
蔷薇属植物与现代月季品种杂交亲和性研究
2015-07-05 07:02赵红霞丁晓六潘会堂张启翔
西北植物学报 2015年4期
赵红霞,王 晶,丁晓六,罗 乐,潘会堂,张启翔
(花卉种质资源创新与分子育种北京市重点实验室,国家花卉工程技术研究中心,城乡生态环境北京实验室,北京林业大学 园林学院,北京100083)
现代月季为月季花(Rosa chinensis)、法国蔷薇(R.gallica)及其它几种蔷薇属植物经反复杂交、回交培育成的可以四季开花的品种类群,其花色丰富、花期长、花香迷人、株型各异,是世界上最重要的观赏植物之一[1]。目前,现代月季栽培品种有2万多个,其中80%是通过现代月季品种间杂交获得的,品种间杂交很难再引入新的遗传信息,育种潜力已非常有限[2]。同时,由于现代月季育种过程中长期重视观赏性状的提高,忽视抗逆性状的改良,导致现代月季品种的综合抗逆性越来越差[3],这些问题是现代月季品种间杂交难以解决的[4-5]。
蔷薇属植物种类丰富,参与创造现代月季品种的蔷薇属种只有10~15个[6],仅占蔷薇属种类(200多种)的1/10,中国原产的参与现代月季品种培育的蔷薇属植物也仅10种左右[2],大量的蔷薇属植物尚未应用到现代月季中,许多种类具有优良的特性,如芳香、抗寒、抗旱、耐盐碱、抗病等,对现代月季品种的进一步改良具有重要的意义[7]。将这些优异基因资源引入现代月季的最佳、最直接的途径就是用现代月季品种与野生蔷薇属植物进行远缘杂交。1867年,Guillot利用杂种长春月季和茶香月季杂交得到了第一个杂交茶香月季品种‘La France’(‘法兰西’)[8]。美国的Bryson等[9]利用抗寒性强的疏花蔷薇(R.laxa)与现代月季进行远缘杂交,杂交F1代经过3代回交,培育出能耐-38 ℃低温的聚花月季新 品 种‘Carefree Beauty’(‘无 忧 女’)。Byrne等[10]利用玫瑰(R.rugosa)、光叶蔷薇(R.wichuriana)与12个现代月季品种进行远缘杂交,培育出抗黑斑病的新品种。黄善武等[11]用弯刺蔷薇(R.beggeriana)与现代月季杂交,得到了生长健壮、抗性强的‘天山之光’、‘天香’等优良品种。马燕等[12]利用中国古老月季品种、野生蔷薇属植物[单瓣黄刺玫(Rosa xanthina f.normalis)、报春刺玫(R.primula)、木香(R.banksiae)、黄蔷薇(R.hugonis)、疏花蔷薇]与现代月季品种远缘杂交,培育出多个观赏性状优良、抗性强的品种,如‘一片冰心’、‘珍珠云’、‘春芙蓉’等。这些研究均证明野生蔷薇属植物在现代月季品种改良中具有巨大潜力。
远缘杂交往往存在不亲和性,亲缘关系越近,其获得杂种的可能性就越高,相反,若物种间亲缘关系较远,其杂交成功率就较低[13-14]。如2个亲本亲缘关系太近,则难于达到品种改良的目的。人们主要通过测定杂交结实率、果实膨大程度、有胚率、幼胚生长量等指标确定各亲本之间的杂交亲和性。张颢等[15]利用20个切花月季品种分别做母本和父本进行杂交,发现花粉生活力基本与杂交结实率成正相关,认为花粉生活力是影响月季杂交结实率的关键因素,但不是唯一因素。宋杰等[16]以二倍体的金樱子(R.laevigata)为父本与四倍体的现代月季品种杂交,认为花粉管在柱头上生长异常以及受精后胚败育是杂交不亲和的主要因素。
