
苹果属海棠亲缘关系的AFLP分析
2015-07-02 01:42李厚华郭亦博王亚杰
西北农林科技大学学报(自然科学版) 2015年8期
徐 曼,李厚华 ,郭亦博,赵 冰,王亚杰,高 艳
(1 西北农林科技大学 林学院,陕西 杨凌 712100;2 内蒙古阿里河林业局,内蒙古 呼伦贝尔 165450)
苹果属海棠亲缘关系的AFLP分析
徐 曼1,李厚华1,郭亦博2,赵 冰1,王亚杰1,高 艳1
(1 西北农林科技大学 林学院,陕西 杨凌 712100;2 内蒙古阿里河林业局,内蒙古 呼伦贝尔 165450)
【目的】 在分子水平上探究苹果属海棠野生种、栽培种及相近属植物之间的亲缘关系,为分类学研究、海棠育种提供参考。【方法】 选用4对AFLP引物(M-CTC/E-AGC,M-CTA/E-ACG,M-CAT/E-ACT和M-CAA/E-ACG),分析苹果属海棠9个野生种、8个栽培种以及4个山楂属植物和2个梨属植物的遗传多样性。【结果】 (1)共扩增出条带114条,其中多态性条带108条,多态位点百分率达94.7%,供试材料表现出丰富的遗传多样性。(2)供试材料在相似系数为0.73时被分为3组,分别为苹果属组、山楂属组和梨属组,因此AFLP分子标记的方法可以将苹果属与梨属、山楂属完全区分开。(3)秦巴山区野生苹果属海棠在相似性系数为0.80时被聚为4组,聚类结果支持Rehder将湖北海棠和垂丝海棠列入山荆子系的分类方法,不支持Langenfelds建立独立的湖北海棠系的分类方法。【结论】 基于遗传相似性系数的聚类结果与形态学分类结果基本一致,既验证了形态学分类的正确性,又解决了各分类系统间存在的分歧。
海棠;AFLP;分子标记;亲缘关系;分类
海棠是苹果属(Malus)多种植物和木瓜属(Chaenomeles)几种植物的通称。苹果属海棠(crabapple),是指苹果属具较小果实(直径≤5 cm)的类型。苹果属海棠既可作为重要的观赏花木,又具有食用价值和药用价值,如垂丝海棠(M.halliana)、西府海棠(M.micromalus)等具有优良的观赏性状,常用于园林绿化;湖北海棠叶中含有丰富的黄酮类物质,可制成保健茶等[1]。
