
入侵植物小飞蓬光合日变化及与环境因子相关性分析
2015-09-10 15:24王晓红纪明山
江苏农业科学 2015年8期
关键词:相关性
王晓红 纪明山


摘要: 为从生理生态角度探讨小飞蓬的光合特性与入侵潜力之间的关系,在小飞蓬生长旺盛期测定其叶片光合日变化进程及与环境因子的相关性。结果表明,小飞蓬叶片净光合速率日变化呈双峰曲线,峰值分别为 28.72、24.36 μmol/(m2·s),存在明显的光合“午休”现象,此时净光合速率为21.22 μmol/(m2·s), 小飞蓬净光合速率的下降主要是由于气孔限制因素所致;相关性分析表明,小飞蓬叶片净光合速率主要受光合有效辐射、空气温度和相对湿度的影响,且各环境因子之间显著相关。将小飞蓬净光合速率与各环境因子做回归分析得出r=0.926,说明小飞蓬净光合速率92.6%的日变化是由生态因子日变化引起的。小飞蓬具有的上述光合特性及其对环境因子的适应能力,为其具备超强侵染能力,进而成为入侵植物提供了基础条件。
关键词: 入侵植物;净光合速率;光合有效辐射;相关性
中图分类号: Q945.11 文献标志码: A
文章编号:1002-1302(2015)08-0121-04
生物入侵(biological invasions)是指物种从原来的分布区扩展到新的地区,能繁殖扩散并可以维持种群的过程 [1]。生物能够成功入侵并造成危害,不仅与其自身的生物学特性有关,还受到光照、营养水平等诸多环境因子的影响。通过对薇甘菊(Mikania micrantha) [2]、紫茎泽兰(Eupatorium adenophorum) [3]等的研究发现,外来杂草对环境的适应能力很大程度上与其光合特性有关。小飞蓬(Conyza canadensis)原产于北美洲,入侵我国后在各地均有分布,是我国分布最广的入侵物种之一,也是部分地区草坪中的重要杂草之一 [4-5]。小飞蓬能够在不同地带、不同生态系统广泛分布,可见其光合作用对周围环境有着一定的适应机制。在自然光条件下,小飞蓬净光合速率明显高于伴生植物 [6];通过小飞蓬与加拿大一枝黄花(Solidago canadensis)的光合特性对比研究发现,小飞蓬净光合速率的变化幅度及光饱和点下最大净光合速率都明显高于加拿大一枝黄花 [7-8]。上述报道集中在小飞蓬与伴生植物之间光合特性的比较,有关小飞蓬光合日变化动态及与环境因子之间的相关性却未见报道。本研究拟通过测定小飞蓬光合作用的日变化进程,分析其动态及与主要光合环境因子之间的相关性,从生理生态角度探讨小飞蓬成功入侵的光合机理,以便为具体防控提供理论依据。
1 材料和方法
1.1 研究区概况
试验样地位于辽宁省沈阳市沈阳农业大学百草园(41°46′N,123°25′E)内草坪种植区,草坪草为草地早熟禾(Poa pratensis)。近几年,该草坪逐渐被小飞蓬入侵,并成为该群落的优势杂草。该区海拔50 m,气候属北温带大陆性季风气候,四季分明,雨热同季,冬季较长,年均温8.1 ℃,≥10 ℃ 有效积温3 570.6 ℃,无霜期150~170 d,最冷月平均气温-16.5 ℃,最热月平均气温25.3 ℃,年降水量 721.9 mm,集中于7—8月,年蒸发量1 600 mm。
1.2 试验方法