本试验以培育观赏性状好、芳香、抗逆性强的月季品种为目标,选11个现代月季品种和香水月季为母本,以抗逆性强的6个蔷薇属种、变种及变型和5个玫瑰品种为父本,进行远缘杂交,对亲本的花粉与柱头形态进行了扫描电镜观察,并对花粉生活力进行统计,探索不同杂交组合的结实性,分析了现代月季品种与野生蔷薇属植物及玫瑰的杂交亲和性,以期为蔷薇属植物远缘杂交亲本选配及提高远缘杂交育种效率提供依据。其中,美蔷薇(R.bella)、腺果大叶蔷薇(R.macrophylla var.glandulifera)、荷花蔷薇(R.multifloraf.carnea)、香水月季(R.odorata)均是优异的蔷薇属种质资源,但很少甚至尚未参与到现代月季品种改良工作中。前期研究发现,以野生蔷薇属植物[12]和玫瑰品种[17]为母本与现代月季品种杂交,结实率、杂种种子出苗率都很低,再加之与现代月季品种存在花期不遇的问题,所以本实验未设置以野生蔷薇属植物、玫瑰品种为母本的杂交组合。
1 材料和方法
1.1 材 料
所用杂交母本为香水月季和11个现代月季品种(‘波塞尼娜’、‘赞歌’、‘粉和平’、‘红衣主教’、‘仙境’、‘花房’、‘萨曼莎’、‘橘红潮’、‘曼海姆宫殿’、‘金玛丽’、‘红双喜’),父本为6种野生蔷薇属植物(单瓣黄刺玫、黄刺玫、报春刺玫、美蔷薇、腺果大叶蔷薇、荷花蔷薇)及5个玫瑰品种(‘大红紫枝’、‘粉紫枝’、‘紫枝’、‘白紫枝’、‘北京单瓣白’)。植株生长健壮,正常开花结实。各供试材料的基本信息详见表1。
1.2 花粉和柱头形态扫描电镜观察
在花朵尚未开放时,取父母本的花药或柱头,花药阴干散粉,柱头经0.1mol·L-1磷酸缓冲液(pH 7.34)冲洗后置于2.5%的戊二醛中固定,于4 ℃冰箱中保存,观察前利用系列乙醇溶液(30%→50%→60%→70%→80%→90%→95%→100%)脱水,每级脱水15~20min,釆用Leicaemcpd 030临界干燥仪干燥,经离子溅射仪(Eikoib-5ion Coater)喷金后在Hitachi S-3400N 扫描电镜下观察、拍照。重复3次,利用Image Pro-Plus 6.0软件测量花粉极轴长及赤道轴长、柱头表面直径、乳突直径及乳突长度,各取60个测量值,计算平均值及标准差。

表1 杂交亲本基本信息Table 1 Information of the parents used in interspecific crossing
非正常花粉率=非正常花粉粒数/花粉粒总数×100%
1.3 杂 交
