
利用SSR标记分析福建省选育福云半同胞系茶树品种遗传多样性
2015-08-24 08:59:41陈志辉单睿阳林郑和游小妹钟秋生陈常颂福建省农业科学院茶叶研究所福建福安355015
茶叶学报 2015年4期
陈志辉,单睿阳,林郑和,游小妹,钟秋生,陈常颂(福建省农业科学院茶叶研究所,福建 福安 355015)
利用SSR标记分析福建省选育福云半同胞系茶树品种遗传多样性
陈志辉,单睿阳,林郑和,游小妹,钟秋生,陈常颂*
(福建省农业科学院茶叶研究所,福建 福安 355015)
利用SSR分子标记技术对福建省选育的福云半同胞系茶树品种遗传多样性进行研究。通过18对SSR引物PCR扩增13个参试品种,共获得108条谱带,其中多态性谱带占67.59%。遗传相似系数分析发现,半同胞系品种间相似系数在0.75~0.91之间,而对照种相似系数在0.68~0.97之间,说明对照种遗传多样性比半同胞系更丰富。13个参试品种聚类结果表明,早春毫等3个对照种与其它品种遗传距离最远,同时这3个品种由于相互间的遗传距离较远而没有聚合成簇;剩余10个品种聚合成2组,其中5个半同胞系全部聚合于Ⅰ组,而母本福鼎大白茶与其它4个对照种聚合于Ⅱ组。从聚类图中进一步看出对照种的遗传多样性比半同胞系丰富。通过分析聚类结果与遗传相似系数发现遗传距离与品种间的地理距离、品种的亲本来源及品种性状特征具有重要的相关性。茶树遗传多样性分析为茶树分子标记辅助选择、茶树杂交育种及遗传改良提供依据,以及为茶树栽培选种及引种提供指导作用。
福云半同胞系;遗传多样性;SSR分子标记
半同胞系是一种近亲系,在林木品种遗传改良与选育中被广泛应用[1-4]。茶树是我国重要的经济林木,对茶树品种进行遗传改良具有重要的生产应用价值。福鼎大白茶和云南大叶种是我国茶树育种的重要骨干亲本。据统计,我国育成的66个国家级茶树无性系品种中,以福鼎大白茶为亲本选育的品种或衍生品种有12个(占18.18%);以云南大叶种为亲本选育的品种或衍生品种有29个(占43.94%)[5-6]。福鼎大白茶与云南大叶种杂交后代表现出显著的杂种优势,是优良的茶树育种材料。福建省农业科学院茶叶研究所郭元超研究员利用福云系列的杂种优势,选育出多个国家级、省级茶树品种。其中福云6号在福建、广西、浙江有较大面积栽培,湖南、江西、四川、贵州、安徽、江苏、湖北等省有引种,是当前全国推广面积最大的人工杂交品种,为茶产业发展做出了突出贡献。
