
脂氧合酶(LOX)在脂肪酸氧化中的作用研究进展
2015-10-21 08:58:09曲清莉傅茂润代红飞
食品研究与开发 2015年10期
曲清莉,傅茂润,代红飞
(齐鲁工业大学食品与生物工程学院,山东济南250353)
脂氧合酶(LOX)在脂肪酸氧化中的作用研究进展
曲清莉,傅茂润*,代红飞
(齐鲁工业大学食品与生物工程学院,山东济南250353)
脂氧合酶在脂肪酸氧化中具有重要作用,一直是人们的研究热点。本文系统论述了脂氧合酶尤其是其活性中心的结构,分析和比较了几种LOX作用机制的优缺点,如自由基理论、质子受体理论、氢转移理论等;从分子生物学角度研究了脂氧合酶的基因表达,并对其功能进行了分析;最后从温度、pH、底物、水分及抑制剂等方面讨论了脂氧合酶的调控技术研究进展。
脂氧合酶;作用机制;基因克隆;调控
脂肪氧化包括自动氧化、光氧化和酶促氧化:自动氧化是活化的含烯底物与基态氧3O2发生的游离基反应,包括链引发、链传递和链终止3个阶段;光氧化是不饱和双键与单线态氧(1O2)直接发生的氧化反应;酶促氧化是由脂氧合酶参加的氧化反应,是本文讲述的重点。脂氧合酶(lipoxygenase,LOX,EC1.13.11.12)又名脂肪氧化酶、脂肪加氧酶或类胡萝卜素氧化酶。脂氧合酶在生物体内专一催化含有顺,顺-1,4-戊二烯结构的多元不饱和脂肪酸的加氧反应,生成具有共轭双键的多元不饱和酸的氢过氧化物[1]。氢过氧化物不稳定,当体系中的浓度增至一定程度时,就开始分解,进一步生成醛、醇或酮等具有令人不愉快的哈喇味的小分子,导致油脂酸败。油脂氧化产生的小分子化合物可进一步发生聚合反应,生成结构复杂的聚合物。
本文系统论述了脂氧合酶的结构和作用机制,从分子生物学角度研究了脂氧合酶的基因表达,并对其功能进行了分析,并从温度、pH、底物、水分及抑制剂等方面讨论了脂氧合酶的调控进展,以期为后续脂氧合酶和脂肪酸氧化的研究提供有益参考。
1脂氧合酶的结构
脂氧合酶的来源不同,其氨基酸的组成不同,植物脂肪氧合酶的氨基酸残基比动物脂肪氧合酶多25%[2],如大豆脂肪氧合酶(LOX-1)的一条蛋白单链具有839个氨基酸残基。植物脂氧合酶系包括约(25~30)×103个单聚体蛋白、参与细胞膜或底物的结合的N端结构域,和约(55~65)×103个单聚体蛋白、包含α-螺旋,锚定酶的催化位点的C端结构域。
脂氧合酶的功能域是由金属离子和氨基酸残基组成的,植物脂氧合酶的金属离子是铁,真菌脂氧合酶是锰。植物脂氧合酶的氨基酸残基一般是三个组氨酸、一个天冬氨酸和一个异亮氨酸,哺乳动物脂氧合酶系是四个组氨酸和一个异亮氨酸[3]。植物脂氧合酶催化中心与铁离子有着密切的关系,Tomchick等通过对野生型大豆脂氧合酶的4种突变体Q495E,Q495A,Q697N,Q697E进行动力学及结构研究,发现铁第二配位层的改变会使其氢键重排,确定酶作用底物与Gln495相互作用的位置,从而影响酶活力[4]。
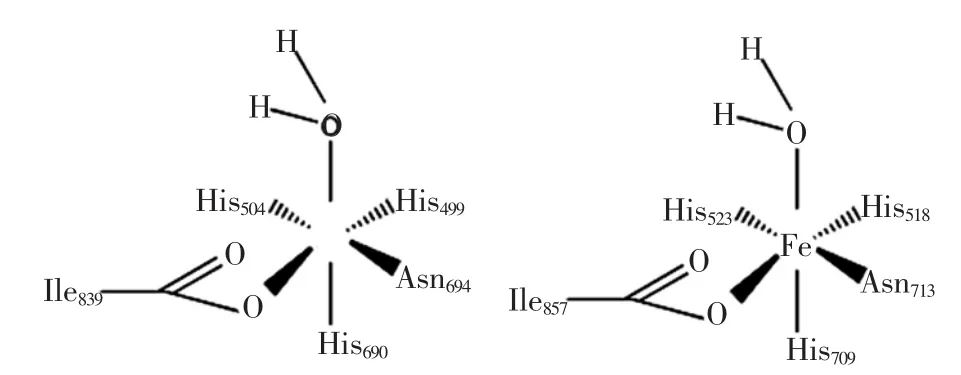