试验于2011年6月15日至25日进行,选择全天晴朗无云的天气,用Li-6400便携式光合测定仪(Li-Lor Inc,USA),在2 cm×3 cm透明叶室内测定小飞蓬的光合日动态,时间为08:00—18:00,试验步长1 h。随机设置1 m×1 m样方5个,每个样方内选择处于营养生长期且健康无病虫害小飞蓬植株1株,选取由上至下第2、3片叶,并保持叶片自然生长角度不变,每株测定2片叶,每片叶测3次,共测30次。每测完1个叶片对其挂牌编号,便于下次测定。由于小飞蓬的叶片狭长,不能充满叶室内,试验结束后摘下编号叶片,用方格纸计算叶面积,并重新计算各光合参数。为反映自然状况下小飞蓬光合特征与环境因子的关系,测定过程未进行温、湿度和CO2的控制,气体流速400 mmol/s。
1.3 测定指标
小飞蓬光合特性因子:净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)、叶面饱和蒸汽压亏缺(VPDl)、叶片温度(Tl);环境因子:光合有效辐射(PAR)、空气温度(Ta)、空气相对湿度(RH)、空气CO2浓度(Ca)。根据光合和环境参数可计算水分利用率WUE=Pn/Tr和气孔限制值Ls=1-Ci/Ca。
1.4 数据分析
采用SPSS 17.0和Excel 2007软件对数据进行分析。首先分析光合特性因子的日动态分布规律,然后利用线性回归阐明环境因子对净光合速率的影响,并进行显著性检验。
2 结果与分析
2.1 环境因子日变化
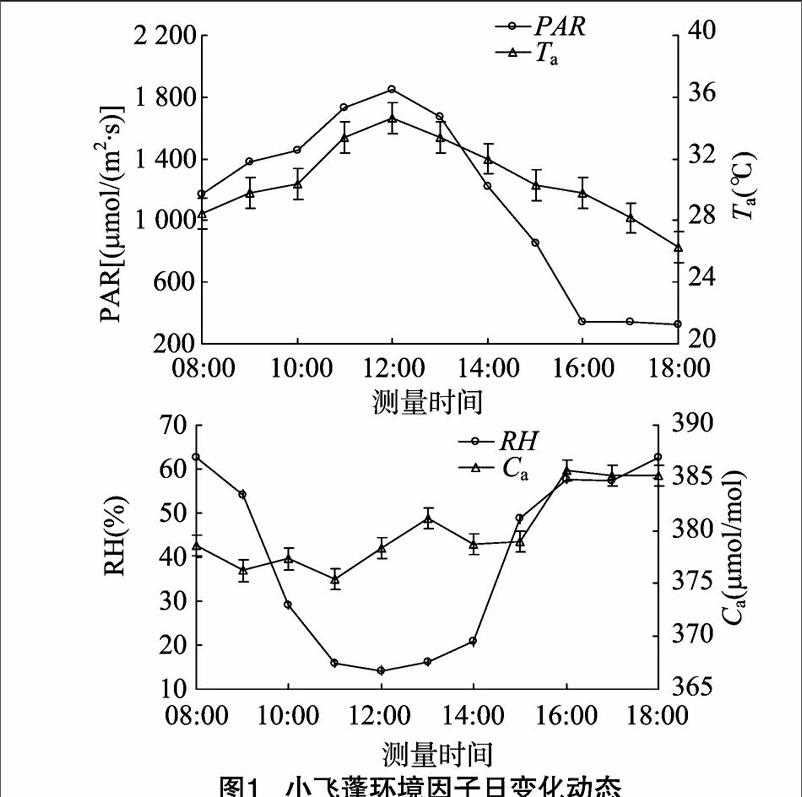
植物的光合作用受外界各种环境因子的影响,而这些因子也时刻处于变化之中。由图1可知,PAR的变化较大,09:00—14:00 均在1 200 μmol/(m2·s)以上,呈单峰曲线,峰值出现在12:00,此时达到1 849.67 μmol/(m2·s)。随着PAR的变化,Ta也表现出先上升后下降的单峰曲线,于12:00达峰值34.64 ℃。RH的变化呈倒钟形。Ca的变化呈现波折性,早晨的Ca浓度最高(385.25 μmol/mol),12:00降至最低(365.41 μmol/mol),随后又有小幅的上升,至14:00又升至峰值(376.98 μmol/mol),然后再次下降,且有小幅波动。
[FK(W18][TPWLX1.tif]
2.2 小飞蓬光合作用日变化
一般条件下,植物的光合速率日变化曲线大致呈双峰型或单峰型 [9-10]。如图2所示,小飞蓬Pn变化呈双峰曲线,峰值分别出现在11:00 [28.72 μmol/(m2·s)]和14:00[2436 μmol/(m2·s)]。由此可以看出,Pn第1个峰值明显高于第2个峰值。13:00光合速率降至低谷,呈现明显的光合“午休”现象。
[FK(W22][TPWLX2.tif;S+2mm]
Tr是植物水分代谢的一个重要的生理指标,小飞蓬Tr在08:00—10:00之间呈现一定的波动。10:00以后的变化与Pn基本一致,也出现双峰曲线,峰值分别出现在11:00和 15:00。13:00左右出现Tr降低现象。一般,植物Tr受诸多因素的影响,特别是小气候因素影响较大。因此,小飞蓬Tr在10:00之前出现了不规律的变化,很有可能是由于温度和湿度的波动导致的。但通过与Pn间对比可发现,小飞蓬Tr在很大程度上受Pn活动状态决定。
