
回交重组自交系中SSⅢ-1、SBE3和PUL基因对稻米蒸煮食味品质的影响
2015-06-28 16:54许顺菊向珣朝康翠芳龙小林苏文丽杨博文吴家富
西北植物学报 2015年10期
许顺菊,向珣朝,康翠芳,龙小林,苏文丽,杨博文,吴家富
(西南科技大学植物分子遗传育种实验室,四川绵阳621010)
回交重组自交系中SSⅢ-1、SBE3和PUL基因对稻米蒸煮食味品质的影响
许顺菊,向珣朝*,康翠芳,龙小林,苏文丽,杨博文,吴家富
(西南科技大学植物分子遗传育种实验室,四川绵阳621010)
该研究利用颗粒结合淀粉合成酶基因(Wxb)与可溶性淀粉合成酶Ⅱa基因(SSⅡ-3)均相同的籼型光温敏核不育系‘广占63S'和潜力恢复系CG173R为亲本,经过回交和多代自交,以构建的回交重组自交系(BILs)BC1F10代株系为供试材料,分析各株系的基因型组成及其蒸煮食味品质(ECQs)和RVA谱,以解析与品质相关微效基因的遗传效应。结果显示:(1)双亲在焦磷酸化酶大亚基基因(AGPlar)、分支酶基因Ⅲ(SBE3)、脱分支酶基因(PUL)、可溶性淀粉合成酶Ⅰ基因(SSⅠ)和可溶性淀粉合成酶SSⅢ-1基因(SSⅢ-1)的基因位点存在差异。(2)SSⅢ-1、SBE3和PUL基因分别在BC1F10代株系中存在单基因分离,SSⅢ-1和SBE3基因、SSⅢ-1和PUL基因在BC1F10代株系中均存在双基因的分离。(3)不同基因型及其互作与蒸煮食味品质(ECQs)中的表观直链淀粉含量(AAC)、胶稠度(GC)以及RVA谱的部分特征值存在显著或极显著的效应。(4)SSⅢ-1单基因只对AAC有极显著影响;SBE3基因和SSⅢ-1基因互作对AAC有极显著影响,对峰值时间(PeT)、成糊温度(Pa T)、GC有显著性影响;PUL基因和SSⅢ-1基因互作对PeT、PaT和回复值(CSV)有极显著影响,对最高粘度(PKV)、热浆粘度(HPV)、崩解值(BDV)、冷浆粘度(CPV)、消减值(SBV)、AAC和GC有显著影响。研究表明,在Wxb和SSⅡ-3基因背景下,参与淀粉合成的微效基因SSⅢ-1与SBE3和SSⅢ-1的互作效应极显著影响水稻AAC,SBE3和SSⅢ-1的互作效应与PUL和SSⅢ-1的互作效应显著影响GC,PUL和SSⅢ-1的互作效应极显著影响PaT,这些发现将对改良稻米品质和加快水稻优质育种具有重要意义。
水稻(Oryza sativa L.);回交重组自交系(BILs);SSⅢ-1基因;SBE3基因;PUL基因
水稻是以淀粉为主的粮食作物之一。近几十年来,水稻产量得到了大幅度提高,稻米品质依旧普遍较低,使得育种研究者更加关注稻米品质的改良。蒸煮食味品质(eating and cooking qualities,ECQs)是稻米品质的主要指标之一,通常用3个理化特性来衡量:表观直链淀粉含量(apparent amylose content,AAC)、胶稠度(gelconsistency,GC)和糊化温度(gelatinization temperature,GT)。