以往对苹果属的分类,主要依据形态标记,如叶形、果形、嫩枝绒毛、叶缘锯齿、萼片宿存与否等[2]。由于苹果属植物具有多型性,形态特征不能充分解决苹果属种系分类问题。因此,试验研究成为苹果属植物分类与系统演化关系研究的重要依据[3]。对苹果属植物分类学的研究主要包括细胞分类学、孢粉学、植物化学分类学和分子系统学等。分子系统学作为最新的分类手段之一,常用的分析技术有RFLP、RAPD、SSR、AFLP、ITS等,其中AFLP分子标记技术是一种建立在PCR技术基础上的DNA指纹分析技术,其多态性丰富,使得从基因组水平探讨苹果属种间、组系间及与相近属间的亲缘关系成为可能[4]。AFLP标记在种下水平的遗传多样性研究中也发挥着重要作用,已被广泛用于龙眼品种(系)、梨新品种、山东石榴品种的遗传多样性及亲缘关系研究[5-7]。
运用分子技术对苹果属的分类研究主要有:梁国鲁等[8]用AFLP技术对该属23个种共31个类型的分析,印证了传统分类研究的结论,明确了佛罗伦萨海棠等疑难种的分类地位;石胜友等[9]利用AFLP标记对变叶海棠、陇东海棠、花叶海棠的亲缘关系进行分析, 结果与形态学、细胞学和同工酶的结果一致,在分子水平上揭示了变叶海棠的杂种起源;郭翎等[10]通过AFLP技术得出亚洲原产的苹果属植物对于苹果及观赏海棠品种的发展起重要作用的结论,并建议将原属花楸组植物除佛罗伦萨苹果外全部并入苹果组,取消脱萼组,将其组内的植物全部并入苹果组。
苹果属分类系统繁多,分类系统间存在差异。本研究运用AFLP分子标记技术和聚类分析方法,探究分布于秦巴山区的野生海棠种质资源、常见海棠栽培种及其相近属植物材料之间的亲缘关系,以期为分类学研究提供依据,为培育海棠新品种提供理论支持。
1 材料与方法
1.1 材 料
植物材料采集于秦巴山区、陕西杨凌西北农林科技大学博览园、北京植物园、中国科学院植物所、河北怀柔、陕西府谷。其中,山荆子、湖北海棠、垂丝海棠、三叶海棠、变叶海棠、陇东海棠、河南海棠、滇池海棠为野生种,而平邑甜茶、八棱海棠、西府海棠、花红、楸子、海棠花、海红果为常见的栽培种[11]。于2012-07采集无病虫害的幼嫩叶片,用变色硅胶干燥后,保存于-20 ℃冰箱内。供试材料编号及来源见表1。
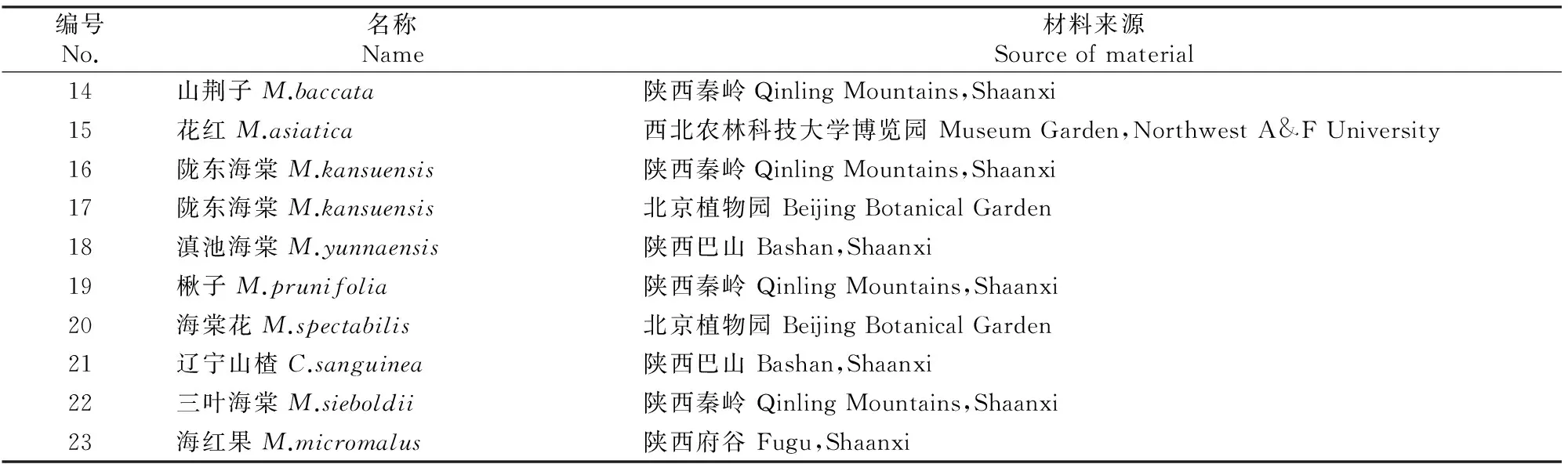
续表1 Continued table 1
1.2 方 法
1.2.1 DNA提取 采用植物基因组DNA试剂盒(天根,北京)提取各试验材料的基因组DNA。将提取的DNA用1.5 g/L琼脂糖凝胶检测纯度。采用紫外分光光度计(六一,北京),选DL2000 DNA作为对照来估计DNA浓度。之后稀释每个样品至50 ng/μL,-20 ℃保存。