随着PAR和Ta全天呈单峰曲线变化(图1),Tl变化也呈单峰曲线,但变化趋势相对于Ta来说较缓,最高温出现在14:00,相对于Ta滞后2 h。08:00—11:00之间,Ta上升较缓,VPDl变化较小;Gs则呈现一定的波动。随着Ta升至最高点(12:00),VPDl迅速上升,Gs则有小幅降低。随着Ta的不断下降,VPDl和Gs也出现不同趋势的下降。Ci在全天的变化趋势与Gs基本一致。由此可得出,环境因子的变化,引起了小飞蓬内在生理因子产生相对的变化。
Ls反映植物叶片对大气CO2 相对利用效率的大小 [11]。经计算分析,小飞蓬叶片Ls在09:0018:00之间的变化趋势呈双峰曲线,峰值分别出现在10:00和13:00(图2)。Farquhar等认为,影响Pn下降有气孔限制和非气孔限制2方面因素。气孔限制因素是由于Gs的下降,阻止了CO2的供应;非气孔限制因素是由于叶肉细胞光合能力的下降,导致叶肉细胞质对CO2的利用降低,从而使胞间CO2的含量升高 [12]。一般用Ls和Ci的变化方向作为判别依据和标准,其中Ci是关键指标,当Pn和Gs下降时,Ls增大和Ci降低,则是气孔限制;若Ls减小和Ci增大或不变,则为非气孔限制。由图2可知,11:00—13:00,Pn的下降由气孔限制所致;而14:00—16:00 Pn的下降则为非气孔限制所致。由此可说明,小飞蓬光合速率的变化由不同机制调节。
小飞蓬WUE日变化趋势与Pn日变化趋势相一致,呈不明显的双峰曲线,且第1个峰值(1.19 μmol/mmol)明显高于第2个峰值(0.93 μmol/mmol)。08:00—11:00之间,随着PAR的增加,Ta上升,Pn增加速率超过Tr,WUE上升;此后随着Ta的上升,VPDl升高,叶面蒸发力增加,光合作用呈现“午休”现象,Tr增加速率超过Pn,WUE下降。14:00随着Pn的增大,WUE呈小幅上升。植物的WUE是由净光合速率和蒸腾速率决定的。小飞蓬的WUE变化说明其固定CO2能力较强。
2.3 小飞蓬光合速率与环境因子相关性分析
Pn的变化是由各种生理因子和环境因子综合作用的结果。小飞蓬在各测定时刻的平均Pn与环境因子的相关分析表明(表1),小飞蓬Pn与PAR、Ta呈极显著正相关,与RH呈极显著负相关,而与Ca之间的相关性不明显。由上述结果可知,小飞蓬叶片Pn主要受PAR、Ta和RH的影响。
[FK(W9][HT6H][STHZ][JZ]表1 小飞蓬净光合速率和环境因子相关分析
[HJ*5][BG(!][BHDFG3,WK5,WK24W] 指标 [ZB(][BHDWG1*2,WK24W]相关系数
[BHDWG1*2,WK6。4W] Pn PAR Ta RH[ZB)W]
[BHDG1*2,WK5,WK6。4DWW]PAR 0.95 * *
Ta 0.71 * * 0.85 * *
RH -0.69 * * -0.82 * * -0.92 * *
Ca 0.05 -0.11 -0.37* 0.45 * * [BG)F]
注:表中“* *”表示在0.01水平显著相关;“*”表示0.05水平显著相关。