图1 LOX-1和LOX-3的铁活性中心X-射线晶体结构Fig.1X-ray crystal structure of iron active center of LOX-1 and LOX-3
2脂氧合酶的作用机制

表1 脂氧合酶催化反应的几种不同机制Table 1Several different mechanisms of reaction catalyzed bylipoxygenase

续表1脂氧合酶催化反应的几种不同机制Continue table 1Several different mechanisms of reaction catalyzed by lipoxygenase
3脂氧合酶的分子生物学研究
3.1脂氧合酶的基因克隆与序列分析
自20世纪80年代利用分子克隆技术首次确定了大豆LOX-1的完整氨基酸序列以来[13],研究者已经从拟南芥、水稻、小麦、大麦、番茄、马铃薯、烟草、猕猴桃等植物中分别克隆到了脂氧合酶基因家族成员。其中从拟南芥中克隆到至少6个LOX家族成员,均有不同的功能表现。根据GeneBank公布的氨基酸序列,我们构建了一个由3个分支组成的植物脂氧合酶基因系统进化树[14]。
朱凤云等以油桐种子的转录组数据库中的脂氧合酶基因的部分cDNA序列为基础,采用RT-PCR和RACE技术从油桐种子中克隆到此基因的全长cDNA,长度为2 690bp,编码797个氨基酸,起始密码子为ATG,终止密码子为TAA,并其进行了序列分析,发现油桐LOX氨基酸序列与茶树、葡萄、辣椒、马铃薯的同源性高达99%,并推测其属于9-LOX[15]。
张波等利用猕猴桃EST库及相关生物信息学手段,从果肉组织克隆了脂氧合酶基因家族成员AdLox1-6。AdLox1、AdLox2、AdLox3和AdLox4为全长cDNA,编码区长度分别为2 739、2 595、2 739和2 709 bp,AdLox5和AdLox6为cDNA片段,长度为1 359 bp和1 557 bp。其中猕猴桃AdLox1、AdLox3、AdLox4和AdLox6属13-LOX类型,AdLox2和AdLox5属于9-LOX类型,与已知功能的番茄、烟草和马铃薯等植物LOX基因家族成员具有序列同源性[16]。
3.2脂氧合酶的基因表达与功能分析
脂氧合酶基因家族根据其催化亚油酸中加氧位置的不同,分为9-LOX、13-LOX两种类型[17],9-LOX和13-LOX在序列长度上有差异,13-LOX在N端含有额外约60个氨基酸残基[18]。它们有很多同工酶,在同一物种的不同发育阶段、不同的胁迫作用,可诱导相应基因的表达。
植物脂氧合酶基因的表达受到发育信号的调节。脂氧合酶基因的表达属于信号转导途径中的早期事件,能对外界以及胞内信号迅速作出反应。特异的LOX基因在大麦、黄瓜和大豆种子萌发的早期阶段被诱导。环境胁迫因素也可诱导植物脂氧合酶基因的表达[19]。通过研究拟南芥、马铃薯[20]等植物,发现机械损伤和食草动物进食能够诱导LOX基因的表达。猕猴桃中6种脂氧合酶基因在果实成熟期间有不同的基因表达,AdLox1和AdLox5具有乙烯依赖性,而AdLox2、AdLox3、AdLox4和AdLox6对果实成熟有负调节作用。