稻米淀粉粘滞性谱(rice starch viscosity,RVA profile)反映的是稻米淀粉与一定量的水混合后的米浆在加热、高温和冷却等条件下粘度不断变化而呈现的糊化曲线,可以比较灵敏地反映不同水稻品种间淀粉的品质差异[1],因此稻米淀粉的粘滞特性也是反映食味品质的重要指标之一。稻米品质很大程度上取决于淀粉的品质,稻米淀粉可分为直链淀粉和支链淀粉,支链淀粉的结构及两类淀粉所占比例共同决定了稻米淀粉品质[2]。因此,深入研究稻米淀粉合成相关基因对稻米理化特性的影响是开展淀粉品质遗传改良的基础。
直链淀粉的合成主要由颗粒结合淀粉合成酶基因(Wx)调控。支链淀粉的生物合成过程相当复杂,由可溶性淀粉合成酶基因(SSS)、分支酶基因(SBE)和脱分支酶基因(DBE)协同调控合成[3-4],每一类酶又有多个同工型(isoform)。近年来,稻米淀粉合成相关基因(starch synthesis-related genes,SSRGs)对理化指标和RVA特征值的影响已经有大量的报道,表明SSⅡ-3(可溶性淀粉合成酶基因)是调控GT的主效基因[5-9],而GC、AAC以及大部分的RVA谱特征值主要由基因Wx调控[10-15]。已有的研究表明,控制各理化指标的主效基因相同的品种,其品质指标却存在着很大差异[16],表明稻米淀粉合成不仅由上述主效基因调控,其他淀粉合成相关基因也参与其中,因此,也应当重视研究除主效基因以外的其他淀粉合成相关基因对稻米品质的影响。
可溶性淀粉合成酶(soluble starch synthase,SS)主要存在于质体的基质中,在叶片和贮藏器官中有多种同工型[17-20],其与分支酶一起参与支链淀粉的合成。水稻可溶性淀粉合成酶包括SSⅠ、SSⅡ、SSⅢ和SSⅣ。SSⅢ-1和SSⅢ-2是SSⅢ基因的2种同工型,SSⅢ-1主要是在叶片中特异性表达[21-22]。与SSⅢ-1相比,SSⅢ-2与稻米品质的关系更密切,但是王芳等研究表明SSⅢ-1对稻米ECQs及RVA谱同样有影响。SBE3(SBEⅡb)是淀粉分支酶(starch branching enzyme,SBE)基因的1种,Nishi等研究发现水稻和玉米的直链淀粉扩展体(ae)属于SBE3功能缺失突变体,RBE3(SBE3家族)的活力减弱,或消失可能引起突变植株中直链淀粉含量的提高。同时他们发现RBE3突变引起支链淀粉DP<17的分支降低,特别是8<DP<12的分支比例[23]。PUL(即极限糊精酶基因)是脱分支酶(debranching enzyme,DBE)基因的1种,在水稻基因组中,只存在1个拷贝的PUL基因[24]。Kubo A等[25]和Fujita N等[26]研究证实PUL不仅参与水稻胚乳中的淀粉降解过程,而且在淀粉的合成过程中也发挥有一定作用。PUL基因发生突变后,胚乳中支链淀粉聚合度小于13的短链明显增加,而B2-3链的平均链长要比野生型增加约3个葡萄糖残基,说明PUL基因对支链淀粉的精细结构起重要作用[26]。因此,研究SSⅢ-1、SBE3和PUL基因对稻米品质的影响有重要意义。