随着生物技术的发展,分子标记技术在茶叶研究中得到广泛应用,尤其在遗传多样性及亲缘关系鉴定中使用最多[7-15]。分子标记技术经历了RFLP、RAPD、AFLP、ISSR、SSR、SNP等发展阶段,其中SSR即简单序列重复(Simple sequence repeats,SSR),因其具有重复性好、多态性高、共显性、易于检测和鉴别能力高等优点,而被广泛应用[16-20]。在分子标记技术不断发展过程中,它的应用也从早期的茶树种质资源鉴定[21-26]逐步转向分子标记辅助筛选、遗传连锁图谱构建及QTL定位[27-29]等研究领域。本研究预期利用SSR分子标记技术,分析福建省选育的福云半同胞系茶树品种的遗传多样性,为茶树分子标记辅助选择,茶树杂交育种与遗传改良提供根据,以及在茶树栽培选种与引种中提供指导。
1 材料与方法
1.1 参试品种(见表1)
1.2 试验方法
采用CTAB法提取茶树基因组DNA。SSR反应体系、扩增程序及PAGE胶电泳检测参考张成才等[18]的方法。
1.3 条带统计及聚类分析方法
根据PAGE胶电泳胶图统计条带,对于同一引物的扩增产物,迁移位置相同的条带记为1个位点,即扩增阳性(有较为清晰条带出现)赋值为“1”,扩增阴性(无条带或者弱带)赋值为“0”,所得数据输入Excel建立原始数据矩阵,待进一步分析。其中任一位点若有一个或一个以上个体不同于其他个体即为一个多态位点。以上所得数据按SHAN邻接法对供试种质资源进行UPGMA遗传相似性聚类,并绘制树状聚类图。数据统计分析在NTSYS软件系统下进行。
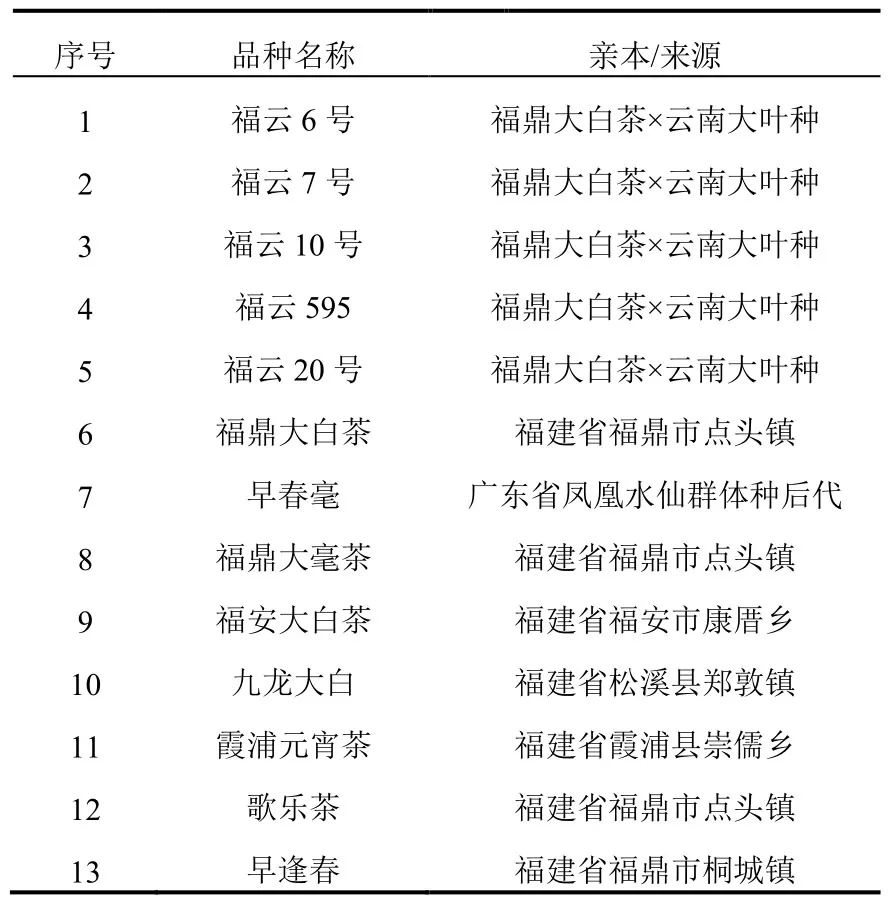
表1 参试品种及来源Table 1 Cultivars studied and their origins
2 结果与分析
2.1 PCR扩增与电泳及谱带统计分析