课题组前期杂交试验发现一些杂交组合授粉后果实不膨大,不能获得杂交果实,加之一些杂交亲本花量不足,本试验根据材料的实际情况共设置72个杂交组合(表2)。杂交分别在2012 年5~7 月和2013年5~8月进行。在6:00~7:00采尚未开放花蕾,于室内用干净的镊子将花药捋下,在阴凉的条件下置于硫酸纸盒中,约24h后,将花粉收集并分装在离心管中保存,置于-20 ℃干燥条件下储存备用。授粉前用悬滴法测定各亲本花粉生活力,培养基为150g·L-1蔗糖+0.1g·L-1H3BO3+0.3 g·L-1Ca(NO3)2+0.2g·L-1MgSO4+0.1g·L-1K2NO3,25 ℃条件下培养24h,测定花粉萌发率(花粉管长超过花粉半径以上视为萌发)。每个品种或种重复3次,每个样品取3个视野进行观察,每个视野不少于100粒花粉,计算花粉萌发率。

表2 授粉组合Table 2 Pollination combinations in the experiment
花粉萌发率=萌发花粉粒数/花粉粒总数×100%
在下午15:00~17:00,选取母本植株上次日将要开放且发育良好的花蕾去雄,严格套袋。次日8:00~10:00进行授粉,授粉后套袋隔离,当天傍晚重复授粉[20]。
1.4 杂交结实率统计
杂交当年11月份采收果实,记录各个杂交组合的结实情况。果实采收后置于室温条件,剥出种子,观察并记录种子形态,统计种子数量,计算杂交结实率。
结实率=结实数/授粉花数×100%
1.5 种子贮藏、播种及出苗率的统计
剥出的种子经流水冲洗24h,以1∶3的比例将种子与湿润的沙子混合,置于恒温培养箱(20 ℃~24 ℃,湿度60%)中处理2周后,置于4℃冷库中贮藏45~60d,然后于50孔的穴盘中播种,采用珍珠岩和草炭(5~10mm)混合基质(体积比1∶3)。萌发长出2~3片真叶后上盆,计算出苗率。
出苗率=出苗数/播种数×100%
1.6 数据统计分析
数据用SPSS 17.0软件进行单因素方差分析,用Duncan法进行多重比较(P≤0.01)。
2 结果与分析
2.1 杂交亲本花粉形态及花粉生活力
各杂交父本的正常花粉均具三孔沟,花粉的极面观为三裂圆形,赤道面观为长椭圆形,可见1~2条萌发沟。花粉粒大小中等,极轴长(P)34.00~43.86μm,赤道轴长(E)16.01~19.20μm,种间存在显著差异,P/E值1.98~2.30。其中美蔷薇的花粉粒最小,极轴长、赤道轴长分别为34.00μm、16.01μm;荷花蔷薇花粉粒最大,极轴长、赤道轴长分别为43.86μm、19.06μm(表3)。父本材料不同程度地存在花粉败育现象,败育的花粉粒与正常花粉相比,表现为花粉粒小、畸形、内凹、空瘪、皱缩等(图版Ⅰ)。不同材料间非正常花粉率存在显著差异,黄刺玫的非正常花粉率最高,达到74.66%(表3),所有以黄刺玫为父本的杂交组合授粉后2周到1个月内果实干枯脱落,均未获得果实。其他父本材料的非正常花粉比例为15.60%~64.59%,均有杂交组合获得杂交果实。