康翠芳等[27]研究了水稻淀粉合成相关基因SSⅠ、SSⅢ-1和PUL对稻米品质的影响,表明PUL基因在后代分离过程中,不同基因型之间,GC、CSV具有显著性差异。SSⅢ-1和SSⅠ基因的互作效应对GT的效应达极显著水平,对AAC、GC、PKV、SBV的效应达显著水平。由于实验样本数较少,仅给出了平均值,未能对不同基因型组合品质性状的平均值作方差分析。基于上述工作基础,本研究仍然利用以Wxb和SSⅡ-3主效基因相同,AGPlar、SBE3、PUL、SSⅠ和SSⅢ-1基因有多态性的双亲构建的BILs高世代株系为研究材料,增加样本数量,分析非糯水稻中SSⅢ-1、PUL和SBE 3基因在后代中单基因分离、双基因分离以及双基因互作对稻米ECQs的影响,解析上述3个基因的主要作用因子及其功能,以期为稻米品质改良提供更充分的理论依据。
1 材料与方法
1.1 供试材料
水稻籼型(Oryza sativa ssp.indica)光温敏核不育系‘广占63S'和潜力恢复系CG173R[广恢128 ×BC2﹛[(蜀恢527×Kitaake)×蜀恢527]蜀恢527﹜,即蜀恢527与粳稻Kitaake杂交,F1用蜀恢527做轮回亲本再回交2代后,选优株做父本与广恢128做母本再杂交于F7代稳定,测恢后定名]杂交得到F1代,与潜力恢复系回交产生的BC1F1,通过单株选择构建的BC1F10株系作为实验材料,属于回交重组自交系(backcross inbred lines,BILs)。
利用田志喜等[28]开发的19对SSRGs的分子标记和蔡秀玲等[29]开发的分子标记PCR-AccI,检测双亲‘广占63S'和CG173R在淀粉合成相关基因位点上的基因型。双亲Wxb和SSⅡ-3基因型相同,AGPlar、SBE3、PUL、SSⅠ和SSⅢ-1基因型不同。2014年4月将这些基因型有多态性的单株种植于西南科技大学农学实验基地,当年8月底收获种子,得到4个BC1F10株系:株系S10共15株、株系S134共17株、株系S63共29株和株系S52共19株。对BILs不同株系分单株取样,稻谷经过电动砻谷机(日本Kett,型号TR-200)脱壳为糙米,通过实验用小型精米机(Pearlest,日本Kett)磨成精米后,进一步经高速锤式通用粉碎磨(瑞典Perten,Lm3100型)磨成米粉,过100目筛后放置1~2个月当含水量达到12%时测定稻米ECQs。
1.2 亲本及各株系基因型检测
对BILs不同株系分单株取样,每个单株取适量叶片,经MP Fastprep样品快速破碎系统(美国MP medicals)处理,按微量SDS法提取基因组DNA[30],重复2次。
利用田志喜等[28]开发的分子标记检测亲本淀粉合成相关基因的基因型。利用蔡秀玲等[29]开发的分子标记PCR-AccI检测Wx基因第一内含子+1位碱基类型。这些用于检测的分子标记的碱基序列见表1,PCR扩增程序参见田志喜等[28]和蔡秀玲等[29]。