本试验采用SSR分子标记技术研究福建省选育的福云半同胞系茶树品种遗传多样性。实验从40对SSR引物中筛选获得18对扩增效果好,条带清晰的引物,利用该批引物对13个参试品种进行PCR扩增及聚丙烯酰胺凝胶电泳,在通过固定、银染与显色,最后进行读带与统计分析。SSR分子标记扩增效果见图1,该图所展示的是引物A134对13个参试品种扩增的谱带情况。实验共从18对SSR引物的电泳胶图中获得108条谱带,平均每个引物扩增的谱带数6条,其中73条有多态性,多态性程度为67.59%。条带统计及聚类分析详见材料与方法。
2.2 遗传多样性分析
福建省选育的福云半同胞系茶树品种共有5个,即福云6号、福云7号、福云10号、福云595和福云20号,由福鼎大白茶(母本)与云南大叶种(父本,群体种)自然杂交,通过单株育种法选育而成。本实验13个参试品种详见表1,其中序号1、2、3、4、5为半同胞系茶树品种,序号6为母本福鼎大白茶,实验为了更好的呈现品种间的遗传多样性程度,随机选取7个福建省选育的其它绿茶品种作为对照种。

图 1 SSR引物A134对13个参试品种扩增产物的聚丙烯酰胺凝胶电泳图Fig. 1 Polyacrylamide gel electrophoresis on amplified products from 13 cultivars using SSR primers, A134
对参试品种的SSR分子标记实验数据进行遗传相似性及聚类分析发现,福云半同胞系各品种间的遗传相似系数在0.75~0.91之间(见表2),其中福云6号与福云595遗传相似系数最低(为0.75),相似系数最高为福云595与福云7号(为0.91)。从半同胞系聚类图(见图2a)可看出,5个半同胞系与母本可明显分成2组,其中福云6号与福云10号聚于一组;福云7号、福云595、福云20号及母本福鼎大白茶聚成一组。
对随机选取的7个对照种的遗传多样性分析发现,7个对照种的相似系数介于0.68~0.97之间(见表2),其中相似系数最低为早春毫与福安大白茶以及早春毫与九龙大白茶,都为0.68,最高为福鼎大毫茶与九龙大白茶(为0.97)。通过比较半同胞系和对照种的相似系数可看出,半同胞系间的相似系数比较集中地分布于0.75~0.91之间,遗传距离跨度较小,而对照种相似系数在0.68~0.97之间,跨度较大。尽管如此,半同胞系各品种间仍呈现出较高的遗传多样性,从0.75~0.91的相似系数可看出,半同胞系各品种间的遗传距离既不靠地太近也不离地太远。这是由于亲本福鼎大白茶与云南大叶种的遗传距离较远造成,2个亲本不仅地理距离远,而且生物学特征差异显著,遗传背景差异大,而杂交后代又各自遗传了双亲的一半遗传物质,又具有较高的同源性,所以相似系数不会出现太高和太低的极端情况,0.75~0.91比较真实地呈现出本实验半同胞系品种间较高的遗传差异性。