本实验中,不同父本材料的花粉生活力差异较大,萌发率最高为单瓣黄刺玫,为89.3%,最低为黄刺玫,仅为15%(表3)。除黄刺玫外,其他10种父本材料的花粉生活力均超过30%,可以作为杂交父本。各父本的花粉生活力与非正常花粉率不存在绝对的负相关关系,但趋势基本一致。
2.2 杂交母本的柱头形态
月季柱头是干柱头,顶端具柱状凸起,柱头表面覆盖着乳突细胞(图版Ⅱ)。本实验中,各母本材料的柱头表面直径、乳突细胞长度、直径均差异显著,柱头表面直径581.91×428.76μm~981.41×873.22 μm(表4),部分母本柱头仅裂片凹陷处有凸起的乳突细胞,乳突细胞直径及长度难以测量(图版Ⅱ,5、7、8、9、11、12)。

表3 父本正常花粉的形态特征及花粉萌发率Table 3 The pollen morphology and pollen germinating rate of the male parents
2.3 杂交结实及杂种种子出苗情况
本实验共设计杂交组合72 个,杂交2 553 朵花,共有37个组合获得杂交果实(表5),35个组合未获得杂交果实(各组合授粉数量10~45 朵),43个组合未获得杂种后代。
不同的杂交组合亲和性各异,并无明显规律。根据结实率和杂种种子出苗率,筛选出14个亲和性较好的杂交组合,包括‘橘红潮’×美蔷薇、‘橘红潮’×荷花蔷薇、‘仙境’×报春刺玫、‘仙境’×腺果大叶蔷薇、‘花房’ב紫枝’、‘波塞尼娜’ב大红紫枝’、‘赞歌’×荷花蔷薇、‘萨曼莎’×美蔷薇、香水月季×单瓣黄刺玫、香水月季×美蔷薇、香水月季×腺果大叶蔷薇、香水月季×报春刺玫、香水月季ב大红紫枝’、‘粉和平’ב北京单瓣白’等,结实率均在30%以上,且均获得了杂种后代。经2年的观察发现,大部分杂交不亲和的组合在授粉后子房不膨大,1~2周内干枯脱落,如‘曼海姆宫殿’ב北京单瓣白’、‘金玛丽’ב粉紫枝’、‘花房’×黄刺玫。也有少数组合授粉后子房膨大,但在授粉后1~2月内陆续落果,如‘红双喜’×单瓣黄刺玫、‘粉和平’×单瓣黄刺玫。
不同杂交组合的单果种子数量也存在差异,其中‘赞歌’×美蔷薇组合最高,达39粒/果,‘花房’ב紫枝’组合的单果种子数最少,仅为1.61 粒/果。千粒重反映了种子的饱满程度,本实验中,相同母本的杂交组合获得的种子千粒重有差异,其中以香水月季为母本的组合最为明显,香水月季×美蔷薇组合的种子千粒重为52.11g,而香水月季ב大红紫枝’组合仅为11.74g,但千粒重与不同组合杂种种子的萌发能力没有相关性。实验发现,有些杂交亲和性好的组合获得大量杂种种子,但种子萌发率非常低,甚至不萌发,如‘粉和平’ב北京单瓣白’组合获得637 粒种子,但仅出苗13 株,出苗率仅为2.04%,香水月季×腺果大叶蔷薇组合获得117粒种子,但均未出苗。
由表5可知,荷花蔷薇、报春刺玫、美蔷薇、腺果大叶蔷薇、单瓣黄刺玫、‘大红紫枝’与所试多数现代月季品种均有较高的结实率,适合作杂交父本。黄刺玫与所有现代月季品种均不结实,不适合做父本。现代月季与玫瑰的杂交结实率普遍低于现代月季与其他蔷薇属植物的结实率,而出苗率普遍高于其他蔷薇属植物,结实率最高的组合为‘粉和平’ב北京单瓣白’(69.40%),出苗率最高的组合为‘波塞尼娜’ב大红紫枝’(52.86%)。以其它蔷薇属植物作父本时,结实率较高的组合有‘萨曼莎’×美蔷薇(84.62%),出苗率较高的组合有‘花房’×报春刺玫(21.88%)。
