
茶尺蠖潜在飞行能力的研究
2015-08-24 08:53徐秀秀蔡晓明边磊罗宗秀陈宗懋中国农业科学院茶叶研究所农业部茶树生物学与资源利用重点实验室浙江杭州310008
茶叶学报 2015年4期
徐秀秀,蔡晓明,边磊,罗宗秀,陈宗懋(中国农业科学院茶叶研究所/农业部茶树生物学与资源利用重点实验室,浙江 杭州 310008)
茶尺蠖潜在飞行能力的研究
徐秀秀,蔡晓明,边磊,罗宗秀,陈宗懋*
(中国农业科学院茶叶研究所/农业部茶树生物学与资源利用重点实验室,浙江 杭州 310008)
为探究茶尺蠖成虫的飞行扩散能力,本研究利用昆虫飞行信息系统,室内测定了茶尺蠖不同日龄、不同性别及交配前后成虫的飞行能力。结果表明,雌雄成虫均以2日龄的飞行能力最强,平均飞行距离分别达9.14 km和8.16 km,一次最远的飞行距离分别为2.56 km和1.93 km。雌成虫的飞行能力在3日龄时显著下降,雌雄成虫的飞行能力仅在3日龄时表现显著差异。交配对2日龄雌雄成虫的飞行能力无显著影响。
茶尺蠖成虫;日龄;性别;飞行能力;交配
茶尺蠖(EctropisobliquaProut)属鳞翅目(Lepidoptera)尺蛾科(Geometridae),是茶树最主要的害虫之一。该虫在我国所有的茶区均有发生,尤其以长江中下游危害最为严重[1]。主要以幼虫取食茶树叶片为害,发生严重时使茶园大面积成为秃枝,严重地影响茶叶的产量和品质[2]。长期以来,化学防治一直是控制茶尺蠖猖獗危害的主要手段。但是,化学农药的大量使用,不仅使害虫产生了严重的抗药性,还对其天敌及其他有益生物造成了不利影响,削弱了自然因素的生态调控作用[3]。同时还造成茶叶中农药的大量残留,直接影响了人体的健康和茶叶的出口贸易[4]。近年来,人们开始考虑一些其他的防治方法来替代化学农药,从而减少其对人类身体健康和环境的不利影响,如茶尺蠖性信息素及其他引诱剂的研究和应用[5-6],诱虫灯的开发与利用[7-8],寄生和捕食性天敌的利用,植物源农药[9-10]、病毒制剂[11-12]的研究等都取得了一定的进展。
无论是使用性信息素或其他引诱剂引诱成虫,还是应用诱虫灯对其进行诱杀,都是将“诱集陷阱”固定,使茶尺蠖根据其对信息素的喜好和趋光性等进入诱捕器。然而任何防治方法都需考虑到成本,而这些诱捕器的合理布置是节约成本的最根本手段。要做到最大程度地降低成本并达到最好的诱杀效果,除了筛选出光源的合适波长和信息素的合适配方以外,还需要了解昆虫的活动能力,从而确定诱捕器的悬挂高度和密度[13]。但是关于茶尺蠖扩散行为和飞行能力的研究国内外均未见报道。因此本研究以飞行磨吊飞昆虫作为主要手段,在实验室条件下测定了茶尺蠖不同日龄和不同性别成虫的飞行能力,以及交配对其飞行能力的影响,以期为研究茶尺蠖成虫的诱杀技术提供参考。
1 材料方法
1.1 虫源及处理