1.1.3 试验用虾 2018年4月从距武汉农业气象试验基地30 min车程的虾苗养殖基地购入平均质量3~5 g的虾苗,每个网箱虾苗投放数量均为20尾。
研究发现,抑制脂氧合酶基因表达的拟南芥转基因植株,叶片组织茉莉酸含量显著低于野生型[21];番茄脂氧合酶基因的反义表达,会显著减少果实香气物质含量[22];转基因抑制马铃薯脂氧合酶基因转录会显著减少叶片己醛积累[23];降低马铃薯脂氧合酶基因表达可以导致块茎变小和产量下降[24]。
脂氧合酶在真核生物中参与不饱和脂肪酸的代谢,是催化脂质降解的关键酶,也是茉莉酸合成途径的关键酶[25],植物膜脂中富含的亚油酸和亚麻酸是其主要的反应底物[26],还可能参与了植物挥发性化合物如己醛和己烯醛的合成。研究表明,脂氧合酶在植物的生长、发育、衰老以及环境胁迫反应中起重要调节作用,并与果实乙烯合成、后熟软化和香气形成等进程关系密切。脂氧合酶催化反应生成的超氧自由基和氢过氧化物均可能参与了乙烯的生物合成,并由此调控果实软化[27]。
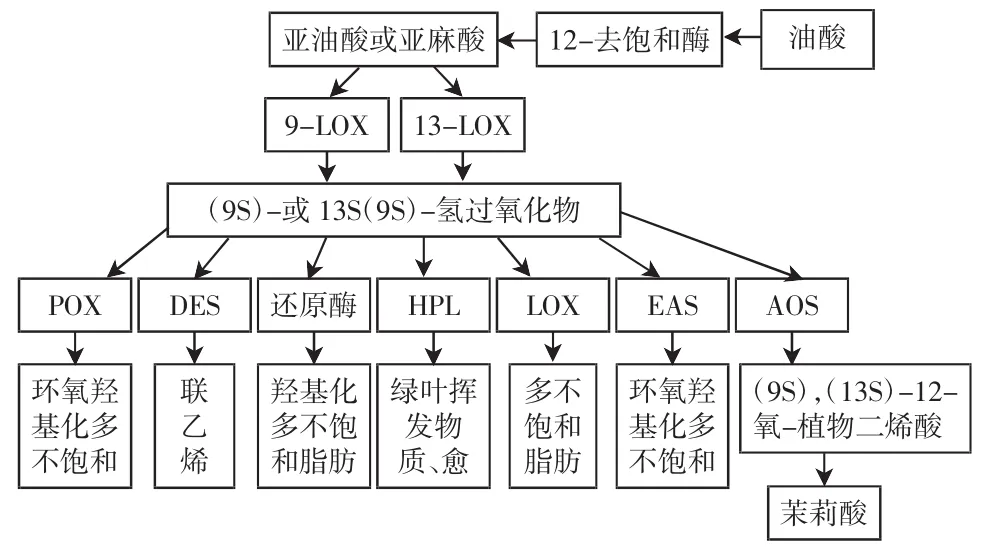
图2 植物脂氧合酶的代谢途径Fig.2Metabolic pathways of plant lipoxygenase
4脂氧合酶的调控
脂肪氧合酶会产生两种有害的副作用:一是造成有营养价值的多不饱和脂肪酸损失,二是产生导致酸败的氧化产物,在加工保藏期间产生不良的风味或导致食品在其他方面的质量的下降,因此,很多情况下,采用各种方法抑制脂氧合酶活性是十分必要的。
另一方面,脂氧合酶在面包加工过程能够漂白胡萝卜素色素,在面团混合期间释放结合态脂肪,形成高消耗功率下面团的流变学行为,在面团混合期间形成游离自由基,对面团醒发有重要影响[28]。在烟叶烘烤过程中,脂氧合酶能充分降解叶绿素和类胡萝卜素,对烟叶质体色素的降解和烟叶变黄具有促进作用,减少活性氧积累,使烟叶生命活动时间更长,进而产生更多的香气物质,相应降低有害成分[29]。因此,在这些加工过程中,如何保持或提高脂氧合酶活性是需要我们考虑的。
4.1温度调控
大量研究证实,多数脂氧合酶最适反应温度在30℃~45℃之间[30]。低于最适反应温度时处理脂氧合酶,其自身温度随处理时间的延长不断增加,反应物分子动能随之增加,酶活性增加,低温24 min内处理不会出现热失活现象。高于最适温度时,均出现酶活下降趋势,且温度越高下降越快,因为高温加速酶蛋白变性,而这种变性随时间累积而加强[31]。常温下植物果实软化衰老加快,脂氧合酶活性较高,在贮藏前期,低温可显著抑制脂氧合酶活性。田寿乐等分别在0℃和20℃下贮藏鲜枣果实,30 d内0℃贮藏酶活性只有20℃的一半,有效延缓了果实软化[32]。