‘橘红潮’、‘金玛丽’、‘粉和平’、‘仙境’、‘赞歌’等现代月季品种与较多的父本杂交可以结实,适合作为杂交母本。其中,‘橘红潮’与单瓣黄刺玫、报春刺玫、美蔷薇、腺果大叶蔷薇、荷花蔷薇杂交组合的结实率分别为14.42%、23.26%、47.17%、21.74%、56.25%,但与玫瑰杂交均未获得果实。‘金玛丽’与单瓣黄刺玫、报春刺玫、美蔷薇、腺果大叶蔷薇、荷花蔷薇、‘白紫枝’杂交的结实率分别为5.66%、2.13%、3.03%、8.82%、25%、8.45%;‘粉和平’与美蔷薇、荷花蔷薇、‘大红紫枝’、‘北京单瓣白’的杂交结实率分别为5.8%、4.35%、15.6%、69.4%。
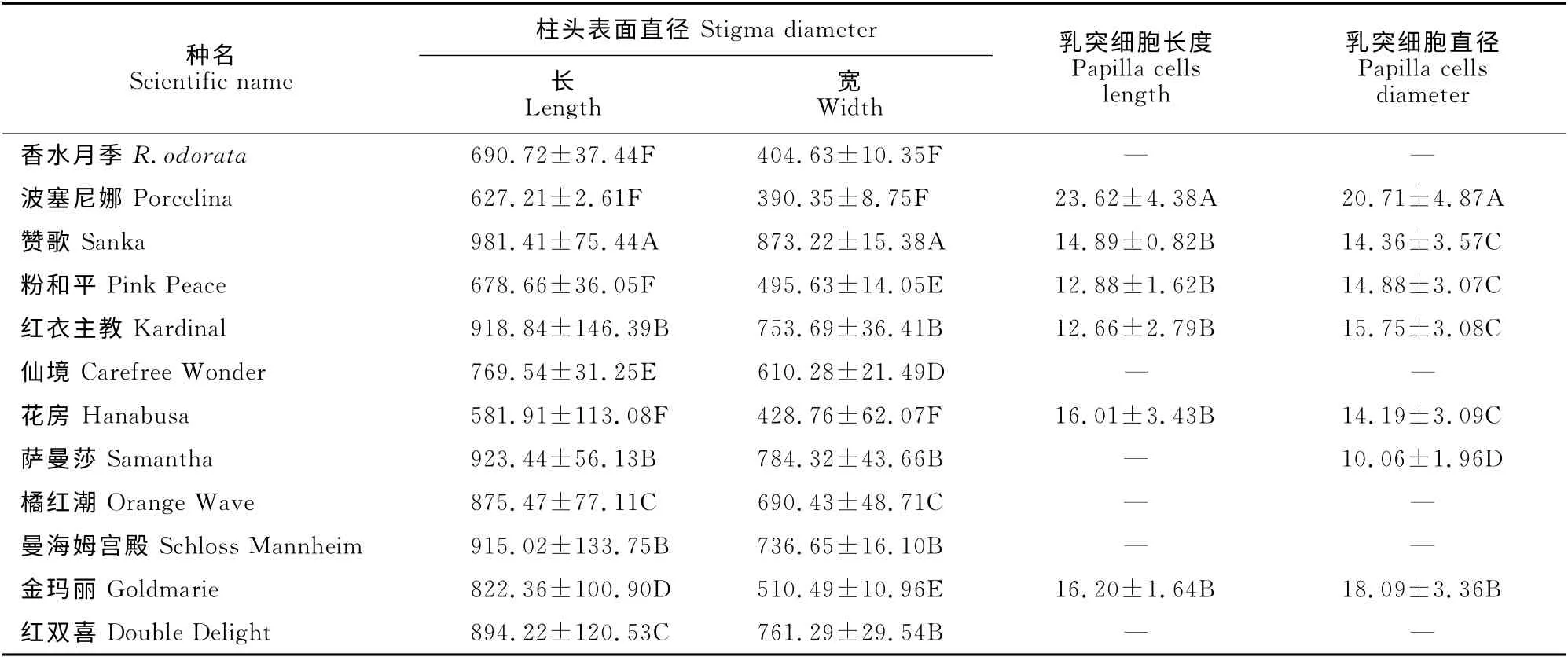
表4 杂交母本柱头的形态特征Table 4 The stigma morphology of the female parents/μm
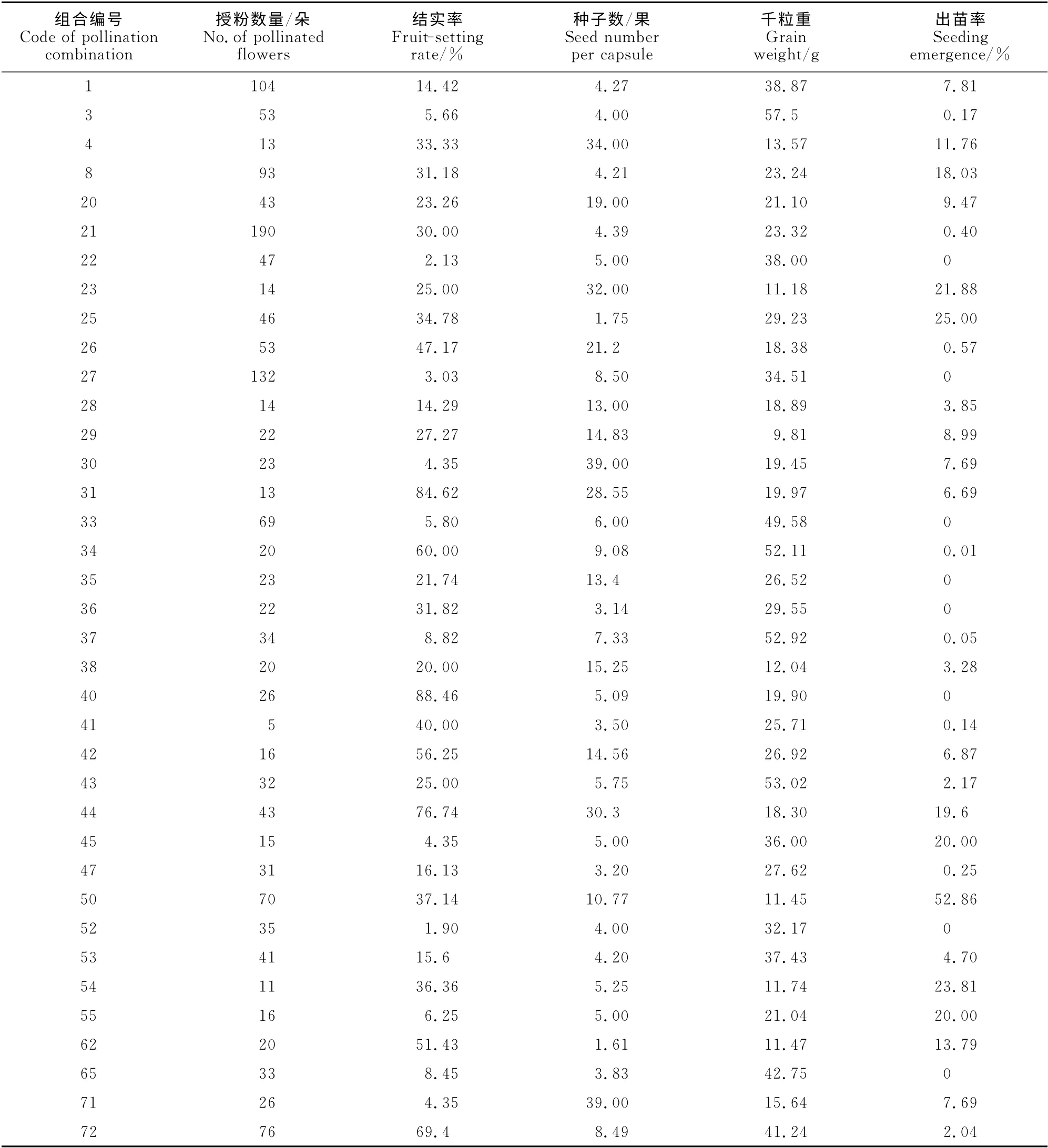
表5 37个获得杂交果实的组合结实统计表Table 5 Fruit setting and seed germination rates of 37pollination combinations
除黄刺玫外,以香水月季做母本的杂交组合均有较高的结实率,结实率最高的组合包括香水月季×腺果大叶蔷薇(88.64%)、香水月季×美蔷薇(60.00%),但这2个组合种子几乎不能萌发,杂种种子出苗率较高的组合包括香水月季×报春刺玫(25.00%)、香水月季ב大红紫枝’(23.81%)、香水月季×单瓣黄刺玫(18.03%)。
3 讨 论
远缘杂交是进行种质创新和品种改良的重要途径,在月季育种中具有非常大的应用潜力,但也存在很多问题需要解决,其中,杂交亲和性是需考虑的关键问题之一。影响杂交亲和性的因素有很多,包括花粉的生活力、花粉形态、柱头形态、亲缘关系、染色体倍性等 。
3.1 花粉生活力、花粉、柱头形态与杂交亲和性
在杂交育种的过程中,花粉生活力与杂交结实率有一定的关系。本实验中除黄刺玫外,所采用的父本材料的花粉生活力均超过30%,非正常花粉率较低,适于做授粉亲本。另外,母本自身的育性也是影响结实率的关键因子[22],柱头是接受花粉的部位,其形态影响到接受花粉、花粉萌发、花粉管伸长等过程,柱头表面的乳突形态则会影响到花粉管进入花柱[23]。对现代月季品种柱头形态的观察发现,有的柱头形成2裂片,中间有轻微的凹陷,仅在凹陷处有凸起的乳突细胞,有的柱头2个裂片非常明显,乳突细胞饱满,生长均匀。这两种形态为月季柱头发育的不同阶段,均具可授性,后者为授粉的最适阶段。本研究所涉及的蔷薇属植物花粉、柱头形态结构相似,对杂交亲和性影响不大;但不同种的花粉生活力差异较大,对杂交亲和性有一定的影响。张颢等发现切花月季品种间杂交时,花粉生活力基本与杂交结实率成正相关[15],与本实验结果基本一致。