PCR反应在BIO RAD DNA Engine Dyad Peltier Thermal Cycler上运行,PCR反应为10μL体系:2×Reaction Mix(含Mg2+)5μL,5 U/μL Taq DNA Polymerase 0.3μL,1 umol/L引物各1μL;25 ng/μL DNA模板1μL;dd H2O 1.7μL。所有药品和试剂购置于成都市博瑞克生物技术有限公司和天根生化技术科技(北京)有限公司。
分子标记SSⅢ-1M1和PUL M2扩增程序为:94℃预变性4 min;94℃变性1 min,58℃退火30 s,72℃延伸30 s,45个循环;最后72℃延伸10 min,PCR产物保存于4℃,取产物3%琼脂糖电泳。
分子标记SBE3 M2扩增程序为:94℃预变性4 min;94℃变性1 min,58℃退火1 min,72℃延伸1 min,45个循环;最后72℃延伸10 min,PCR产物保存于4℃,取产物3%琼脂糖电泳。
1.3 表观直链淀粉含量与胶稠度的测定
按国家标准GBT15683-1995[31]测定各试样稻米粉的AAC和GC,每个指标重复测定3次。
1.4 RVA谱粘滞性的测定
采用配套软件为TCW(Thermal Cycle for Winows,4500型)的粘度速测仪(澳大利亚Newport Scientific仪器公司)来测定稻米淀粉粘滞性谱(RVA谱)。按照AACC操作规程[32],稻米粉含水量为12.0%时,取样3.0 g和蒸馏水25.00 m L加于罐内。仪器开始运行后,搅拌器在起始10s内以960 r/min转动,之后保持在160 r/min;罐内温度变化如下:50℃保持1 min,以12℃/min升温到95℃,95℃保持2.5 min,再以12℃/min降温到50℃,50℃保持1.4 min。粘度单位是随机单位(RVA arbitrary unit,RVU)。重复测定3次。
1.5 统计方法
研究所得数据采用Excel 2003和DPS 9.50统计软件进行单因素方差分析,根据基因型检测结果对数据进行分类,利用DPS 9.50进行二因素裂区试验统计分析不同基因在不同基因型下AAC、GC及RVA谱特征值的变化特征。

表1 水稻淀粉合成相关基因的分子标记Table 1 Molecular markers used to amplify starch synthesis-related genes in rice
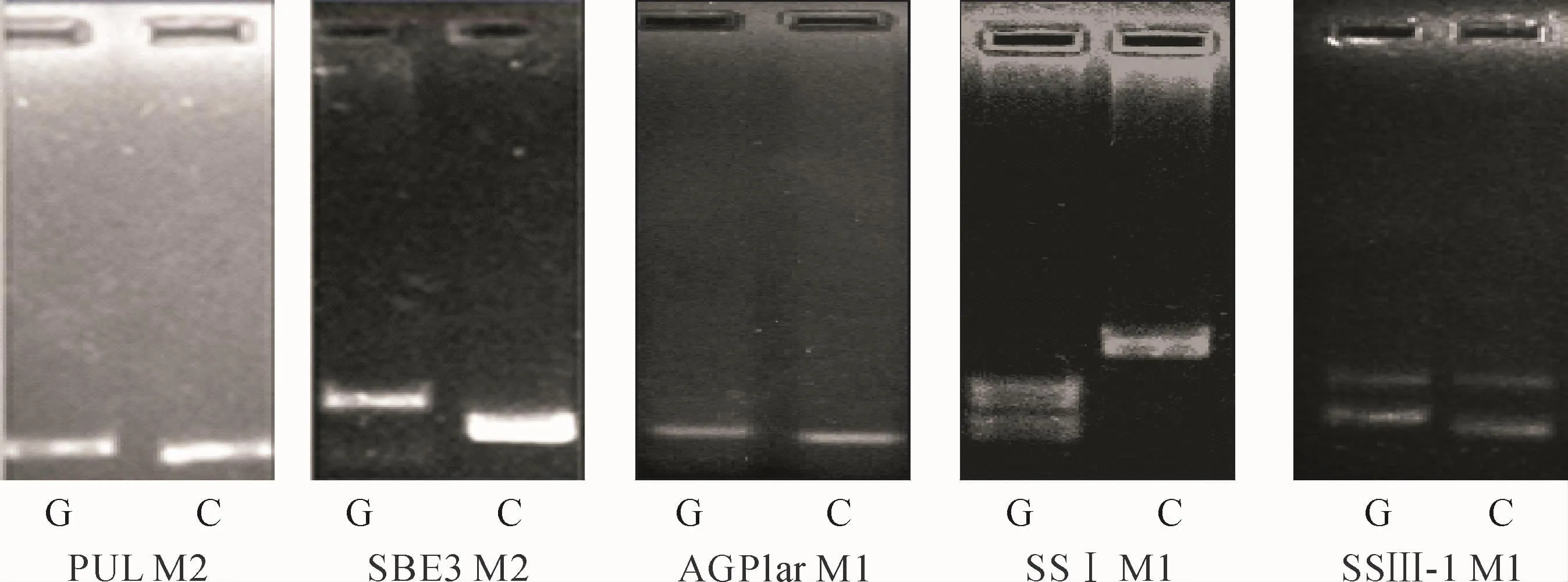
图1 差异分子标记的检测结果G.广占63S;C.CG173RFig.1 The testing results of different molecular markers with polymorphism in parents G.Guangzhan 63S;C.CG173R
2 结果与分析
2.1 BC1F10株系各单株的基因分型