1.2.2 AFLP分析 采用Vos等[12]的方法并稍作改动对样品进行AFLP分析。反应程序如下:从23个样品的基因组DNA中分别取5 μL 于37 ℃ 酶切连接3 h(内切酶采用EcoRⅠ和MseⅠ),将得到的DNA模板用于预扩增。预扩增采用MseⅠ+C/EcoRⅠ+A引物组合,预扩增程序为:94 ℃变性30 s,56 ℃复性30 s,72 ℃延伸1 min, 进行24 个循环。对预扩增的PCR产物进行10倍稀释,采用能产生清晰条带的4种引物组合对稀释的PCR产物进行选择性扩增。选择性扩增采用+3/+3引物组合,程序的第1步为13个循环(第1个循环94 ℃变性30 s,65 ℃复性30 s,72 ℃延伸1 min,接下来的12个循环,退火温度每循环减少0.7 ℃),第2步为24个循环( 94 ℃变性30 s,56 ℃复性30 s,72 ℃延伸1 min)。反应结束后在 20 μL 反应体系中加入 5 μL 加样缓冲液,95 ℃变性8 min后迅速将 PCR管放于冰上以防止复性,然后将样品置于-20 ℃冰箱保存用于电泳检测。本试验采用聚丙烯酰胺凝胶电泳和银染的方法进行检测。银染后的板晾干后在 X 线胶片观察灯下观察。
1.2.3 聚类分析 对扩增产物的电泳结果采用“0-1”系统记录谱带位置,观察电泳图谱中同一位置上DNA条带的有无,有记为1,无记为0,形成0/1矩阵图输入计算机。采用多变量分析系统(NTSYS2pc,2.10版) 软件中的DICE法计算遗传相似系数,然后根据相似系数用SAHN Clustering和非加权组平均法(UPGMA) 进行聚类分析。
2 结果与分析
2.1 AFLP扩增片段的多态性
从选择性引物组合中,筛选出扩增条带丰富、分辨能力强的引物对,分别为M-CTC/E-AGC、M-CTA/E-ACG、M-CAT/E-ACT、M-CAA/E-ACG。引物M-CAA/E-ACG组合对供试样本的AFLP扩增图谱见图1。
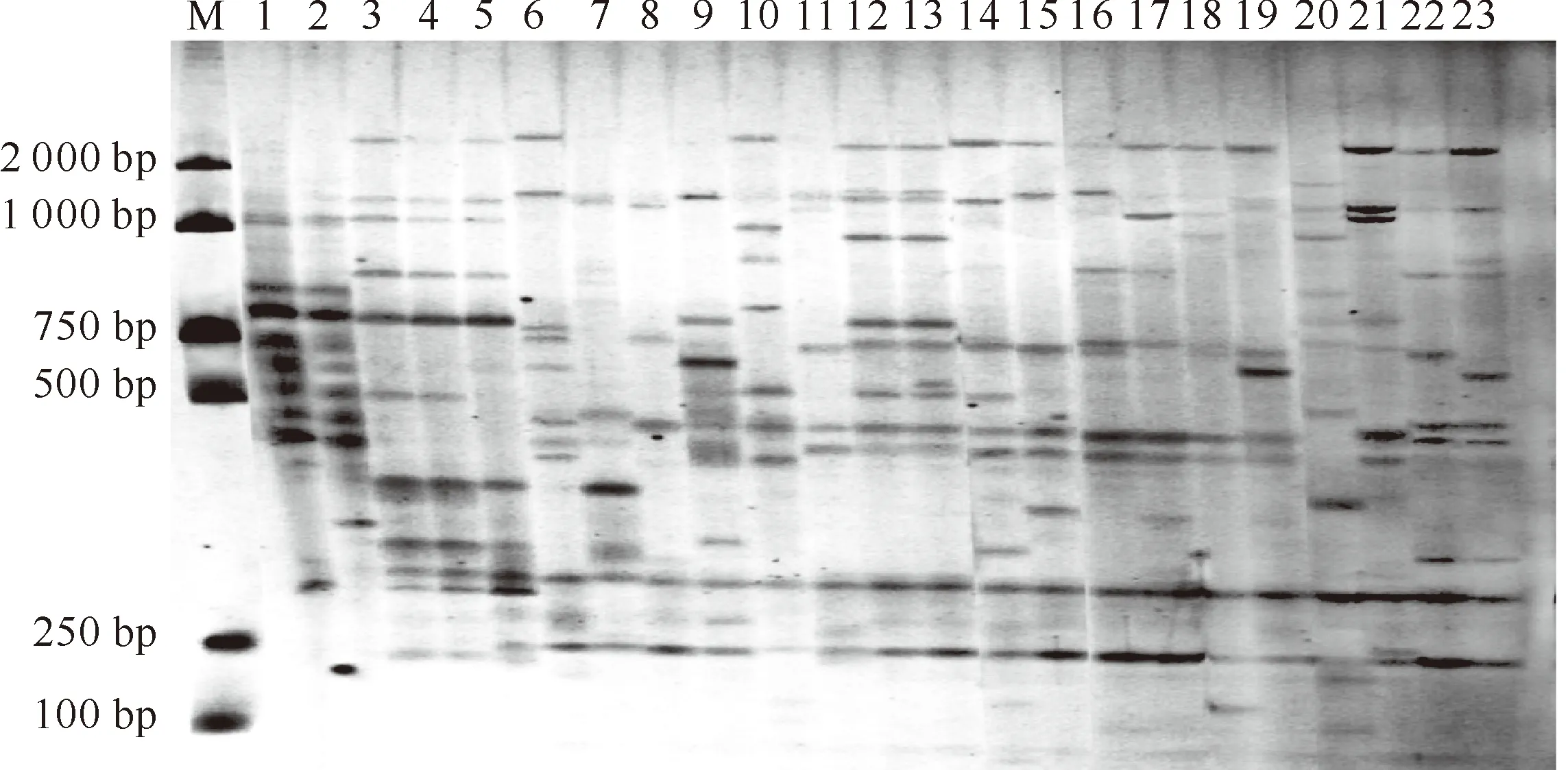
图1 引物M-CAA/E-ACG组合对供试植物样本的AFLP扩增图谱
M.Takara DL2000 DNA Marker;1~23.材料编号同表1
Fig.1 AFLP fingerprinting patterns using M-CAA/E-ACG