3.2 亲本亲缘关系与杂交亲和性
本实验的父本材料中,单瓣黄刺玫、黄刺玫、报春刺玫均为芹叶组,单瓣黄刺玫、报春刺玫与现代月季品种的杂交亲和性较好,但黄刺玫与现代月季杂交存在杂交不亲和现象,这可能与黄刺玫重瓣性强、非正常花粉率高、花粉萌发率低有关系。美蔷薇、腺果大叶蔷薇、玫瑰均为桂味组,美蔷薇、腺果大叶蔷薇与现代月季杂交亲和性好,玫瑰品种中,除‘大红紫枝’外,其他品种与现代月季的杂交亲和性均较差。荷花蔷薇为合柱组,其与大部分现代月季品种均获得了杂交种子和杂交苗,杂交亲和性很好。现代月季品种与芹叶组、桂味组、合柱组的杂交亲和性不一致,可能与现代月季品种复杂而多样的遗传来源有关系。与现代月季品种不同,月季组的香水月季与单瓣黄刺玫(芹叶组)、报春刺玫(芹叶组)、美蔷薇(桂味组)、腺果大叶蔷薇(桂味组)、玫瑰‘大红紫枝’均具有较好的杂交亲和性。
本实验中,现代月季品种与玫瑰品种的杂交结实率普遍低于现代月季与其它蔷薇属植物的杂交组合,而杂种种子出苗率则普遍高于与蔷薇属植物的杂交组合。而在刘佳的实验中,现代月季品种与玫瑰品种杂交的结实率较高[17]。这可能与杂交亲本的遗传来源不同有关系,尚需进一步探讨。
3.3 亲本倍性与杂交亲和性
杂交亲本的染色体倍性相同时杂交易获得成功,所获杂种苗也更容易成活[24]。赵丹等[25]对柿(Diospyros kaki)杂交研究时,发现相同倍性的父、母本杂交亲和性大于倍性不同的父、母本。徐顺超等[26]发现不同倍性的百合品种作为亲本的杂交亲和性不同,杂交后代的倍性也不同,不同倍性百合品种间杂交存在一定程度的不亲和性。本试验中,美蔷薇、腺果大叶蔷薇为四倍体,其他父本材料均为二倍体,所用的现代月季品种均为四倍体,但现代月季品种与美蔷薇、腺果大叶蔷薇的杂交结实率并不高,甚至比与二倍体父本杂交的结实率还低。香水月季为二倍体,其与腺果大叶蔷薇、美蔷薇、玫瑰‘大红紫枝’(二倍体)、报春刺玫(二倍体)、单瓣黄刺玫(二倍体)的结实率均较高,分别为88.46%、60%、36.36%、34.78%、31.8%,香水月季与四倍体父本材料杂交的亲和性要高于与二倍体杂交。说明亲本的染色体倍性不是影响蔷薇属植物杂交亲和性的最关键因素。
本实验中,许多组合杂交亲和性较高,并获得了杂交种子,但一些组合的种子不能萌发或萌发率较低。近年来,离体胚培养技术有了很大发展,已经成为远缘杂交克服杂种胚败育的主要手段,在百合[27]、葡萄[28]等园艺植物上成功应用,为研究远缘杂交不亲和机制展示了可喜的前景。在蔷薇属杂交研究中,Kim 等[29]、林 娅 等[30]、王 丽 花 等[31]、金 菁等[32]先后利用胚挽救方法获得蔷薇属植物种间杂交后代。在下一步研究中,有必要通过胚挽救技术来克服蔷薇属植物种间杂种种子萌发率低的问题。
[1] 陈俊愉.中国十大名花[M].上海:上海文化出版社,1989.