对上述因子进一步进行偏相关性分析(表2) 表明,在控制其他因素的条件下,PAR与Pn的偏相关系数为0.56,在相关分析中的相关系数为0.95,这说明还有其他因素通过PAR对Pn表现了正向作用力。同样,在偏相关分析中,Pn与RH呈正相关(r=0.44),而在相关分析中Pn与RH呈负相关(r=-0.69),说明RH在其他因素干扰条件下对Pn产生了负向作用。空气中由于CO2浓度相对较为稳定,使得在自然条件下,其对净光合速率的影响不显著。但在偏相关分析中,Ca与Pn呈极显著负相关,这也说明了其他因素通过Ca对Pn产生了正向作用。通过对小飞蓬的净光合速率与环境因子的相关性和偏相关性分析,表明各个环境因子对净光合速率的变化均产生影响。
[FK(W8][HT6H][STHZ][JZ]表2 小飞蓬净光合速率和环境因子偏相关分析
[HJ*5][BG(!][BHDFG3,WK5,WK24W] 指标 [ZB(][BHDWG1*2,WK24W]偏相关系数
[BHDWG1*2,WK6。4W] Pn PAR Ta RH[ZB)W]
[BHDG1*2,WK5,WK6。4DWW]PAR 0.56 * *
Ta 0.24 0.41*
RH 0.44* -0.27 -0.58 * *
Ca -0.70 * * -0.12 0.28 -0.13 [BG)F]
注同表1。
将实际测得的小飞蓬Pn与各环境因子做回归分析,得出Pn与PAR、Ta、RH、Ca的回归方程式为Pn=-27.31+019PAR-0.624Ta+0.121Ca+0.027RH。经F检验,此回归模型P<0.001;经t检验,变量与自变量的相关性达到了显著水平,各环境因子是影响小飞蓬Pn的主要因子,R2=0926,也说明小飞蓬Pn 92.6%的日变化是由生态因子的日变化引起的。
3 讨论
植物光合作用是分析环境因子影响植物生长和代谢的重要手段,其日变化是1 d内各种生理生态因子综合效应的最终反映。外来入侵植物在入侵地往往表现出一定的生长优势,生长量累积的速度高于本地伴生植物,即光合利用率较高,光合积累能力较强 [13]。本研究对入侵植物小飞蓬光合作用的Pn、Gs、Tr及PAR等数据进行同期观测,发现小飞蓬的净光合速率日变化呈双峰曲线,具有明显“午休”现象;蒸腾速率日变化基本呈双峰曲线;叶温日变化呈单峰曲线;胞间CO2浓度与气孔导度日变化呈一定的波动变化;水分利用率与净光合速率的变化趋势基本一致。
小飞蓬叶片净光合速率日变化呈明显双峰曲线,在13:00左右有1个光合“午休”低谷。白兰花(Michelia alba)和羊草叶片的净光合速率同样存在光合“午休”现象 [14-15],此现象在自然界普遍存在。导致这种现象的原因与光合有效辐射、胞间CO2浓度、气孔阻力以及空气相对湿度有关。小飞蓬出现光合“午休”现象时,环境中的光合有效辐射变化幅度不大;因此,可以说光强不是引起小飞蓬光合作用“午休”的直接原因。但是,由于环境因子之间的相关性,光强的变化会引起温度和湿度产生相关的反应。在一定温度范围内,温度与净光合速率呈正相关 [16]。当空气温度高于光合作用的最适温度时,净光合速率随温度上升而下降。这是由于高温引起催化暗反应的有关酶钝化、变性甚至遭到破坏,同时高温还会导致叶绿体结构发生变化和受损,加剧植物的呼吸作用,而且使CO2溶解度下降,叶片温度和蒸腾速率增高,叶片失水严重,造成气孔关闭,使CO2供应不足,这些因素的共同作用,必然导致光合速率急剧下降 [17-18]。本研究表明:小飞蓬出现光合“午休”时,Tl为29.26 ℃,未达到全天最高点,说明叶片温度并未影响小飞蓬的光合能力。那么引起光合速率下降的原因可能是气孔阻力。在13:00时,Ci浓度略降,Gs为全天最低[0.39 mol/(m2·s)]。许多研究发现植物的Gs与Pn呈显著相关性 [19-21]。根据Farquhar理论,Gs下降是引起小飞蓬光合“午休”的主要原因 [12]。