王贵禧等将大久保桃采后先在8℃下经过9 d冷锻炼,然后再在0℃下冷藏,脂氧合酶活性得到有效抑制,桃的硬度保持较好,出库后能够正常后熟软化,在预防了冷害的同时,较好地保持了大久保桃的品质[33]。Lopez等发现10℃贮藏核桃可以很好地抑制脂氧合酶活性,使得核桃品质可以保持最少一年[34]。钟诚提出初榨橄榄油在贮藏过程中会因脂氧合酶作用而产生较大的风味变化,引起常见的品质缺陷,亦应低温贮藏[35]。宋虎卫等发现油柰果实可于(3±1)℃贮藏,脂氧合酶活性仅为常温贮藏是的一半,贮藏期可达77 d以上[36]。加工豆奶时,将未浸泡的脱壳大豆在加热到80℃~100℃的热水中研磨10 min使脂氧合酶失活,消除了不良风味。
4.2pH调控
通常在自然生长环境中,生物体要面临许多环境胁迫因素如酸碱度的改变,会调控LOX基因的表达,使其发挥不同的生理功能,如改变脂氧合酶的作用位置特异性,在pH 6.0时,大豆LOX-1氧化亚油酸产生12%的9(S)-HPOD,但在pH 9.0时,只产生13(S)-HPOD[37]。在极酸和极碱条件下,几乎检测不到脂氧合酶的活性;在pH6.0~9.0之间一般会出现最适pH,由于脂氧合酶是一个多基因家族,存在多种同工酶,不同脂氧合酶的最适pH存在差异,有的还会出现2个活性峰值,如Mahanta等研究发现,毛尖茶叶中脂氧合酶在pH7.5和pH9.0时有活性峰[38],刘夫国等证实糯玉米中脂氧合酶在pH6.0和pH8.0时有活性峰[31]。姜爱丽等通过高CO2对采后蓝莓进行冲击处理,显著降低其pH至2.7,10 d内处理后蓝莓的酶活性仅是未处理的1/3,有效控制了腐烂率,保持了较高的果实硬度,并使有效贮藏期延长到50 d左右[39]。将食品材料调节到pH偏酸性再热处理,也是使脂肪氧合酶失活的有效方法,如将大豆在pH3.88和水一起研磨,然后再烧煮,能使脂氧合酶变性,消除不良风味。
4.3底物浓度调控
通过改变底物多元不饱和脂肪酸浓度,可以很方便的调控脂氧合酶。在一定底物浓度范围内,随底物浓度增加脂氧合酶活性提高,而超过一定范围,底物浓度增加而脂氧合酶活性提高不显著,可能是由于反应体系中底物浓度过高时,有一部分在空气中自动氧化为羟基过氧化物,当羟基过氧化物达到一定浓度后,抑制了脂氧合酶的活性[40]。
4.4氧分压调控
不同氧分压条件下,脂氧合酶活性会有很大变化。脂氧合酶的活性在金针菇贮藏的前7天左右呈现对数性增长,随着时间的延长而逐渐下降;80%氧分压时,在整个贮藏过程中脂氧合酶活性明显高于其他试验组,而20%~50%低氧分压时,脂氧合酶活性较低,且下降速度更为明显[41]。脂氧合酶催化多不饱和脂肪酸反应通常在需氧条件下进行,但在缺氧的情况下脂氧合酶也有活性,被认为可以催化从脂肪酸提取氢自由基,然后启动进一步的自由基反应[42]。还有实验证明,在低氧浓度下大豆脂氧合酶会丧失位置特异性,产生相等量的9(S)-HPOD和13(S)-HPOD混合物,而在正常情况下会产生95%的13(S)-HPOD。Song等将猕猴桃用氮气处理(缺氧处理)6 h可以降低LOX酶活性,使其在1℃条件下14 d之内保持较高的硬度,并延缓贮藏期内果实脂质过氧化作用[43]。孙谷畴等在空气和高浓度氧(45%)条件下贮藏荔枝2 d,果皮和果肉中脂氧合酶活性增高,而在低浓度(5.3%)氧下则无增高,有效减缓氧化和过氧化作用并延缓果皮褐变[44]。