对照种聚类结果分析发现(见图2b),7个对照种没有明显的聚类成簇,相互间的遗传距离比较均匀的间隔开,遗传多样性更丰富,遗传距离跨度大,遗传背景更为复杂,所以与对照种相比,半同胞系(见图2a)的遗传多样性相对较低。
以上相似系数和聚类结果分析表明,半同胞系品种间具有较高的遗传多样性,但比对照种遗传多样性低。
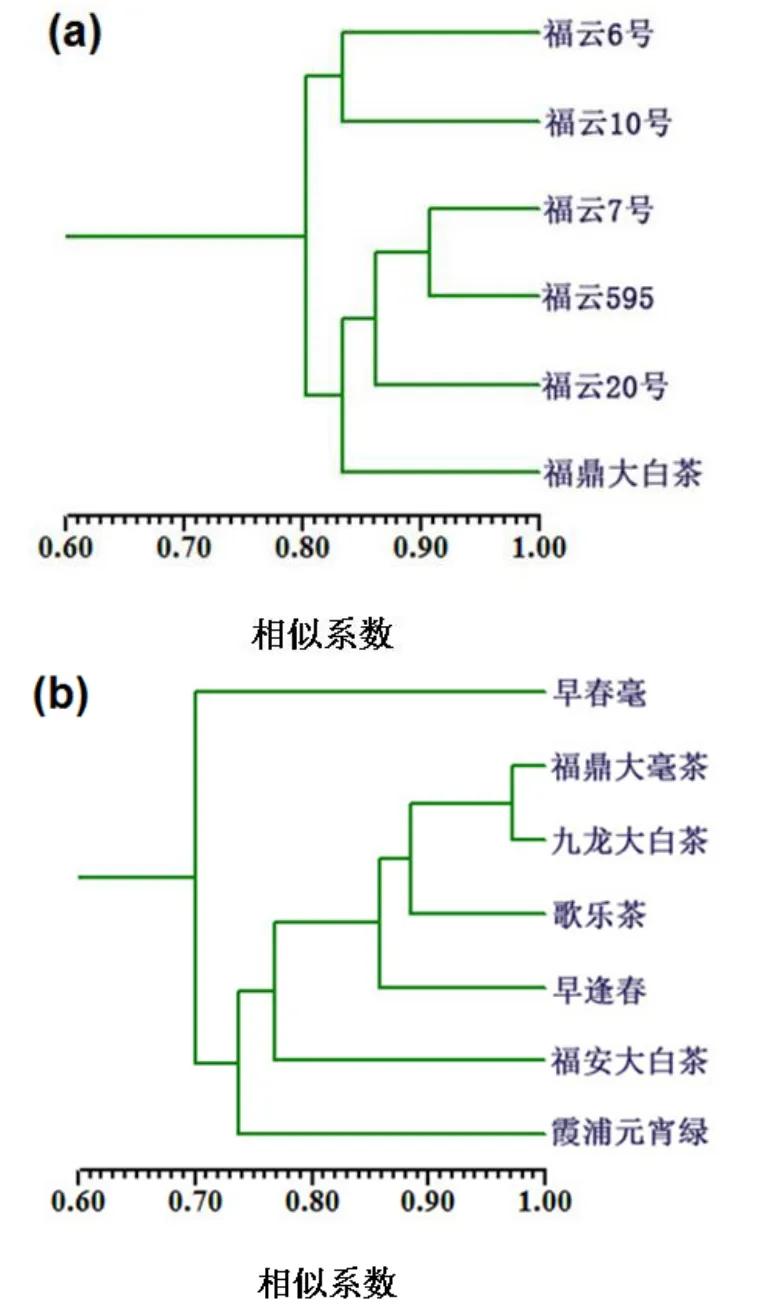
图 2 福云半同胞系(a)及对照种(b)聚类图Fig. 2 UPGMA cluster analysis on Fuyun half-sib lines(a)and cultivars for comparison as control (b)
2.3 聚类分析
13个参试品种的遗传相似系数及聚类结果(见表2,图3),从图3中可看出早春毫与所有品种距离最远,其次是霞浦元宵茶,然后是福安大白茶。剩余的10个品种聚合成2组,其中5个半同胞系正好聚于Ⅰ组,即福云6号、福云10号、福云7号、福云595、福云20号。该5个半同胞系还可进一步分成a 与b两个亚组,a亚组含有福云6号与福云10号,b亚组含有福云7号、福云595及福云20号。第Ⅱ组包含福鼎大白茶与4个对照种,分别为福鼎大白茶、歌乐茶、福鼎大毫茶、九龙大白茶和早逢春。
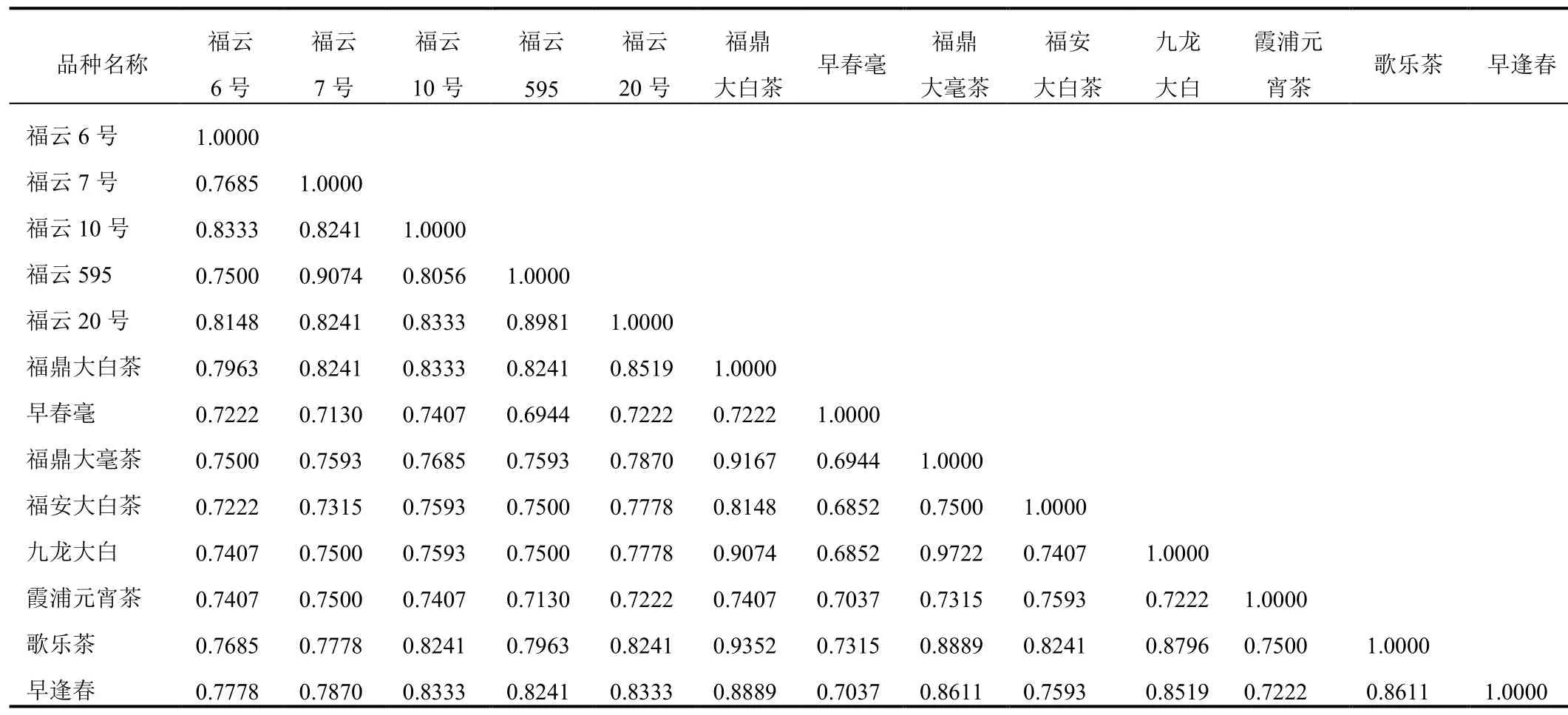
表2 13个参试品种的遗传相似系数Table 2 Genetic similarity coefficient of 13 tested cultivars
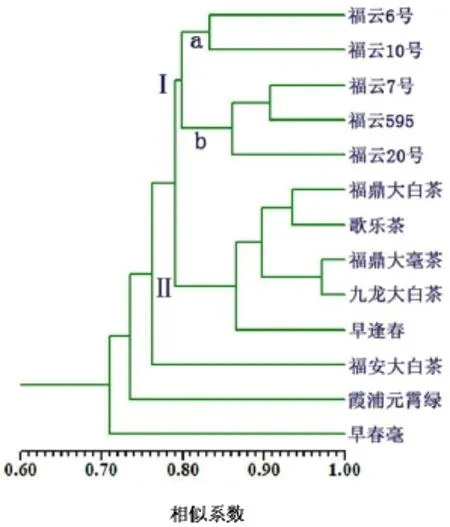
图 3 13个参试品种的遗传相似性聚类图Fig. 3 UPGMA cluster analysis on 13 cultivars
通过分析 13个参试品种的聚类图以及品种间的遗传距离,可看出其中有很强的规律性,特别与品种的发源地、品种的亲本来源及品种性状特征具有重要的相关性。