植物光合作用是陆地生态系统能量吸收固定、物质分配转化、碳水循环的基础环节,也是陆地表层物质循环和能量交换的重要生理生态过程。自然条件下,植物光合作用受多个环境因子的交互影响 [22-25]。光合有效辐射是光合作用中光能的唯一来源,CO2则是光合作用的基本原料 [25-26]。欧志英等认为短期高浓度CO2处理使植物净光合速率显著增强,但长期处理则使净光合速率显著下降 [27]。但本研究发现净光合速率的高低与空气中的CO2浓度相关性不明显。原因可能是由于CO2浓度高低并不是小飞蓬净光合速率的主要限制因子,此时净光合速率主要受光合有效辐射等其他环境因子影响。本研究发现小飞蓬叶片净光合速率对光合有效辐射呈显著正相关(P<0.01)。丁继辉等发现水稻的净光合速率随着温度和光合有效辐射强度的增高而急速上升 [28],邓穗生等发现番荔枝的净光合速率和光合有效辐射变化趋势几乎一致 [29]。植物为了提高本身的竞争能力,在长期的进化过程中,其净光合速率能适应周围不断变化的环境因子。小飞蓬作为一种典型入侵植物,能够在不同地带、不同生态系统广泛分布,可见其净光合速率对周围环境因子存在一定的适应机制。在我国74个地区的入侵植物中小飞蓬的出现频率最高,且生态位相对较宽 [30],证明了小飞蓬对环境的强大适应能力和对资源的强大竞争能力。
通过对小飞蓬净光合速率与环境因子之间的相关性分析,可得出环境因子的综合作用使得小飞蓬日光合特性产生一系列相应的生理生态响应,且这种响应已经通过遗传机制成为一种稳定的性状表现出来。本试验得出的结论有助于对小飞蓬入侵机制的研究和防治策略的制定。
参考文献:
[1] Elton C S. The ecology of invasions by animals and plants[M]. London:Chapman and Hall,1958:3-5.
[2] 温达志,叶万辉,冯惠玲,等. 外来入侵杂草薇甘菊[JP3]及其伴生种基本光合特性的比较[J]. 热带亚热带植物学报,2000,8(2):139-146.
[3]刘伦辉,谢寿昌,张建华. 紫茎泽兰在我国的分布、危害与防除途径的探讨[J]. 生态学报,1985,5(1):1-6.
[4]高兴祥,李 美,高宗军,等. 外来入侵植物小飞蓬化感物质的释放途径[J]. 生态学报,2010,30(8):1966-1971.
[5]高兴祥,李 美,高宗军,等. 外来物种小飞蓬的化感作用初步研究[J]. 草业学报,2009,18(5):46-51.
[6]王晓红,纪明山. 入侵植物小飞蓬及其伴生植物的光合特性[J]. 应用生态学报,2013,24(1):71-77.
[7]胡天印,方 芳,郭水良,等. 外来入侵种加拿大一枝黄花及其伴生植物光合特性研究[J]. 浙江大学学报:农业与生命科学版,2007,33(4):379-386.
[8]顾智华,王济军,周 捷,等. 不同光强处理下加拿大一枝黄花的短期光合特性研究[J]. 杂草科学,2009(4):23-27.
[9]张治安,杨 福. 光合速率日变化及其与环境因子的相关关系[J]. 中国农业科学,2006,39(3):502-509.