4.5水分含量调控
在一定程度上,水分含量的增加会提高脂氧合酶的活性,可能由于脂氧合酶在水油界面上发生作用,高的水分含量加大了水油界面,从而使酶活较大。据郝利平等实验分析表明,核桃在7℃下贮藏6个月内,水分含量8%比3%的脂氧合酶活性大1.6倍左右[45]。我国对需要进行贮藏的核桃仁含水量一般要求为6%~8%以下,以减弱脂氧合酶活性,延长贮藏时间[46]。为了延缓谷物陈化,一般要求贮藏条件为低温干燥,以减少脂氧合酶催化脂肪过氧化。李玉红等发现菜豆种子在含水量为3.8%时,脂氧合酶活性显著低于其它含水量的种子,可以提高其抗老化劣变能力和发芽率,表现出较高的生活力[47]。
4.6脂氧合酶抑制剂调控
目前,脂氧合酶抑制剂有异羟肟酸类、去甲二氢愈创木酸类、黄酮类和脂氧合酶激活蛋白抑制剂等,根据作用方式一般划分为氧化还原抑制剂、非氧化还原抑制剂、铁离子配体抑制剂和FLAP抑制剂等[48]。
研究较多的黄酮类抑制剂是亲脂还原剂,它作用于LOX活化部位的铁离子,把+3价活性状态的铁离子还原为+2价的非活性状态,干扰了铁离子催化循环,从而阻止LOX的激活[49]。
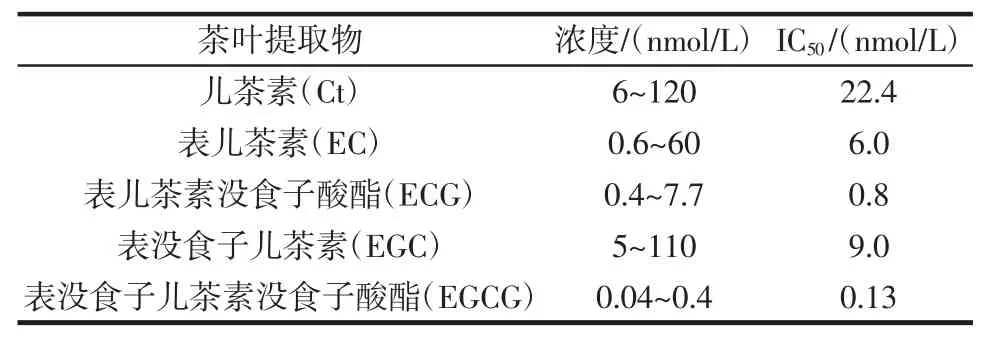
表2 茶叶提取物对脂氧合酶的抑制效果[50]Table 2Inhibitory effect of tea extracts on Lipoxygenase
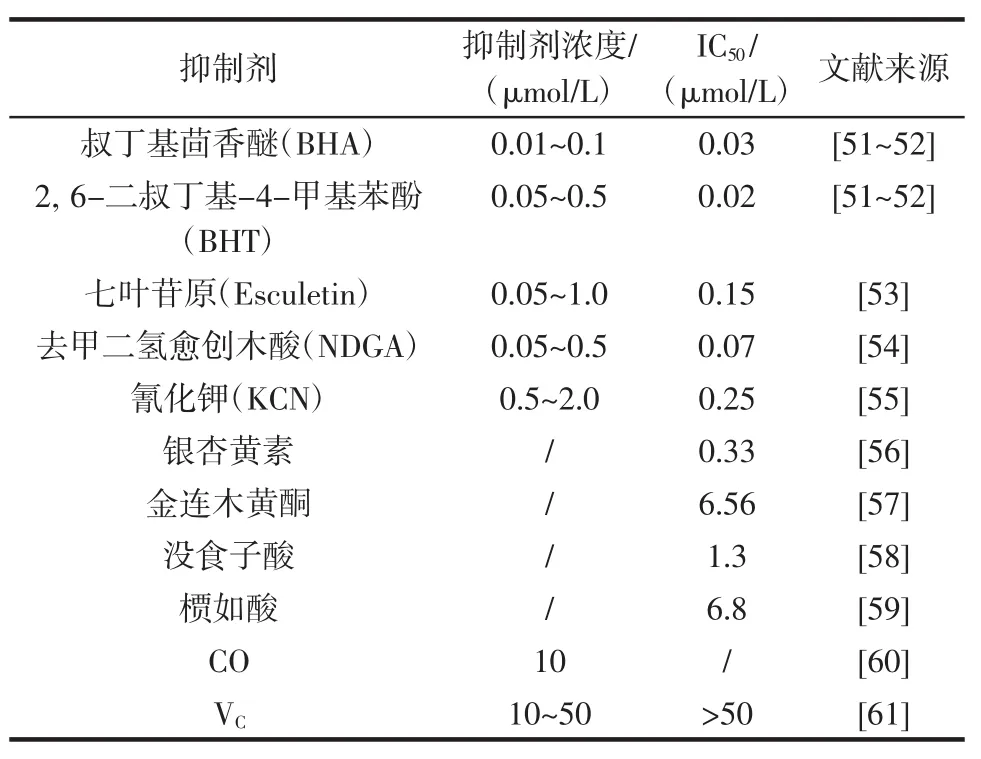
表3 其它物质对脂氧合酶的抑制效果Table 3Inhibitory effect of other substances on Lipoxygenase
5结论
脂肪氧化与我们生活息息相关,人们对脂肪氧化和脂氧合酶的研究也在一直进行,且取得了很大成就。脂氧合酶的结构及其反应机理逐渐成熟,脂氧合酶的调控、分子生物遗传学等方面也有很大进展。
但脂氧合酶研究中仍然存在很多问题,脂氧合酶促进脂肪氧化的反应机理还没有统一的观点,对脂质代谢的调控作用研究不多,9-LOX和13-LOX生理功能及其基因表达的差异性尚未有系统的研究,其生化途径,尤其是9-LOX途径没有明确。由于脂氧合酶有多种同工酶,不同来源的酶在相应物种中具体的生理作用、作用机理及在信号途径中扮演的角色有待我们进一步的研究。
[1]熊卫东,章银良,马庆一,等.脂氧合酶活性抑制剂的筛选及应用保鲜的研究[J].食品科技,2004(9):84-87
[2]蔡琨,方云,夏咏梅.植物脂肪氧合酶的研究进展[J].现代化工,2003,23(增刊):23-27
[3]李兴军.脂氧合酶分子结构与反应机制[J].粮食科技与经济,2012,37(4):57-60.