M.DL2000 DNA Marker (Takara);1-23.The serial numbers of samples are the same as Table 1
用这4对引物对供试材料的基因组进行扩增,共扩增出条带114条, 其中多态性条带108条。平均每对引物扩增出条带28.5条,多态性条带平均为27条,多态位点百分率达94.7%(表2)。可见,供试材料间在分子水平上酶切位点的分布具有广泛的差异,其遗传多样性极为丰富。

表2 4对AFLP选择性扩增引物产生的条带多态性Table 2 Polymorphism of AFLP bands obtained by selective amplification based on the 4 primer pairs
2.2 遗传多样性聚类分析
在用于AFLP分析的引物组合中,根据选择性扩增产物经聚丙烯酰胺凝胶电泳所得的大量清晰条带,从中选取108条多态性条带,进行遗传相似系数计算后,再经过聚类分析,构建了23份供试材料的亲缘关系树状图(图2)。这23份材料的相似系数介于0.68~0.94,根据相似性关系,在相似系数为0.73时,可以将这些材料分为3个组:Ⅰ.梨属组,包括白梨和沙梨;Ⅱ.山楂属组,包括裂叶山楂、山楂、辽宁山楂、野山楂;Ⅲ.苹果属组,包括所有的苹果属野生种以及栽培种(图2)。
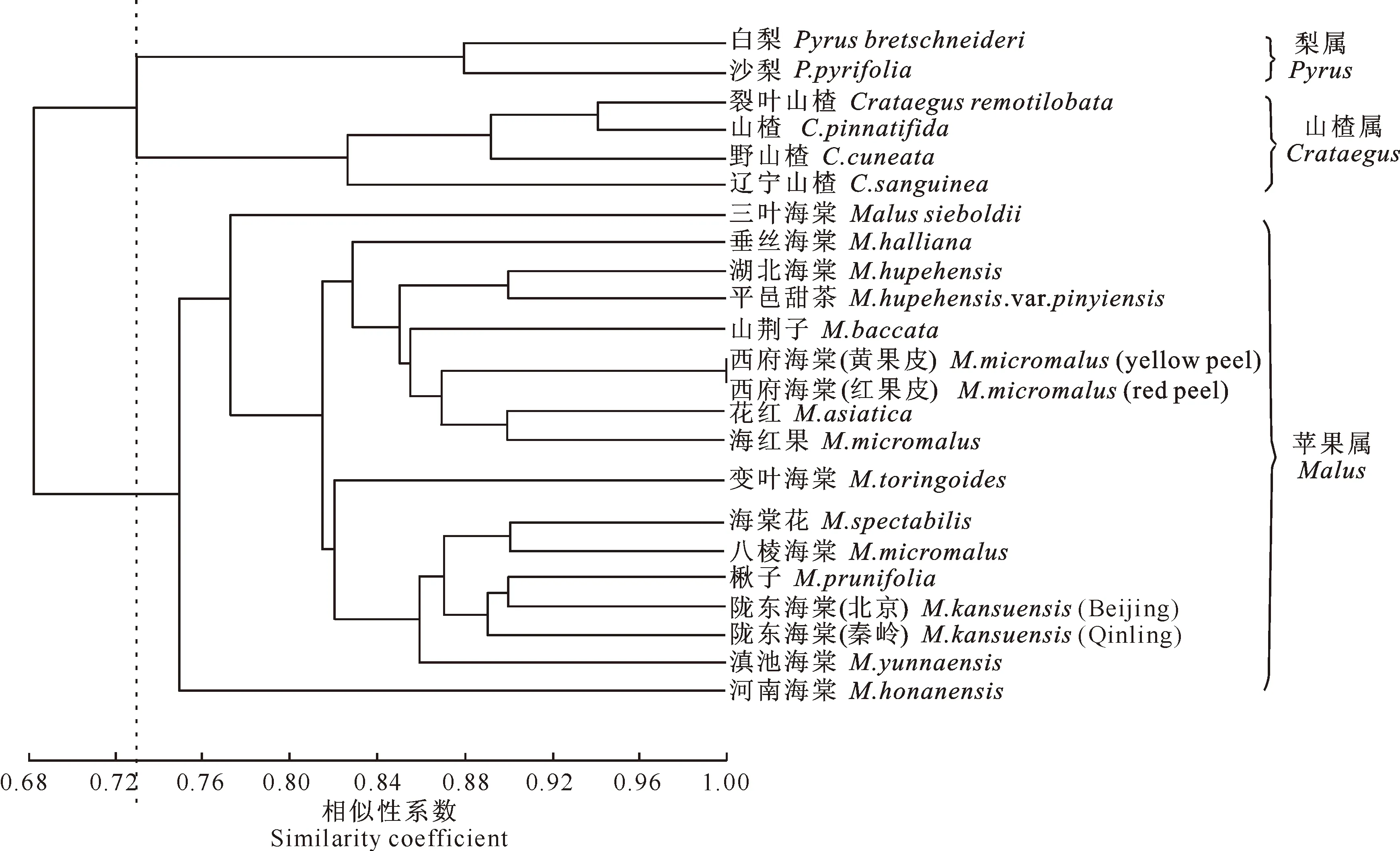
图2 23个供试植物样本AFLP分析亲缘关系树状图Fig.2 Dendrogram of 23 samples based on AFLP data
选取海棠野生种样本的相似性系数进行聚类分析,得到野生种亲缘关系树状图(图3)。在相似性系数为0.80处,可将这些采自秦巴山区的野生苹果属海棠聚为4组:Ⅰ组为山荆子系,包括湖北海棠、垂丝海棠和山荆子;Ⅱ组为三叶海棠系,包括三叶海棠;Ⅲ组为陇东海棠系,包括变叶海棠、陇东海棠;Ⅳ组为滇池海棠系,包括滇池海棠和河南海棠(图3)。
试验中的栽培种包括平邑甜茶、八棱海棠、西府海棠、花红、楸子、海棠花、海红果等,这些栽培种没有野生种的自然分布区,尽管存在着一些栽培种的亲本记载,但其亲本尚存在不确定性。试验表明,这些栽培种均可以与中国的野生海棠聚类在一起(图2)。