2014年6~8月从杭州市西湖区茶园中采集茶尺蠖幼虫,携至室内(25±2℃,RH 40%~50%,光周期14L:10D)用新鲜茶枝饲养。幼虫化蛹后,在解剖镜下将雌雄蛹分开放置。待成虫羽化后,将相同日龄的雌雄蛾配对放入养虫笼,分别于不同日龄进行飞行能力测试;另一部分同日羽化的茶尺蠖雌雄蛾进行单对配对,于1日龄晚间(22:00~0:00)观察其交配情况,将交配过的处理进行标记,并于2日龄时测定其飞行能力;另一种处理是将羽化后的雌雄蛾分开放置,待2日龄时分别对雌雄蛾进行飞行能力测试。成虫用棉球蘸取10%的蜂蜜水饲喂,为防止霉变或变干,新鲜棉球每日更换一次。
1.2 仪器
飞行能力测试采用佳多科工贸有限公司生产的由电脑控制的24通道昆虫飞行磨采集系统。该系统可自动记录昆虫飞行过程中的飞行速度、飞行时间以及飞行距离的变化,并可对测试数据进行分析处理。
1.3 测定方法
所有的测试均在相同的环境条件下进行。吊飞过程中室内完全保持黑暗,温度控制在25±1°C,湿度控制在30%~40%。吊飞前,先将供试个体放在指形玻璃管里冷冻麻醉10 min,然后用毛笔将前胸背板处的鳞毛除去,用百多胶将前胸背板粘接于直径为0.8 mm塑料管上,再将该塑料管插在飞行磨吊臂上(注:保持昆虫飞行的切线方向与吊臂垂直),吊臂半径为10 cm。吊飞试验于每日19:00左右开始,吊飞时间为12 h,次日7:00左右结束。除4日龄外,每个处理重复次数不小于20次。
1.4 统计分析
不同日龄成虫飞行能力(平均飞行速度,累计飞行时间和累计飞行距离,一次最大飞行距离,一次最长飞行时间和最大飞行速度)均采用飞行磨自带的软件进行初步的分析。在进行多重比较之前,将数据进行对数转换并检验方差齐性,然后采用LSD法对不同日龄成虫的飞行能力进行差异显著性比较,而同一日龄不同性别的比较采用t检验来进行分析。所有数据均采用SPSS18.0软件进行分析。
2 结果分析
2.1 不同日龄茶尺蠖成虫飞行距离的频率分布
茶尺蠖成虫12 h内在飞行磨上的飞行距离接近于对数分布(图 1)。随着日龄的增长,总飞行距离在0~4 km之间的虫数所占的比例增大,3日龄成虫的飞行距离在0~4 km内已达80%,而至4日龄时,所有成虫的飞行距离均在0.5 km之内。1、2日龄的飞行能力较强,飞行能力大于4 km的数量分别达50%和60%以上。
2.2 不同日龄茶尺蠖成虫的飞行能力
在12 h无光照条件下,分别测定了不同日龄雌雄成虫的飞行能力(表 1)。结果表明,雌雄成虫均以2日龄的飞行能力最强,飞行距离分别达到9.14 km 和8.16 km。对雄虫来讲,无论是在总的飞行距离、平均飞行速度上,1~3日龄之间的雄成虫均无显著差异,而4日龄则显著减小;而2、3日龄雄虫的平均飞行时间也存在显著差异,4日龄雄虫则显著减小至0.18 h。对雌虫来说,1、2日龄飞行距离、飞行时间和平均飞速显著高于3、4日龄。