[4]Tomchick D R,Phan P,Cymborowski M,et al.Structural and functional characterization of second-coordination sphere mutants of soybean lipoxygenase-1[J].Biochemistry,2001,40(25):7509-7517
[5]Minor W,Steczko J,Stec B,et al.Crystal structure of soybean lipoxygenase L-l at 1.4 angstrom resolution[J].Biochemistry,1996,35(33):10687-10701
[6]Solomon E I,Brunold T C,Davis M I,et al.Geometric and electronic structure/function correlations in non-heme iron enzymes[J].Chemical reviews,2000,100(1):235-350
[7]Tappel A L,Boyer P D,Lundberg W O.The reaction mechanism of soy bean lipoxidase[J].JournalofBiological Chemistry,1952,199(1):267-281
[8]Borowski T,Broclawik E.Catalytic reaction mechanism of lipoxygenase.A density functional theory study[J].The Journal of Physical Chemistry B,2003,107(19):4639-4646
[9]Corey E J,Nagata R.Evidence in favor of an organoiron-mediated pathway for lipoxygenation of fatty acids by soybean lipoxygenase[J].Journal of the American Chemical Society,1987,109(26):8107-8108
[10]Nelson M J,Seitz S P.The structure and function of lipoxygenase[J]. Current opinion in structural biology,1994,4(6):878-884
[11]Andreou A,Feussner I.Lipoxygenases-structure and reaction mechanism[J].Phytochemistry,2009,70(13):1504-1510
[12]Hammes-Schiffer S.Hydrogen tunneling and protein motion in enzyme reactions[J].Accounts of chemical research,2006,39(2):93-100
[13]da Silva Fortunato F,de Almeida Oliveira M G,Brumano M H N,et al.Lipoxygenase-induced defense of soybean varieties to the attack of the velvetbean caterpillar(Anticarsia gemmatalis Hübner[J]. Journal of Pest Science,2007,80(4):241-247
[14]张波,李鲜,陈昆松.脂氧合酶基因家族成员与果实成熟衰老研究进展[J].园艺学报,2007,34(1):245-250
[15]朱凤云,陈鸿鹏,谭晓风,等.油茶脂氧合酶基因全长cDNA的克隆与序列分析[J].中南林业科技大学学报,2012,32(10):45-51
[16]张波,李鲜,陈昆松.基于EST库的猕猴桃脂氧合酶基因家族成员的克隆[J].园艺学报,2008,35(3):337-342
[17]范海兰,洪伟,吴承祯,等.土壤水分胁迫对短葶山麦冬光响应特性的影响[J].福建林学院学报,2011,31(3):193-197
[18]朱林,许兴.植物水分利用效率的影响因子研究综述[J].干旱地区农业研究,2005,23(6):204-209
[19]Grechkin A.Recent developments in biochemistry of the plant lipoxygenase pathway[J].Progress in Lipid Research,1998,37(5):317-352
[20]Royo J,León J,Vancanneyt G,et al.Antisense-mediated depletion of a potato lipoxygenase reduces wound induction of proteinase inhibitors and increases weight gain of insect pests[J].Proceedings of the National Academy of Sciences,1999,96(3):1146-1151
[21]Bell E,Creelman R A,Mullet J E.A chloroplast lipoxygenase is required for wound-induced jasmonic acid accumulation in Arabidopsis[J].Proceedings of the National Academy of Sciences,1995,92(19):8675-8679
[22]Chen G,Hackett R,Walker D,et al.Identification of a specific isoform of tomato lipoxygenase(TomloxC)involved in the generation of fatty acid-derived flavor compounds[J].Plant Physiology,2004,136(1):2641-2651