首先遗传距离最远的品种为早春毫,其次是霞浦元宵茶,然后是福安大白茶。其中早春毫为迎春自然杂交后代,而迎春是广东凤凰水仙群体种的后代,所以与其它参试品种的发源地都离得最远(其它品种都来自福建省内),聚类结果充分体现出遗传距离与地理距离的高度相关性。另外2个品种霞浦元宵茶与福安大白茶,分别来自福建省内的2个不同地方,相互间的地理距离也较远。而剩余的10个品种中有9个品种来源地都与福鼎市相关,所以聚类结果呈现出遗传距离与品种来源地极强的相关性,地理距离越远遗传距离越大,反之越小。同时这3个距离最远的品种具有鲜明的品种特征,性状独特,与其它品种表型差异显著,其中早春毫叶大芽壮,芽期早,形态特征与广东凤凰水仙相似,特别是叶形与其它参试品种差别明显;霞浦元宵茶为特早生品种,几乎是目前知道的芽期最早的品种;而福安大白茶叶大肥厚,芽肥壮,毫显,混倍体,结实率极低。
剩余的10个品种聚合成2组,如图3所示,其中所有参试的半同胞系全部聚于Ⅰ组,聚类结果进一步说明半同胞系的遗传背景比对照种更为狭窄。Ⅰ组可进一步分成a,b两个亚组,该聚类结果与5个半同胞系的表型特征相吻合,聚于a亚组的福云6号与福云10号在5个半同胞系中其形态特征最为接近,都属于中小叶种,表型都偏向母本福鼎大白茶;b亚组中的福云7号、福云595及福云20号表型都偏向云南大叶种,属小乔木型,叶子肥大,芽肥壮,树形较高大。
第Ⅱ组包含福鼎大白茶与4个对照种(歌乐茶、福鼎大毫茶、九龙大白茶、早逢春)。以上5个品种除了九龙大白茶发源于福建省南平市松溪县,其它4个品种都发源于福建省福鼎市,进一步说明发源地越近遗传距离也越近。其中福鼎大白茶来源于福鼎市点头镇,小乔木型、中叶类、早生种;歌乐茶来源于福鼎市点头镇,小乔木型、大叶类、早生种;福鼎大毫茶来源于福鼎市点头镇,小乔木型、大叶类、早生种;九龙大白茶发源于南平市松溪县郑墩镇,小乔木型、大叶类、早生种;早逢春来源于福鼎市桐城镇,小乔木型、中叶类、特早生品种;以上品种都属于毫多、芽壮、生长势强的类型。但是遗传距离最近的两个品种却不来自同一地方的品种,而是来自福鼎市的福鼎大毫茶与来自南平市的九龙大白茶。其相似系数高达0.97,同时这两个品种外形也极为相似:叶大、芽壮、毫显、生长势强。但发源地距离却较远,其中原因可值得进一步研究,是否九龙大白茶来源于福鼎大毫茶的后代,或者它们有共同的亲本来源。