利用LSD法分析了不同日龄茶尺蠖成虫的最远持续飞行距离,最大飞行速度和最长持续飞行时间(表 2)。结果表明,雌、雄成虫的各项飞行特征均以 2日龄最强,一次最远的飞行距离分别达到2.56±0.73 km和1.93±0.50 km。对雄蛾来说,在1~3日龄之间各项飞行特征差异不大,而4日龄显著减弱;而雌成虫则在3日龄时各项飞行指标显著减弱,一次最远的飞行距离仅为0.19 km,4日龄的飞行能力最弱。
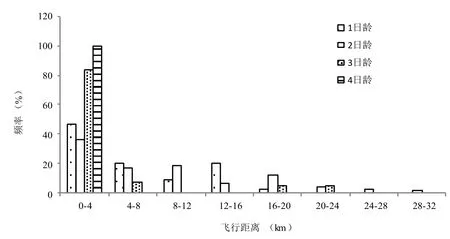
图 1 不同日龄茶尺蠖成虫的飞行距离频率分布Fig. 1 Flying distances of adult E. oblique of different ages
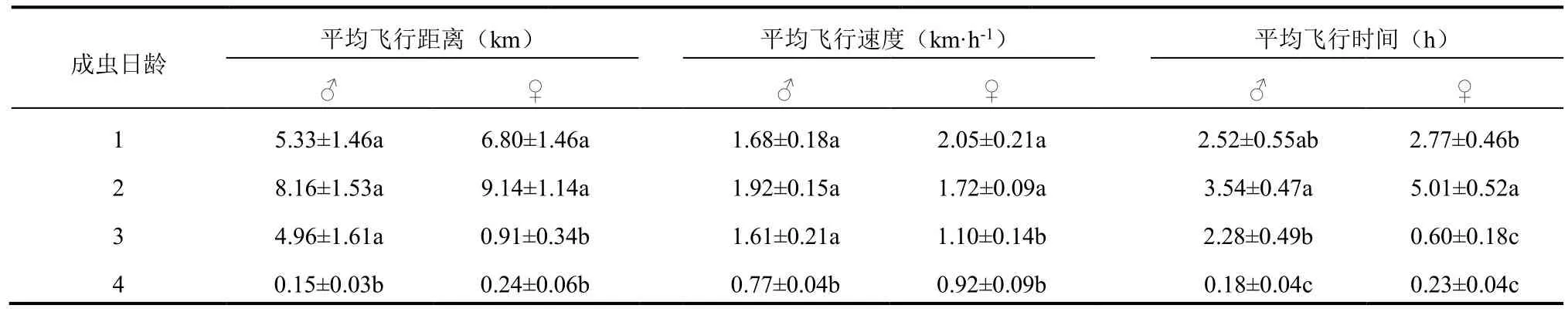
表1 不同日龄茶尺蠖成虫的飞行能力Table 1 Flight capacities of adult E. oblique of different ages

表2 不同日龄茶尺蠖成虫的一次最远飞行距离,最大飞行速度和最长飞行时间Table 2 Longest continuous flight, flying speed, and flying duration of adult E. oblique of different ages
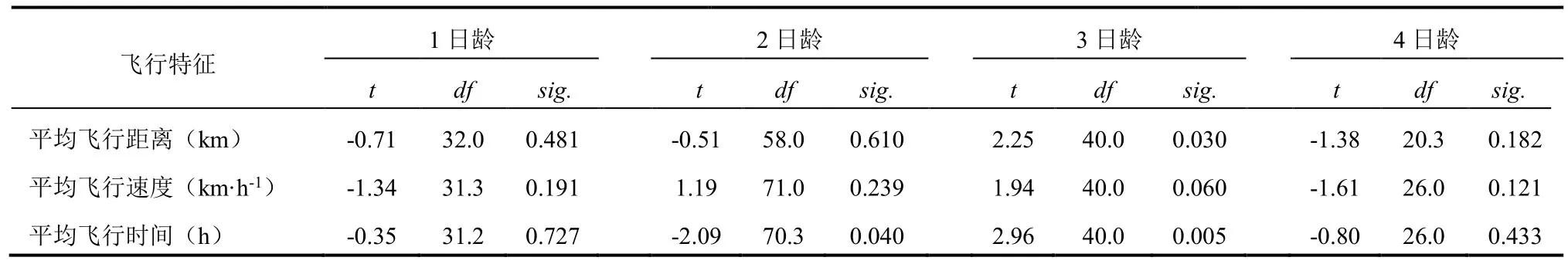
表3 茶尺蠖雌雄成虫飞行能力的比较Table 3 Flight capabilities of male and female adult E. obliqua
2.3 不同性别茶尺蠖成虫的飞行能力
不同性别茶尺蠖的飞行能力也存在某些差异(表3)。t检验结果表明,1、4日龄雌雄成虫的飞行能力无显著差异;仅3日龄的雄蛾的平均飞行距离和平均飞行时间显著大于雌蛾,二者的平均飞行速度则差异不明显。2日龄雌、雄成虫的平均飞行时间也存在显著差异。
2.4 交配和未交配茶尺蠖雌成虫的飞行能力
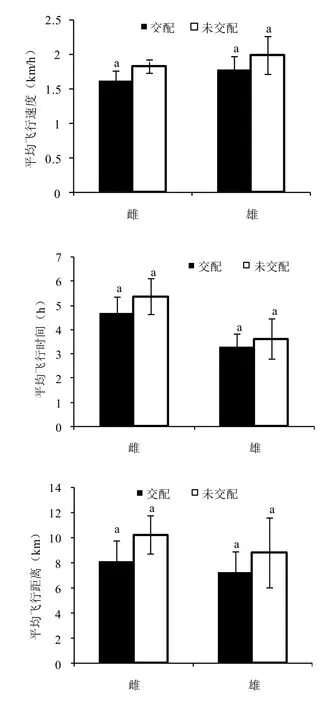
图 2 交配和未交配茶尺蠖成虫飞行能力比较Fig. 2 The flight capabilities of mated and unmated E. obliqua adults
交配与否对茶尺蠖雌蛾和雄蛾的飞行能力均无影响(图2)。从图2可以看出,交配和未交配的2日龄雌雄蛾平均飞行距离、平均飞行速度和平均飞行时间的统计分析差异并不显著。未交配的雌雄蛾平均飞行距离、平均飞行速度和平均飞行时间均较交配后的大,但均未表现出显著差异。
3 讨论
茶尺蠖成虫具有较强的飞行和迁移扩散能力。雌、雄成虫均以2日龄的飞行能力最强,分别达9.14 km和8.16 km,之后逐渐减弱。这与舞毒蛾成虫期的行为表现较为一致,它们在羽化初期已经具备较强的飞行能力[14]。雌雄成虫基本呈现随着日龄增加飞行能力逐渐减弱的现象,相比雄虫来说,雌成虫的飞行能力在3日龄开始已显著下降,并与雄成虫的飞行能力表现出显著差异。这可能是由于羽化后雌虫的卵巢逐渐发育成熟,3日龄已处于产卵盛期,此时其把能量储备供应于产卵而不是飞行。