[2] SUN X ZH(孙宪芝),ZHAO H E(赵惠恩).Review of research measures and development trend of rose breeding[J].Journal of Southwest Forestry College(西南林学院学报),2003,23(4):65-69(in Chinese).
[3] LIU Y G(刘永刚),LIU Q L(刘青林).Evaluation and exploitation of genetic resources in Roses[J].Journal of Plant Genetic Resources(植物遗传资源学报),2004,5(1):87-90(in Chinese).
[4] DE VRIES D PDUBOIS L A M.Developments in breeding for horizontal and vertical fungus resistance in rosesJ.Acta Horticulturae 2001,(552):103-112.
[5] SERGEG U.Improvement of rose varietal creation in world[J].Acta Horticulturae,1999,(495):285-291.
[6] DE VRIES D P,DUBOIS L A M.Rose breeding:past,present,prospects[J].Acta Horticulturae,1996,424:241-246.
[7] 马燕.刺玫月季育种的系统研究[D].北京:北京林业大学,1992.
[8] LIU Z M(柳子明).Roses of China and roses of Europe[J].Acta Horticulturae Sinica(园艺学报),1964,3(4):387-394(in Chinese).
[9] BRYSON S R,BUCK G J.About our cover‘Carefree Beauty’(Bucbi)rose[J].Hort Science,1979,14(2):98,196.
[10] BYRNE D H,BLACK W.The use of amphidiploidy in the development of blackspot resistant rose germplasm[J].Acta Horticulturae,1996,424:269-272.
[11] HUANG SH W(黄善武),GE H(葛 红).Studies on the use of Rosa beggeriana SCHRENK as a source of cold resistance in rose breeding[J].Acta Horticulturae Sinica(园艺学报),1989,16(3):237-240(in Chinese).
[12] MA Y(马 燕),CHEN J Y(陈俊愉).A preliminary study on breeding cultivars for establishing a new rose group-Rejuvenation Rose Group(Rj.)[J].Journal of Beijing Forestry University(北京林业大学学报),1990,12(3):18-25,125(in Chinese).
[13] 张军围.紫薇杂交亲和性及杂交后代性状变异性初步研究[D].武汉:华中农业大学,2012.
[14] CAI M(蔡 明),MENG R(孟 锐),PAN H T(潘会堂),et al.Intergeneric cross-compatibility between Lagerstroemiaand Lawsonia[J].Acta Horticulturae Sinica(园艺学报),2010,37(4):637-642(in Chinese).
[15] ZHANG H(张 颢),TANG K X(唐开学),LI SH F(李树发),et al.Cross fruitfulness of some cut flower cultivars of Chinese rose(Rosa chinensis L.)[J].Journal of Southwest Agricultural University(Nat.Sci.Edi.)(西南农业大学学报·自然科学版),2006,28(3):442-446(in Chinese).
[16] SONG J(宋 杰),WANG G L(王国良).Factors affecting fruiting rate in the crosses between Rosa laevigata Michx.and two rose cultivars[J].Journal of Nanjing Agricultural University(南京农业大学学报),2011,34(5):37-42(in Chinese).
[17] 刘 佳.玫瑰与现代月季杂交育种及杂交亲和性研究[D].北京:北京林业大学,2013.
[18] 王守超.平阴玫瑰品种分类、保存及其信息管理系统的创建[D].山东泰安:山东农业大学,2005.
[19] American Rose Society.Modern roses 12[M].Louisiana:Pediment Publishing,2007.
[20] 张佐双,朱秀珍.中国月季[M].北京:中国林业出版社,2006.
[21] PATRICIA C,BILL M.Self-and cross-compatibility relationships among genotypes and between ploidy of the rose[J].Journal of the American Society for Horticultural Science,1986,111(1):122-125.
[22] GUDIN S.Rose breeding technologies[J].Acta Horticulturae,2001,547:23-26.
[23] LIU L D(刘林德),ZHANG P(张 萍),ZHANG L(张 丽),et al.Pollen viability,stigma receptivity and pollinators of Weigela florida[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2004,24(8):1 431-1 434(in Chinese).
[24] 蔡 旭.植物遗传育种学[M].北京:科学出版社,1988:399-475.