图3 海棠野生种AFLP 分析亲缘关系树状图Fig.3 Dendrogram of wild crabapple species based on AFLP data
3 讨 论
3.1 苹果属与梨属、山楂属的亲缘关系
苹果最早归梨属,英国植物学家Miller将Linnaeus[13]定名的苹果从梨属中独立出来[14],成立了苹果属。德国学者Borkhausen将原已定入梨属中的4个种纳入苹果属,改名为M.baccata(L.) Borkh(山荆子)、M.praecox(Pall.) Borkh(道生苹果)、M.prunifolia(Willd.) Borkh(楸子)、M.spectabilis(Air.) Borkh(海棠花)[15]。然而,根据苹果属与梨属和山楂属形态特征存在的相似性及其在全球范围分布带的多相重复性,可以推断出苹果属与梨属和山楂属存在着共祖关系[15]。目前苹果属植物出现中间杂种的事实也反映了他们之间有较近的亲缘关系。
本试验采用AFLP技术,从DNA角度对3个属的亲缘关系进行了分析,结果发现,苹果属、山楂属(裂叶山楂、山楂、辽宁山楂、野山楂)、梨属(白梨和沙梨)材料在相似性系数为0.73处被聚类在3个不同的组,表明虽然苹果属与梨属和山楂属亲缘关系较近,但用分子标记的方法可以将苹果属与梨属、山楂属完全区分开,支持了前人依据形态学特征对苹果属、梨属、山楂属分类的科学性,也进一步证明了将AFLP分子标记用于植物分类的可行性。
3.2 海棠野生种的亲缘关系
秦巴山区作为我国重要的植物种质资源分布中心,其海棠资源主要有以下几种:山荆子、湖北海棠、垂丝海棠、三叶海棠、变叶海棠、陇东海棠、河南海棠、滇池海棠[16]。根据Rehder[17]对苹果属植物的分类,这些野生种可被分为4个系:山荆子系(山荆子、湖北海棠和垂丝海棠)、三叶海棠系(三叶海棠)、陇东海棠系(变叶海棠、陇东海棠)和滇池海棠系(河南海棠、滇池海棠)。
Rehder分类系统将山荆子、湖北海棠、垂丝海棠都列在山荆子系(Ser.Baccatae),而之后Langenfelds[18]的分类系统主张将多型性发达的湖北海棠种提升为湖北海棠系(Ser.Hupehenses)。李育农[11]支持Langenfelds的主张,新建了湖北海棠系,包含湖北海棠和垂丝海棠2个种。
本研究结果表明,湖北海棠首先与山荆子聚在一起,然后二者再与垂丝海棠聚在一组,因此,根据亲缘关系远近不能将湖北海棠系(湖北海棠、垂丝海棠)与山荆子系分开。本研究结果支持Rehder将湖北海棠和垂丝海棠列在山荆子系的分类方法,不支持Langenfelds建立独立的湖北海棠系的分类方法。