[23]León J,Royo J,Vancanneyt G,et al.Lipoxygenase H1 gene silencing reveals a specific role in supplying fatty acid hydroperoxides for aliphatic aldehyde production[J].Journal of Biological Chemistry,2002,277(1):416-423
[24]Kolomiets M V,Hannapel D J,Chen H,et al.Lipoxygenase is involved in the control of potato tuber development[J].The Plant Cell Online,2001,13(3):613-626
[25]汪仁,沈文飚,江玲,等.水稻种子脂氧合酶基因OsLOX1的原核表达、纯化及鉴定[J].中国水稻科学,2008,22(2):118-124
[26]胡廷章,胡宗利,屈霄霄,等.植物脂肪氧化酶的研究进展[J].生物工程学报,2009,25(1):1-9
[27]何全光,邝健飞,陈建业,等.脂氧合酶在香蕉果实成熟过程中的作用[J].中国农业科学,2011,44(1):118-124
[28]李兴军.脂氧合酶及相关酶在面包加工中的作用[J].粮食科技与经济,2011,36(5):36-40
[29]赵华武,贺帆,石盼盼,等.密集烘烤过程中不同前处理烟叶生理生化变化研究[J].中国农业大学学报,2012,17(3):101-106
[30]李铭韧,史国安.牡丹花瓣脂氧合酶测定方法的研究[J].山东农业科学,2010(9):91-94
[31]刘夫国,金邦荃,牛丽影,等.鲜食玉米脂氧合酶的酶学性质[J].食品科学,2013,34(3):198-201
[32]田寿乐,周俊义.不同贮藏温度与鲜枣果实中保护酶及脂氧合酶活性变化的关系[J].河北农业大学学报,2006,29(1):46-49
[33]王贵禧,王友升,梁丽松.不同贮藏温度模式下大久保桃果实冷害及其品质劣变研究[J].林业科学研究,2005,18(2):114-119
[34]Lopez A,Pique M T,Romero A,et al.Influence of cold-storage conditions on the quality of unshelled walnuts[J].International journal of refrigeration,1995,18(8):544-549
[35]钟诚,薛雅琳,王兴国,等.初榨橄榄油风味化合物研究进展[J].中国油脂,2013,38(8):89-92
[36]宋虎卫,潘东明.不同贮藏温度下油柰采后生理变化及其与耐贮性的关系[J].现代农业科技,2009,21:63-65
[37]李靖,马长乐.植物脂氧合酶研究进展[J].生物学杂志,2007,24(6):5-8
[38]Mahanta P K,Tamuli P,Bhuyan L P.Changes of fatty acid contents,lipoxygenase activities,and volatiles during black tea manufacture[J].Journal of agricultural and food chemistry,1993,41(10):1677-1683
[39]姜爱丽,孟宪军,胡文忠,等.高CO2冲击处理对采后蓝莓生理代谢及品质的影响[J].农业工程学报,2011,27(3):362-368
[40]Perraud X,Kermasha S,Bisanowski B.Characterization of a lipoxygenase extract from Geotrichum candidum[J].Process Biochemistry,1999,34(8):819-827
[41]王成涛,王昌涛,赵磊.不同氧分压条件下金针菇贮藏保鲜相关酶活性的研究[J].食品科学,2011,32(8):300-304
[42]李兴军.谷物脂氧合酶的生理生化特性[J].粮食科技与经济,2011,36(3):34-37
[43]Song L,Gao H,Chen H,et al.短期氮处理(缺氧处理)对采后猕猴桃贮藏过程中抗氧化能力和质膜完整性的影响[J].保鲜与加工,2010(6):55-55
[44]孙谷畴,林植芳,林桂珠,等.不同浓度氧贮藏荔枝的几种酶活性变化[J].广西植物,1992,12(1):36-40
[45]郝利平,杨剑婷.贮藏因素对核桃脂肪酶活性与油脂酸价的影响[J].农业工程学报,2005,21(5):170-172
[46]李喜宏,陈丽.实用果蔬保鲜技术[M].科学技术文献出版社,2000:345-346
[47]李玉红,陈鹏,唐爱均,等.不同含水量菜豆种子老化过程中生理特性的研究[J].园艺学报,2005,32(5):908-910
[48]朱宏翔,胡建安.脂氧合酶抑制剂的研究进展[J].中南大学学报,2008,33(6):541-547
[49]黄云,胡建安.黄酮类化合物对脂氧合酶活性的影响及其生物学作用[J].中国药理学与毒理学杂志,2009,23(6):490-496
[50]Banerjee S.Inhibition of mackerel(Scomber scombrus)muscle lipoxygenase by green tea polyphenols[J].Food Research Internationa,2006,39(4):486-491
[51]何雄,薛长湖,杨文鸽,等.罗非鱼鳃组织中脂肪氧合酶的性质研究[J].水产科学,2005,24(7):15-19
[52]Barlow S M.Toxicological aspects of antioxidants used as food additives[M].Food antioxidants.Springer Netherlands,1990:253-307