从图3中可看出,福鼎大白茶与其半同胞系没有聚于一组,却与发源地相同的品种聚在一起,说明福鼎大白茶遗传背景与发源于福鼎市的品种更接近,与半同胞系遗传距离更远,这是因为半同胞系遗传距离是介于福鼎大白茶与云南大叶种之间,而福鼎大白茶与云南大叶种属于不同的生态型,遗传距离远,所以半同胞系与福鼎大白茶遗传距离也离得较远。
3 讨论
3.1 福云半同胞系有较高的遗传多样性但低于对照种
一般来说 F1代的遗传多样性与父母本的遗传距离相关,并介于两亲本之间。而父母本遗传距离越远,F1代的遗传多样性越丰富。半同胞系之间的遗传多样性则比同胞系间的遗传多样性更丰富,因为父本不确定,来源广泛,遗传背景复杂,所以半同胞系间的遗传多样性要高于同胞系,出现的遗传变异更复杂,也为育种提供了更多的变异材料,更有可能选育出遗传多样性高、性状优异的品种。茶树是异花授粉植物,本实验的福云半同胞系,是福鼎大白茶与云南大叶种的天然杂交后代,父本云南大叶种属于有性群体种,相比其它纯粹自然杂交而来源各异的父本来说,云南大叶种的遗传背景相对固定。通过SSR分子标记分析发现,福云半同胞系间具有较高的遗传多样性,遗传距离在 0.75~0.91之间,但比对照种0.68~0.97要窄,这是因为对照种的亲本来源各不相同,遗传背景更复杂。但福鼎大白茶与云南大叶种由于地理距离遥远,生物学形态差异显著,遗传距离远,杂交后代显示出极强的杂种优势,选育出多个国家级、省级品种。
3.2 分子标记技术在茶树育种及栽培选种中具有重要的指导作用
通过分子标记方法分析茶树品种遗传多样性及亲缘关系,在茶树育种中具有重要的利用价值,比如在农艺性状接近的育种材料中优先挑选遗传距离较远的株系,同时在茶树育种杂交组合的亲本选配中也具有指导作用,选择遗传距离远的品种作为杂交亲本。另外,在茶树栽培选种以及引种过程中也具有指导作用,不宜过多选择遗传距离相近的品种。为了提高当地品种遗传多样性,构建良好的种群生态系统,可多选择遗传距离远的品种。
[1] 韩创举,杨培华,刘永红,等.油松半同胞家系苗期生长性状遗传分析[J].西北农林科技大学学报(自然科学版),2008,36(05):124-128,134.