本研究根据AFLP分子标记对上述野生种的聚类分析结果与Rehder的形态学分类基本一致,仅滇池海棠的聚类存在差异,滇池海棠没有与河南海棠聚在一起,反而与陇东海棠和变叶海棠聚在一起,与Rehder的形态学分类结果有所差别。但是与Huckins[19]在Rehder分类系统上进行调整的结果一致,Huckins将陇东海棠系(Ser.Kansuenses)与滇池海棠系(Ser.Yunnanenses)合并。
3.3 海棠栽培种的亲缘关系
Rehder[17]根据形态学特征推断,西府海棠可能是由山荆子和海棠花杂交而成。由本次聚类结果可以看出,西府海棠与山荆子和花红聚类在一起,而与海棠花的亲缘关系较远,因此推断西府海棠可能是由山荆子和花红杂交而成。从地理起源方面看,西府海棠起源于西府,在历史上位于陕西的宝鸡市被称作西府,而花红同样起源于西北地区,在宝鸡周围的秦岭山区有大片分布区,因此花红作为西府海棠亲本的可能更大。
在苹果属植物中,常存在同名异物和同物异名的现象,容易引起混淆,还需要进一步研究统一。根据中国植物志记载,西府海棠果皮红色[2],而陕西西府海棠果皮黄色,通过对中国科学院植物所西府海棠(中国植物志所用样本)和陕西西府海棠亲缘关系的研究发现,二者相似性系数接近于1,应为同一个栽培种。因此,建议将果皮黄色纳入西府海棠的描述中。
综上所述,利用AFLP分子标记方法进行野生植物的亲缘关系研究,补充了传统形态学分类方法的不足,得出以下结论:1)本研究可以将苹果属与梨属、山楂属完全区分开,也证明了前人依据形态学特征对苹果属、梨属、山楂属分类的科学性;2)苹果属内的聚类结果与传统的形态学分类基本一致,并且能解决不同形态学分类系统间存在差异的问题,如聚类结果支持Rehder将湖北海棠和垂丝海棠列入山荆子系的分类方法,不支持Langenfelds建立独立的湖北海棠系的分类方法;3)海棠栽培种和野生种聚类分析结果表明,秦巴山区野生海棠种质资源在海棠品种的培育中发挥着重要作用。另外,AFLP标记为遗传背景不清楚材料的父母本鉴定提供了一种可行的方法,它可以为新品种的开发和利用提供理论依据。
[1] 屈克义,胡汉环,杜远义,等.湖北海棠叶煎液药效学实验研究 [J].时珍国医国药,2000,11(2):107-108.