利用上述5个品质相关基因位点的分子标记,对BC1F10后代株系基因型进行检测,双亲有差异的分子标记检测结果见图1。BC1F10后代株系中的各单株在某一基因位点上的等位基因与亲本‘广占63S'一致的基因型即为为广占63S型,与亲本CG173R一致的即为CG173R型,等位基因杂合的基因型即为杂合型。各单株在SSⅢ-1位点的基因型为广占63S型、CG173R型及杂合型时分别标记为G型、C型和H型。单株在SBE3位点基因型为广占63S型、CG173R型及杂合型时分别标记为Ⅰ型、Ⅱ型和Ⅲ型。单株在PUL位点基因型为广占63S型、CG173R型以及杂合型时分别标记为1型、2型和3型。
BC1F9中有2个单株在Wxb、SSⅡ-3、AGPlar、SSⅠ、SBE3和PUL基因位点的基因型相同,均只在SSⅢ-1位点发生了单基因分离。因此,这2个单株的种子在BC1F10种成株系得S10和S134。2个株系中各单株在SSⅢ-1位点存在3种基因型分别为G型、C型和H型。另外,不分离的基因中除SBE3基因外,其余各基因位点基因型均一致,株系S10中SBE3的基因型为Ⅱ型,株系S134中SBE3基因型为Ⅰ型,株系S10中SSⅢ-1基因分离情况为:5株为G型、7株为C型、3株为H型;株系S 134中SSⅢ-1基因分离情况为:5株为G型、7株为C型、5株为H型;BC1F9中有1个单株在Wxb、SSⅡ-3、AGPlar、SSⅠ和PUL基因位点的基因型相同,在SSⅢ-1和SBE3位点同时存在分离,在BC1F10种成株系得S63。该株系在SBE3位点存在2种基因型为:Ⅰ型和Ⅱ型;SSⅢ-1位点存在3种基因型为:G型、C型和H型。SBE3基因在株系S63中分离情况为:12株为Ⅰ型、15株为Ⅱ型;SSⅢ-1基因分离情况为:5株为G型、11株为C型、3株为H型;BC1F9中有1个单株在Wxb、SSⅡ-3、AGPlar、SSⅠ和SBE3基因位点的基因型相同,在SSⅢ-1和PUL基因位点同时存在分离,在BC1F10种成株系得S52。该株系在PUL位点存在2种基因型为:1型和2型,SSⅢ-1位点存在2种基因类型为:C型和H型。PUL基因分离情况为:5株为1型、11株为2型。SSⅢ-1基因分离情况为:5株为C型、11株为H型。结果列于表2。图2列出部分检测结果,表明供试单株发生分离的基因可分为3种基因类型:广占63S、CG173R型和杂合型。
2.2 各基因对稻米品质的效应分析
2.2.1 SSⅢ-1基因对稻米品质的影响 由表3可知,株系S10和株系S134中的AAC,在SSⅢ-1各基因型间存在显著性差异,但不同株系的AAC排列均值大小不同。株系S10中3种基因型的AAC均值大小依次为C型<H型<G型,H型样本的AAC介于两亲本类型之间,G型的AAC高于C型;株系S134中3种基因型的AAC均值大小依次为H型<C型<G型,H型样本的AAC最小,G型的AAC最高。二者AAC均值大小顺序不同,可能原因是由于两个株系在SBE3位点的基因型不同,使得SBE3和SSⅢ-1基因间因相互作用而导致的。其他的理化指标及RVA特征值在不同的SSⅢ-1基因型之间未达到显著性水平,说明SSⅢ-1基因的分离在该株系中对RVA谱各特征值以及对GC的影响较小。