而此时的雄成虫仍处于活动高峰期,继续搜寻雌蛾进行交配,完成其繁育后代的使命,从而导致飞行能力产生差异。成虫飞行能力随日龄变化而变化的现象在其他的鳞翅目害虫中也较为常见,如苹果蠹蛾、稻纵卷叶螟、棉铃虫、甜菜夜蛾、小菜蛾等[15-19],这些都是由其自身的生理特性所决定的。茶尺蠖雌雄成虫的平均寿命分别为5.33 d和4.38 d[20],为了尽快繁殖后代,初羽化的成虫已具备较强的飞行能力,至4日龄已到达生命的后期,此时已完成交配和产卵,成虫使命已基本完成,因此飞行能力基本丧失,这与该虫的生物学特征也是相符的。
茶尺蠖成虫一般于1日龄的晚间完成交配,2、3日龄晚间进行产卵[21]。幼虫的活动能力有限,因此交配后雌虫的飞行能力决定着其后代幼虫的危害范围,本实验选用2日龄的成虫用于研究交配对其飞行能力的影响。结果表明,交配对2日龄雌雄成虫的飞行能力并无显著影响,也就是说,交配后的雌成虫并未将能量全部用于产卵,寻找合适的产卵场所和适宜后代生存的寄主也是其生命中不可缺少的一部分。雄成虫一生要经历多次交配,交配对其飞行能力的影响也不显著。本试验结果可以初步的证明,2日龄时更适于茶尺蠖进行远距离的飞行,所以用杀虫灯进行诱杀或性信息素诱集时,对2日龄成虫的诱杀效果可能较佳。
温湿度、光照、幼虫期的食物供应等均可影响茶尺蠖成虫的飞行和扩散能力[22-24],而本研究仅在实验室条件下初步测定了不同日龄成虫的飞行能力。并且飞行磨的强迫飞行与自然条件下昆虫的主动飞行也有一定的区别[25],因此本实验结果只能作为茶尺蠖成虫飞行潜力的一个辅助评估。茶尺蠖成虫在自然条件下的飞行和扩散能力还需进一步研究。
[1] 张汉鹄,谭济才.中国茶树害虫及其无公害治理[M].合肥:安徽科技出版社,2004.
[2] 熊兴平.茶尺蠖防治技术研究进展及展望[J].中国茶叶,2003,(3):15-17.
[3] 徐德良,田晓兰,吴培中.江苏茶尺蠖抗药性评估及治理对策[J].茶业通报,2006,28(1):15-16.
[4] 谭正初,萧力争.我国茶叶质量安全现状与控制[J].茶叶通讯,2008,35(1):18-23.
[5] 赵信.茶尺蠖性信息素的合成、应用及延迟交配对茶尺蠖寿命和繁殖力的影响[D].合肥:安徽农业大学,2012.
[6] 胡义元,黄斌,段巧枝,等.茶尺蠖信息素诱捕茶园茶尺蠖试验报告[J].湖北植保,2014,(6):19-20.
[7] 郭华伟,周孝贵,肖强,等.10种光源对茶园中不同昆虫的诱集效果[J].中国茶叶,2012,(6):18-20.
[8] 万利.昆虫趋光性在茶园害虫防治中的作用[D].武汉:华中农业大学,2014.
[9] 董道青,陈建明,俞晓平,等.茶皂素对雷公藤乳油防治茶尺蠖幼虫的增效作用[J].茶叶科学,2008,28(3):228-233.
[10] 张兰美,陈柏寿.茶核-苏云金防治茶尺蠖的田间药效试验[J].中国茶叶,2011,(11):15.