Qu K Y,Hu H H,Du Y Y,et al.Experiment study of pharmacodynamics ofMalushupehensisdecoction [J].Lishizhen Medicine and Materia Medica Research,2000,11(2):107-108.(in Chinese)
[2] 俞德浚.中国植物志·36卷 [M].北京:科学出版社,1974:372-398.
Yu D J.Flora of China·volume 36 [M].Beijing:Science Press,1974:372-398.(in Chinese)
[3] 钱关泽,汤庚国.苹果属植物分类学研究进展 [J].南京林业大学学报:自然科学版,2005,29(3):94-98.
Qian G Z,Tang G G.A review on the plant taxonomic study on the genusMalusMiller [J].Journal of Nanjing Forestry University: Natural Sciences Edition,2005,29(3):94-98.(in Chinese)
[4] 张 宁,沈红香,高遐虹,等.苹果属部分观赏品种与中国野生种的亲缘关系 [J].园艺学报,2007,34(5):1227-1234.
Zhang N,Shen H X,Gao X H,et al.Phylogenetic relationship between ornamental and wild species ofMalusin China [J].Acta Horticulturae Sinica,2007,34(5):1227-1234.(in Chinese)
[5] 易干军,谭卫萍,霍合强,等.龙眼品种(系)遗传多样性及亲缘关系的AFLP分析 [J].园艺学报,2003,30(3):272-276.
Yi G J,Tan W P,Huo H Q,et al.Studies on the genetic diversity and relationship of longan cultivars by AFLP analysis [J].Acta Horticulturae Sinica,2003,30(3):272-276.(in Chinese)
[6] 王 斐,林盛华,方成泉,等.梨新品种及其亲本的AFLP分析 [J].园艺学报,2007,34(4):847-852.
Wang F,Lin S H,Fang C Q,et al.AFLP analysis of new pear cultivars and their parents [J].Acta Horticulturae Sinica,2007,34(4):847-852.(in Chinese)
[7] 苑兆和,尹燕雷,朱丽琴,等.山东石榴品种遗传多样性与亲缘关系的荧光AFLP分析 [J].园艺学报,2008,35(1):107-112.
Yuan Z H,Yin Y L,Zhu L Q,et al.Genetic diversity and relationship in pomegranate (PunicagranatumL.) cultivars in Shandong revealed by fluorescent-AFLP markers [J].Acta Horticulturae Sinica,2008,35(1):107-112.(in Chinese)
[8] 梁国鲁,余 瑛,郭启高,等.苹果属植物野生种的AFLP分析及亲缘关系探讨 [C]//李育农.2002国际苹果学术研讨会论文集.泰安:山东农业大学,2003.
Liang G L,Yu Y,Guo Q G,et al.AFLP analysis and relationship of wild species of the genusMalus[C]//Li Y N.2002 International apple symposium papers set.Tai’an:Shandong Agricultural University,2003.(in Chinese)
[9] 石胜友,梁国鲁,成明昊,等.变叶海棠起源的AFLP分析 [J].园艺学报,2005,32(5):802-806.
Shi S Y,Liang G L,Cheng M H,et al.AFLP analysis of the origin ofMalustoringoidesHughes [J].Acta Horticulturae Sinica,2005,32(5):802-806.(in Chinese)
[10] 郭 翎,周世良,张佐双,等.苹果属种、杂交种及品种之间关系的 AFLP 分析 [J].林业科学,2009,45(4):33-40.
Guo L,Zhou S L,Zhang Z S,et al.Relationship of species,hybrid species and cultivars in genusMalusrevealed by AFLP markers [J].Scientia Silvae Sinicae,2009,45(4):33-40.(in Chinese)
[11] 李育农.现代世界苹果属植物分类新体系刍议 [J].果树科学,1996,13(S1):82-92.
Li Y N.A primarily modern systematics of genusMalusMill.in the World [J].Journal of Fruit Science,1996,13(S1):82-92.(in Chinese)
[12] Vos P,Hogers R,Bleeker M,et al.AFLP:A new technique for DNA fingerprinting [J].Nucleic Acids Research,1995,23(21):4407-4414.