[2] 杨美寅,李文东,童方平,等.湿地松半同胞家系净光合速率日变异规律研究[J].中国农学通报,2008,24(08):171-176.
[3] 马万侠,陈民生,宋尚文,等.苦楝半同胞家系苗期生长性状的研究[J].山东农业大学学报(自然科学版),2010,41(01):27-30.
[4] 张宏斌,吕东,李秉新,等.青海云杉半同胞子代苗期生长性状遗传分析[J].河北林果研究,2013,28(04):335-338.
[5] 叶乃兴.我国茶树育种的骨干亲本及其系谱分析[J].中国茶叶,2008,30(04):11-13.
[6] 余继忠,黄海涛,姚明哲,等.基于EST-SSR的福云(半)同胞系茶树品种(系)遗传多样性和亲缘关系分析[J].茶叶科学,2010,30(03):184-190.
[7] 刘振,王新超,赵丽萍,等.基于EST-SSR的西南茶区茶树资源遗传多样性和亲缘关系分析[J].分子植物育种,2008,6(01):100-110.
[8] 乔婷婷,马春雷,周炎花,等.浙江省茶树地方品种与选育品种遗传多样性和群体结构的EST-SSR分析[J].作物学报,2010,36(05):744-753.
[9] 王丽鸳,姜燕华,段云裳,等.利用SSR分子标记分析茶树地方品种的遗传多样性[J].作物学报,2010,36(12):2191-2195.
[10] 陈亮,杨亚军,虞富莲.应用 RAPD标记进行茶树优异种质遗传多态性、亲缘关系分析与分子鉴别[J].分子植物育种,2004,2(03):385-390.
[11] 刘振,姚明哲,王新超,等.基于 EST-SSR的福建地区茶树资源遗传多样性和亲缘关系分析[J].中国农业科学,2009,42(05):1720-1727.
[12] 姚明哲,刘振,陈亮,等.利用 EST-SSR分析江北茶区茶树资源的遗传多样性和遗传结构[J].茶叶科学,2009,29(03):243-250.
[13] 王丽鸳,刘本英,姜燕华,等.用 SSR分子标记研究茶组植物种间亲缘进化关系[J].茶叶科学,2009,29(05):341-346.
[14] 姚明哲,陈亮,马春雷,等.ISSR和EST-SSR标记在检测中国、日本和肯尼亚茶树品种遗传多样性上的比较分析[J].分子植物育种,2009,7(05):897-903.
[15] 周炎花,乔小燕,马春雷,等.广西茶树地方品种遗传多样性和遗传结构的EST-SSR分析[J].林业科学,2011,47(03):59-67.
[16] 王让剑,杨军,孔祥瑞,等.福建15个茶树品种SSR遗传差异分析与指纹图谱建立[J].福建农业学报,2014,29(10):970-975.
[17] 雷雨,段继华,罗意,等.2011-15-3等4个茶树新品系亲缘关系的 EST-SSR分析[J].食品安全质量检测学报,2015,6(05):1626-1632.
[18] 张成才,刘园,姜燕华,等.SSR标记鉴定浙江省主要无性系茶树品种的研究[J].植物遗传资源学报,2014,15(05):926-931.
[19] 张洁茹,韦朝领.基于4300DNA分析系统的茶树SSR发掘方法优化与建立[J].茶叶科学,2014,34(05):481-488.
[20] 章志芳,马建强.基于 SSR标记的茶树新品种遗传多样性分析及指纹图谱构建[J].湖南农业科学,2012,(19):1-4.
[21] 王会,梁月荣,刘桂芹.早生优质茶树品种资源筛选及RAPD分子标记分析[J].江苏农业科学,2009,37(03):177-179.