[11] Ma XC, Xu HJ, Zhang CX, et al. Morphological,phylogenetic and biological characteristics of Ectropis obliqua single-nucleocapsid nucleopolyhedrovirus [J]. Journal of Microbiology, 2006,44(1):77-82.
[12] 王定锋,王庆森,吴光远.茶尺蠖病毒研究进展[J].茶叶科学技术,2013,(4):1-5.
[13] 边磊,孙晓玲,陈宗懋.假眼小绿叶蝉的日飞行活动性及成虫飞行能力的研究[J].茶叶科学,2014,34(3):248-252.
[14] 杨帆,骆有庆,黄东辉,等.亚洲型舞毒蛾中国地理种群飞行能力初探[J].中国农学通报,2012,28(28):53-57.
[15] Schumacher P, Weyeneth A, Weber DC, et al. Long flights in Cydia pomonella L. (Lepidoptera:Tortricidae) measured by a flight mill: influence of sex, matedstatus and age [J]. Physiological Entomology, 1997,22(2):149-160.
[16] 徐秀秀.稻纵卷叶螟迁飞与生殖的关系及其生理机制研究[D].北京:中国农业科学院研究生院,2012.
[17] Armes NJ, Cooter RJ. Effects of age and mated status on flight potential ofhelicoverpa armigera (Lepidoptera:Noctuidae)[J]. Physiological Entomology,1991,16(2): 131-144.
[18] Han LZ, Gu HN, Zhai BP,et al. Reproduction-flight relationship in the beet armyworm, Spodoptera exigua (Lepidoptera: Noctuidae) [J]. Environmental Entomology,2008,37(2):374-381.
[19] 魏书军,范潇,顾耘,等.不同日龄及交配前后小菜蛾飞行能力[J].应用昆虫学报,2013,50(2):474-482.
[20] 杨云秋,宛晓春,郑高云,等.茶尺蠖性行为习性初报[J].中国农学通报,2008,24(2):339-342.
[21] 高旭晖,宛晓春,杨云秋,等.茶尺蠖生物学习性研究[J].植物保护,2007,33(3):110-113.
[22] 江幸福,蔡彬,罗礼智,等.温、湿度综合效应对粘虫蛾飞行能力的影响[J].生态学报,2003,23(4):738-743.
[11] 苏春芳,刘爱萍,高书晶,等.温度和湿度对草地螟阿格姬蜂飞行能力的影响[J].中国生物防治学报,2014,30(5):612-617.
[23] 曹雅忠,罗礼智,郭军.粘虫生殖和飞翔与幼虫期营养的关系[J].昆虫学报,1996,39(1):105-108.
[24] 翟保平,张孝羲.迁飞过程中昆虫的行为:对风温场的适应与选择[J].生态学报,1993,13(4):356-363
Flight Capability of the Tea Geometrid, Ectropis oblique Prout (Lepidoptera: Geometridae)
XU Xiu-xiu,CAI Xiao-ming,BIAN Lei,LUO Zong-xiu,CHEN Zong-mao*
(Tea Research Institute, Chinese Academy of Agricultural Sciences, Key Laboratory for Tea Plant Biology and Resource Utilization, Ministry of Agriculture, Hangzhou Zhejiang 310008, China)
This study aimed to determine the flight capability and dispersal of adult tea geometrid, Ectropis oblique Prout. The flying speed, distance and duration of the insect were monitored using the computer-linked flight mills in the laboratory. The flight capability of the insects was at the highest 2 days after their emergence, and mating did not appear to significantly affect it. For the female and male adults, their average flying distances were 9.14 km and 8.16 km, respectively, and the longest continuous flights were 2.56 km and 1.93 km, respectively. The flight capability of tea geometrids tended to increase initially and declined significantly after they were 3-d-old, particularly for the females.
Ectropis oblique Prout; age; gender; flight capacity; mating
S435.711
A
2015-09-02初稿;2015-09-28修改稿
徐秀秀(1985-),女,博士研究生,主要从事昆虫化学生态学研究。
zmchen2006@163.com
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23
茶叶科学(2021年4期)2021-08-12
山西林业(2021年2期)2021-07-21
四川蚕业(2021年1期)2021-02-12
浙江农业学报(2020年8期)2020-08-30
昆虫学报(2020年6期)2020-08-06
昆虫学报(2020年1期)2020-03-03
——茶尺蠖和灰茶尺蠖的识别
中国茶叶(2019年11期)2019-12-03
爱你(2017年13期)2017-11-15
中国马铃薯(2015年3期)2015-12-19