[13] Linnaeus C.Species plantarum holmiae [M].London:Bernard Quaritch Ltd,1753:11.
[14] Miller P.The gardener’s dictionary [M].8th ed.London:John and James Rivington,1768:8.
[15] 李育农.苹果属植物种质资源研究 [M]. 北京:中国农业出版社,2001:3-145.
Li Y N.Study onMalusgermplasm resources [M].Beijing:Chinese Agriculture Press,2001:3-145.(in Chinese)
[16] 郭亦博,李厚华,张延龙,等.秦巴山区苹果属海棠资源调查分析 [J].北方园艺,2013(13):41-44.
Guo Y B,Li H H,Zhang Y L,et al.Investigation and analysis of crabapple germplasm resources ofMalusin Qinba Mountains area [J].Northern Horticulture,2013(13):41-44.(in Chinese)
[17] Rehder A.Manual of cultivated trees and shrubs [MJ].New York:Macmillan Co,1940:389-399.
[18] Langenfelds V.Appletree systema tics [M].Rija:Zinatne,1991:119-195.
[19] Huckins C A.A revision of the sections of the genusMalusMiller [J].Diss Abstr Int,1972,33(3):1031.
AFLP analysis on genetic relationship of crabapples
XU Man1,LI Hou-hua1,GUO Yi-bo2,ZHAO Bing1,WANG Ya-jie1,GAO Yan1
(1CollegeofForestry,NorthwestA&FUniversity,Yangling,Shaanxi712100,China;2ForestryAliheofInnerMongolia,Hulunbeier,InnerMongolia165450,China)
【Objective】 The study investigated the genetic relationship of wild crabapple species, popular cultivars and species of similar genus to provide theoretical support for taxonomic research and breeding new crabapple varieties.【Method】 By applying 4 pairs of AFLP (Amplified Fragment Length Polymorphism) primers (M-CTC/E-AGC,M-CTA/E-ACG,M-CAT/E-ACT and M-CAA/E-ACG),9 samples of wild crabapples from Qinling Mountains,8 samples of common crabapple cultivars and 4Crataegussamples,and 2Pyrussamples were selected for phylogenetic analysis.【Result】 (1) Four primer combinations produced a total of 114 fragments,among which 108 were polymorphic with a polymorphism ratio of 94.7%.(2) With a threshold value of 0.73,the 23 samples were clustered into 3 groups:genusMalus,CrataegusandPyrus.Molecular markers method was successful to separate genusMalus,CrataegusandPyrus.(3) With a threshold value of 0.80,the wild species of Qinling Mountains were clustered into 4 groups.The clustering result was in line with traditional morphological classification,which groupedM.hupehensisandM.hallianato SerialBaccataeRehd instead of a separate Ser.HupehensesLangenf. 【Conclusion】 Clustering based on genetic similarity coefficients was basically in line with morphological classification,which not only verified the correctness of the morphological classification,but also solved the existing differences between various classification systems.
crabapples;AFLP;molecular markers;relationship;classification
2014-01-21
国家林业公益性行业科研专项(201204308);高等学校博士学科点专项科研基金项目(20120204120006)
徐 曼(1989-),女,河南淮滨人,在读硕士,主要从事园林植物分类研究。E-mail:843338466@qq.com
李厚华(1973-),男,山东梁山人,副教授,博士,主要从事植物分子生物学研究。E-mail:lihouhua73@163.com
时间:2015-06-30 13:47
10.13207/j.cnki.jnwafu.2015.08.012
S685.99
A
1671-9387(2015)08-0159-06
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20150630.1347.012.html
猜你喜欢
作物学报(2022年2期)2022-11-06
快乐作文(1.2年级)(2021年2期)2021-09-10
儿童时代·快乐苗苗(2021年4期)2021-06-08
音乐天地(音乐创作版)(2021年11期)2021-02-18
快乐作文(1.2年级)(2019年12期)2019-09-10
快乐作文(1.2年级)(2019年8期)2019-09-10
天然产物研究与开发(2018年10期)2018-11-06
广东农业科学(2017年5期)2017-08-29
传奇故事(破茧成蝶)(2015年5期)2015-02-28