[22] 段云裳,姜燕华,王丽鸳,等.我国茶树主要骨干亲本及其衍生品种(系)的 SSR分析[J].植物遗传资源学报,2011,12(04):533-538.
[23] 金基强,崔海瑞,龚晓春,等.用 EST-SSR标记对茶树种质资源的研究[J].遗传,2007,29(01):103-108.
[24] 晏嫦妤,黄华林,李家贤,等.利用 RAPD分子标记对 30份山茶属种质资源进行分类分析[J].安徽农业科学,2014,42(16):4962-4963,4968.
[25] 王开荣,杜颖颖,李明,等.白化茶树特异性RAPD分子标记研究[J].浙江农业科学,2007,(01):50-55.
[26] 张毅,胡定金.EST-SSR分子标记在茶树品种鉴别中的应用[J].湖北农业科学,2012,51 (23):5406-5408.
[27] Ma, J. Q., M. Z. Yao, C. L. Ma, et al. Construction of a SSR-based genetic map and identification of QTLs for catechins content in tea plant (Camellia sinensis). PLoS One, 2014,9(3):e93131.
[28] Tan, L. Q., L. Y. Wang, K. Wei, et al. Floral transcriptome sequencing for SSR marker development and linkage map construction in the tea plant (Camellia sinensis). PLoS One,2013,8(11):e81611.
[29] Ma JQ, Huang L, Ma CL, et al. Large-Scale SNP Discovery and Genotyping for Constructing a High-Density Genetic Map of Tea Plant Using Specific-Locus Amplified Fragment Sequencing (SLAF-seq). PloS one 2015,10(6):e0128798.
Genetic Diversity of Fuyun Half-sib Teas from Fujian Determined by Using SSR Markers
CHEN Zhi-hui,SHAN Rui-yang,LIN Zheng-he,YOU Xiao-mei,ZHONG Qiu-sheng,CHEN Chang-song*
(Tea research institute of Fujian Academy of Agricultural Sciences, Fuan, Fujian 355015, China)
SSR markers were used to determine the genetic diversity of the teas, Fuyun half-sib lines, from Fujian. Eighteen pairs of SSR primers were applied for PCR amplification on 13 cultivars. There were 108 bands obtained, with 67.59% of polymorphisme. The genetic similarity coefficients of half-sib lines ranged from 0.75 to 0.91, while those of controls were between 0.68 and 0.97. Controls were richer in genetic diversity than the half-sib lines. The cluster of the 13 cultivars indicated onlya remote relationship with 3 controls, including Zaochunhao. The genetic distances among the 3 controls were also too large to be clustered in a same group. The remaining 10 cultivars were clustered into two groups, 5 half-sib lines in Group I and the female parent, Fuding dabaicha,with other 4 controls in Group II. The clustering results further confirmed a greater genetic diversity of controls as compared to the half-sib lines, and significant correlations between the genetic distance and the geographical separation, parental origin, and characteristics of the teas. The results provided information for the marker-assisted selection, cross-breeding, and genetic improvement on the tea, as well as a guide for tea cultivar selection and new variety introduction.
fuyun half-sib lines; genetic diversity; SSR molecular marker
S571.1
A
2015-09-16初稿;2015-11-03修改稿
现代农业产业技术体系建设专项资金资助(CARS-23)。
陈志辉(1975-),男,助理研究员,研究方向:茶树种质资源与遗传育种。E-mail: chenzhihui75@sina.com
陈常颂(1973-),男,副研究员,研究方向:茶树种质资源与遗传育种。E-mail: ccs6536597@163.com
猜你喜欢
环球慈善(2023年6期)2023-12-17 16:39:20
音乐天地(音乐创作版)(2023年6期)2023-10-24 09:20:38
音乐天地(音乐创作版)(2022年1期)2022-04-26 13:51:22
学生天地(2020年24期)2020-06-09 03:08:46
红土地(2019年10期)2019-10-30 03:35:06
商业文化(2016年23期)2016-05-15 05:50:46
华人时刊(2016年17期)2016-04-05 05:50:34
广西林业科学(2016年2期)2016-03-20 05:53:20
漫画月刊·哈版(2016年1期)2016-02-22 18:20:02
表面工程与再制造(2014年2期)2014-02-27 06:45:36
