫夏草胞外多糖抗氧化能力的提高,可能是由于IAA的加入改变了多糖的合成方式,导致多糖的空间结构改变或抗氧化活性高的多糖所占比例增加,从而使冬虫夏草胞外多糖表现出更好的抗氧化性能,但具体原因有待进一步的研究分析。
[1]卯晓岚.中国蕈菌[M].北京:科学出版社,2009:710-716.
[2]程维蓉,段丽红,郑必胜.冬虫夏草及其多糖的研究与应用进展[J].现代食品科技,2006,22(4):284-289.
[3]赵秋容,李建平,吴迪,等.冬虫夏草中多糖提取、纯化及抗氧化性能的研究[J].中国农学通报,2012,28(15):238-242.
[4]吴仪洛(清).本草从新[M].天津:天津科学技术出版社,2003:25-27.
[5]刘高强,王晓玲,杨青,等.冬虫夏草化学成分及其药理活性研究[J].营养研究,2007(1):202-206.
[6]魏原芝,陈书进,赵盼,等.植物激素对杏鲍菇菌丝生长及酶活性的影响[J].安徽农业科学,2012,40(29):14179-4180.
[7]上官端琳,吴素蕊,赵天瑞,等.α-萘乙酸对黑脉羊肚菌生物量、胞内多糖含量及多糖抗氧化性能的影响[J].食品与发酵工业,2013,39(11):1-4.
[8]王谦,刘敏,徐啸晨,等.外源激素对金福菇菌丝营养生长的影响[J].河北大学学报:自然科学版,2012,32(3):286-290.
[9]马荣山,凌岚,方蕊,等.草原白磨菌种分离及菌丝体生长因子实验[J].食品科技,2011,36(4):18-21.
[10]朱朝阳,刘高强,旷思敏,等.冬虫夏草真菌产胞外多糖发酵条件的优化[J].食品科技,2013,38(5):18-21.
[11]Dubois M,Gilles KA,Hamilton JK,et al.Colorimetric method for determination of sugars and related substances[J].Analytical Chemistry,1956(28):350-356.
[12]Smirnoff N,Cumbes Q.Hydroxyl radical scavenging activity of compatible solutes[J].Phytochemistry,1989,28(4):1057-1060.
[13]Blois MS.Antioxidant determination by the use of a stable free radical[J].Nature,2002,26:1199-1200.
[14]孟庆杰,王光全.植物激素及其在农业生产中的应用[J].河南农业科学,2006(4):9-12.
[15]王婉婉,季静.植物激素在月季中的作用及其应用[J].天津农业科学,2009,15(3):29-31.
[16]高雪,徐尚忠,张英汉.生长激素作用机理的研究进展[J].黄牛杂志,2003,29(1):50-53.
[17]崔凤国.生长素的代谢及作用机理[J].生物学教学,2011,36(8):71-72.
[18]毛丽君,林位夫.植物生长素在农业中的应用[J].河北农业科学,2008,12(2):80-84.
Effects of indole acetic acids(IAA)on biomass,extracellular polysaccharide production and antioxidant activity of Cordyceps sinensis(Berk.)Sacc.
ZHANG Zhi-hong1,WU Su-rui2,TAI Li-mei2,FAN Jian1,ZHAO Tian-rui1,*
(1.Yunnan Institute of Food Safety,Kunming University of Science and Technology,Kunming 650500,China;2.Kunming Edible Fungi Institute of All China Federation of Supply of Marketing Cooperative,Kunming 650223,China)
The effect of indole acetic acids(IAA)on biomass,extracellular polysaccharide(EPS)production and antioxidant activity of Cordyceps sinensis were studied in the present work.The results showed that the reasonable concentration of IAA could increase biomass and EPS production of Cordyceps sinensis.Biomass and EPS production were increased first and then decreased with the increase of IAA.Both of biomass and EPS production reached the highest content at 2.5μg/mL IAA,namely 18.3g/L and 2.9g/L,increased 6.3%and 15.4%comparing no IAA,respectively.The appropriate concentration of IAA could also improve the antioxidant activity of EPS.The strongest scavenging effect on hydroxyl radical and DPPH of EPS were 58.9%and 56.6% at the concentrations of 1.0μg/mL and 2.5μg/mL,respectively.
Cordyceps sinensis;IAA;biomass;EPS;antioxidant activity
TS201.1
A
1002-0306(2015)12-0181-04
10.13386/j.issn1002-0306.2015.12.029
2014-08-27
张志红(1989-),男,硕士研究生,研究方向:发酵工程。
赵天瑞(1964-),男,大学本科,副教授,主要从事食品科学与工程方面的研究。
国家科技支撑计划课题(2013BAD16B01)。
猜你喜欢
医疗装备(2023年3期)2023-03-22
山东第一医科大学(山东省医学科学院)学报(2021年7期)2021-10-13
昆明医科大学学报(2021年4期)2021-07-23
昆明医科大学学报(2020年12期)2021-01-26
发明与创新·小学生(2021年12期)2021-01-06
军事文摘(2020年22期)2021-01-04
山东化工(2019年11期)2019-06-26
国际呼吸杂志(2019年1期)2019-03-08
环境保护与循环经济(2017年3期)2017-09-26
汉语世界(2016年3期)2016-11-16