[53]Mohri S,Tokuori K,Endo Y,et al.Prooxidant activities in fish skin extracts and effects of some antioxidants and inhibitors on their activities[J].Fisheries Science,1999,65:269-273
[54]Lambert J,Dorr R,Timmermann B.Nordihydroguaiaretic acid:a review of its numerous and varied biological activities[J].Pharmaceutical biology,2004,42(2):149-158
[55]Banerjee S,Khokhar S,Apenten R K O.Characterization of lipoxygenase from mackerel(Scomber scombrus)muscle[J].Journal of Food Biochemistry,2002,26(1):1-19
[56]Son J K,Son M J,Lee E,et al.Ginkgetin,a Biflavone from Ginko biloba leaves,inhibits cyclooxygenases-2 and 5-lipoxygenase in mouse bone marrow-derived mast cells[J].Biological and Pharmaceutical Bulletin,2005,28(12):2181-2184
[57]Son M J,Moon T C,Lee E K,et al.Naturally occurring biflavonoid,ochnaflavone,inhibits cyclooxygenases-2 and 5-lipoxygenase in mouse bone marrow-derived mast cells[J].Molecular nutrition&food research,2006,29(4):282-286
[58]Ha T J,Nihei K,Kubo I.Lipoxygenase inhibitory activity of octyl gallate[J].Journal of agricultural and food chemistry,2004,52(10):3177-3181
[59]Kubo I,Masuoka N,Ha T J,et al.Antioxidant activity of anacardic acids[J].Food Chemistry,2006,99(3):555-562
[60]李莹.外源CO与乙烯处理在桃果实贮藏过程中对呼吸代谢的影响[J].山西师范大学学报(自然科学版)研究生论文专刊,2013,27:52-55
[61]Saeed S,Howell N.12-Lipoxygenase activity in muscle tissue of Atlantic mackerel(Scomber scombrus)and its prevention by antioxidants[J].Journal of the Science of Food and Agriculture,2001,81(8):745-750
Research Progress of Effect of Lipoxygenase(LOX)on Fatty Acid Oxidation
QU Qing-li,FU Mao-run*,DAI Hong-fei
(College of Food Science and Bioengineering,Qilu University of Technology,Jinan 250353,Shandong,China)
Lipoxygenase is important to fat oxidation and have been a research hotspot.The structure of lipoxygenase especially the active center was elaborated,and several mechanisms of action were compared and analyzed,such as free radical theory,proton acceptor theory,hydrogen transfer theory and so on.Gene expression and function of lipoxygenase from the perspective of molecular biology were analyzed,and the research progress on control techniques of lipoxygenase were expounded,such as the temperature,pH,substrate,water and inhibitor control actions.
lipoxygenase;action mechanism;gene cloning;regulation
10.3969/j.issn.1005-6521.2015.10.037
2014-01-11
山东省优秀中青年科学家奖励基金项目(BS2011NY009)作者简介:曲清莉(1992—),女(汉),在读硕士研究生,研究方向:食品科学基础。
傅茂润(1981—),男,副教授,研究方向:农产品加工和贮藏。
猜你喜欢
世界科学技术-中医药现代化(2021年8期)2021-12-21 07:04:22
疯狂英语·新读写(2021年10期)2021-12-07 02:41:30
中成药(2021年5期)2021-07-21 08:38:32
中华诗词(2020年8期)2020-02-06 09:26:54
奥秘(2019年8期)2019-08-28 01:47:05
上海包装(2019年2期)2019-05-20 09:10:52
中成药(2018年2期)2018-05-09 07:19:49
商周刊(2017年7期)2017-08-22 03:36:21
小猕猴智力画刊(2016年6期)2016-05-14 09:21:40
微生物与感染(2015年1期)2015-02-28 17:42:37