[25] ZHAO D(赵 丹),WANG F(王 飞),JIU L J(酒立君),et al.Study on cross compatibility and fecundity of different ploidy persimmon cultivars[J].Journal of Northwest A&F University(Nat.Sci.Edi.)(西北农林科技大学学报·自然科学版),2012,40(7):141-148(in Chinese).
[26] XU SH CH(徐顺超),GUAN J(管 洁),CAO H Y(曹荷艳),et al.Study on the relationship between Lily crossbreeding affinity and ploidy levels[J].Molecular Plant Breeding(分子植物育种),2014,12(2):316-322(in Chinese).
[27] YU X Y(于晓英),WU T M(吴铁明),NI P(倪 沛),et al.In vitro of immature embryo and regeneration of plant of lily[J].Journal of Hunan Agricultural University(湖南农业大学学报),2000,26(4):286-288(in Chinese).
[28] CHENG L L(程琳琳),WANG Y J(王跃进),SHI Y(石 艳),et al.Study on factors affecting embryo rescue of seedless grape[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2007,27(7):1 317-1 322(in Chinese).
[29] KIM S W,OH S C,LIU J R.Control of direct and indirect somatic embryo genesis by exogenous growth regulators in immature zygotic embryo cultures of rose[J].Plant Cell Tissue and Organ Culture,2003,74(1):61-66.
[30] LIN Y(林 娅),LIU Q L(刘青林).Plant regeneration from immature embryos of roses[J].Journal of Beijing Forestry University(北京林业大学学报),2007,29(3):35-39(in Chinese).
[31] WANG L H(王丽花),QU S P(瞿素萍),TANG K X(唐开学),et al.Induction of hybrid plantlets of Rosa rugosa by embryo rescue[J].Journal of Southwest Forestry College(西南林学院学报),2008,28(6):53-56(in Chinese).
[32] JIN J(金 菁),WANG G L(王国良),LIU G N(刘冠楠).Research on the immature embryo rescue in Rosa distant hybridization[J].China Forestry Science and Technology(林业科技开发),2010,24(4):72-76(in Chinese).

图版Ⅰ 杂交父本的花粉形态(箭头所指为非正常花粉)1.单瓣黄刺玫;2.黄刺玫;3.报春刺玫;4.美蔷薇;5.腺果大叶蔷薇;6.荷花蔷薇;7.玫瑰‘大红紫枝’;8.玫瑰‘粉紫枝’;9.玫瑰‘紫枝’;10.玫瑰‘白紫枝’;11.玫瑰‘北京单瓣白’;12~15.败育花粉形态。PlateⅠ Pollen population view and abortive pollen morphology of male parents(Arrowheads showed the abortive pollens)Fig.1.R.xanthinaf.normalis;Fig.2.R.xanthina;Fig.3.R.primula;Fig.4.R.bella;Fig.5.R.macrophylla var.glandulifera;Fig.6.R.multifloraf.carnea;Fig.7.R.rugosa‘Dahong Zizhi’;Fig.8.R.rugosa‘Fen Zizhi’;Fig.9.R.rugosa‘Zizhi’;Fig.10.R.rugosa‘Bai Zizhi’;Fig.11.R.rugosa‘Beijing Danbanbai’;Fig.12-15.Abortive pollens.

图版Ⅱ 杂交母本的柱头形态1.‘波塞尼娜’;2.‘赞歌’;3.‘粉和平’;4.‘红衣主教’;5.‘仙境’;6.‘花房’;7.‘萨曼莎’;8.‘橘红潮’;9.‘曼海姆宫殿’;10.‘金玛丽’;11.‘红双喜’;12.香水月季。PlateⅡ Stigma morphology of female parents Fig.1.‘Porcelina’;Fig.2.‘Sanka’;Fig.3.‘Pink Peace’;Fig.4.‘Kardinal’;Fig.5.‘Carefree Wonder’;Fig.6.‘Hanabusa’;Fig.7.‘Samantha’;Fig.8.‘Orange Wave’;Fig.9.‘Schloss Mannheim’;Fig.10.‘Goldmarie’;Fig.11.‘Double Delight’;Fig.12.R.odorata.
猜你喜欢
上海农业学报(2019年5期)2019-11-08
世界热带农业信息(2019年9期)2019-01-05
铜仁学院学报(2018年7期)2018-09-08
现代园艺(2017年22期)2018-01-19
中国稻米(2017年6期)2017-12-28
西南农业学报(2017年7期)2017-08-11
山东农业科学(2017年5期)2017-06-05
上海农业学报(2017年3期)2017-04-10
上海农业学报(2017年3期)2017-04-10
数字技术与应用(2016年10期)2017-04-01