[10] Yang J D,Liu Z M. Study on field-grown maize introduced into Tibetan plateau:some characteristics of diurnal variation of photosynthesis[J]. Acta Agronomica Sinica,2002,28:475-479.
[11]Xu B C,Shan L,Huang Z B,et al. Comparative of photosynthetic characteristics of Old World Bluest ems(Bothriochloa ischaemum) and Switch grass(Panicum virgatum)in Loes shilly gully region of China[J]. Grassland of China,2003,25(1):1-4.
[12]Farquhar G D,Sharkey T D. Stomatal conductance and photosynthesis[J]. Rev Plant hysiol,1982,33:317-345.
[13]何锦峰. 外来植物入侵机制研究进展与展望[J]. 应用与环境生物学报,2008,14(6):863-870.
[14]王玉辉,周广胜. 松嫩草地羊草叶片光合作用生理生态特征分析[J]. 应用生态学报,2001,12(1):75-79.
[15]尤 扬,杨立峰,周 建,等. 白兰花秋季光合特性研究[J]. 西北林学院学报,2009,24(6):24-27.
[16]李春鸣,蒲小朋,马姝雯,等. 高寒地区地膜覆盖栽培饲用甜菜光合日变化初探[J]. 西北民族大学学报:自然科学版,2005,26(1):46-48.
[17]许大全. 光合作用气孔限制分析中的一些问题[J]. 植物生理学通讯,1997,33(4):241-244.
[18]许大全. 光合作用“午睡”现象的生态、生理与生化[J]. 植物生理学通讯,1990,26(6):5-10.
[19]Bethenod O,Tardieu F,Katerji N. Relationship between net photosynthetic rate and stomatal conductance in leaves of field-grown maize subjected to soil or soil drying[J]. Photosynthetica,1996,32(3):367-379.
[20]Cai Z Q,Slot M,Fan Z X. Leaf development and photosynthetic properties of three tropical tree species with delayed greening[J]. Photosynthetica,2005,43(1):91-98.
[21] Ye E P,Yu Q. A coupled model of stomatal conductance and [JP3]photosynthesis for winter wheat[J]. Photosynthetica,2008,46(4):637-640.
[22]闻 婧,杨其长,魏灵玲,等. 不同波峰的LED红蓝光质组合对黄瓜苗生长和光合特性的影响[J]. 江苏农业学报,2013,29(3):619-625.
[23]叶子飘. 光合作用对光和CO2响应模型的研究进展[J]. 植物生态学报,2010,34(6):727-740.
[24] 彭丽丽,姜卫兵,韩 健,等. 连续剪梢对金叶女贞不同叶位呈色和光合特性的影响[J]. 江苏农业学报,2013,29(2):399-404.
[25]余叔文,汤章城. 植物生理与分子生物学[M]. 北京:科学出版社,1992:567-569.
[26]Yu Q,Zhang Y Q,Liu Y F,et al. Simulation of the stomatal conductance of winter wheat in response to light,temperature and CO2 changes[J]. Annals of Botany,2004,93(4):435-441.
[27]欧志英,彭长连. 高浓度二氧化碳对植物影响的研究进展[J]. 热带亚热带植物学报,2003,11(2):190-196.
[28]丁继辉,俞双恩,于智恒,等. 农田水位调控下水稻净光合速率日变化规律试验研究[J]. 河海大学学报:自然科学版,2011,39(1):104-108.
[29]邓穗生,陈业渊,林 玲. AP番荔枝的光合特性[J]. 热带作物学报,2005,26(4):34-37.
[30]张 帅. 外来植物小飞蓬入侵生物学研究[D]. 上海:上海师范大学,2010.
猜你喜欢
商情(2016年42期)2016-12-23
商情(2016年42期)2016-12-23
东方教育(2016年4期)2016-12-14
中国实用医药(2016年24期)2016-10-17
中国实用医药(2016年24期)2016-10-17
中国实用医药(2016年24期)2016-10-17
科学与财富(2016年28期)2016-10-14