表2 后代BC1F10株系在各基因位点的遗传表现Table 2 The genetic characteristics of different gene loci in BC1F10lines
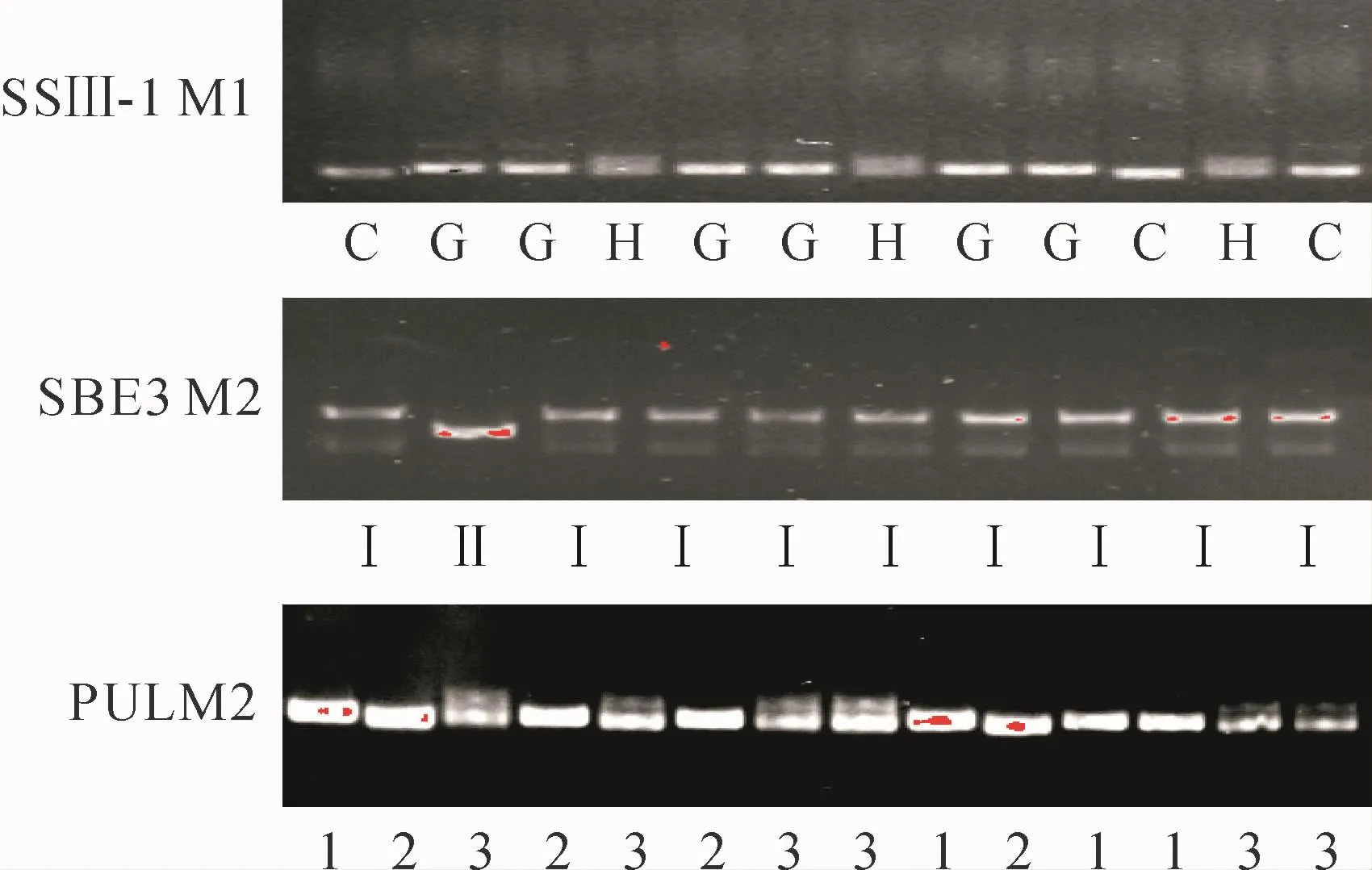
图2 分子标记在部分供试材料中的多态性G.广占63S;C.CG173R;H.广占63S和CG173R杂合Fig.2 Polymorphism of molecular markers in partial materials tested G.Guangzhan63S;C.CG173R;H.Heterzygote of Guangzhan 63S and CG173R
对株系S10和S134混合成一个大群体进行分析,由表3可知,SSⅢ-1基因在AAC上,各基因型间达到了极显著性差异,3种基因型的AAC均值大小依次为C型<H型<G型,G型的AAC高于C型。其他的理化指标及RVA特征值在不同的SSⅢ-1基因型之间未达到显著性水平。由此可知SSⅢ-1基因的分离对AAC有极显著影响,而对RVA谱各特征值和GC的影响均较小。
2.2.2 SBE3和SSⅢ-1基因对稻米品质的影响
(1)SBE3和SSⅢ-1基因同时分离时单基因对稻米品质的影响 由表4可知,SBE3不同的基因型效应除PKV外,在AAC和GC及其他的各理化指标上均达到显著或极显著差异水平。其中在HPV、CPV、CSV、SBV、PaT、Pe T和GC上达到极显著水平,在BDV和AAC上达到显著水平。其中在HPV、CPV、CSV、SBV和Pe T上Ⅰ<Ⅱ;在PKV、BDV、Pa T、Pe T、AAC和GC上Ⅱ<Ⅰ。SSⅢ-1不同的基因型效应除PKV和AAC外,在GC及其他的各理化指标上均达到显著或极显著差异水平。其中在HPV、CPV、CSV、SBV、Pa T、PeT和GC指标上达到极显著水平,在BDV上达到显著水平。在PKV、BDV、Pa T和GC上G<C<H,在HPV、CPV、CSV、SBV和Pe T上H<C<G,在AAC上C<H<G。表明在相同Wxb基因表达的非糯水稻中SSⅢ-1和SBE3基因对稻米蒸煮食味品质有显著影响。由于SSⅢ-1和SBE3基因在株系S63中同时存在基因分离,可能存在两者之间的互作效应。
(2)SBE3和SSⅢ-1基因的互作效应分析 为探讨SSⅢ-1和SBE3基因间的互作效应,根据株系中两基因不同的基因型组合方式,将此株系参试材料分为5类(表5),这些不同SSⅢ-1和SBE3基因型组合的稻米品质的均值及差异显著性见表5。由表5可知在不同的组合方式下HPV和AAC在各基因型间存在显著差异;CPV、SBV、PeT、Pa T、CSV和GC在各基因型间存在极显著性差异。在不同SSⅢ-1基因型的遗传背景下各理化指标BDV、Pa T和GC中SBE3基因型为Ⅰ型的值均大于Ⅱ型,而HPV、CPV、SBV、PeT和CSV中SBE3基因型为Ⅱ型的值均大于Ⅰ型。说明SSⅢ-1和SBE3基因存在相互作用。
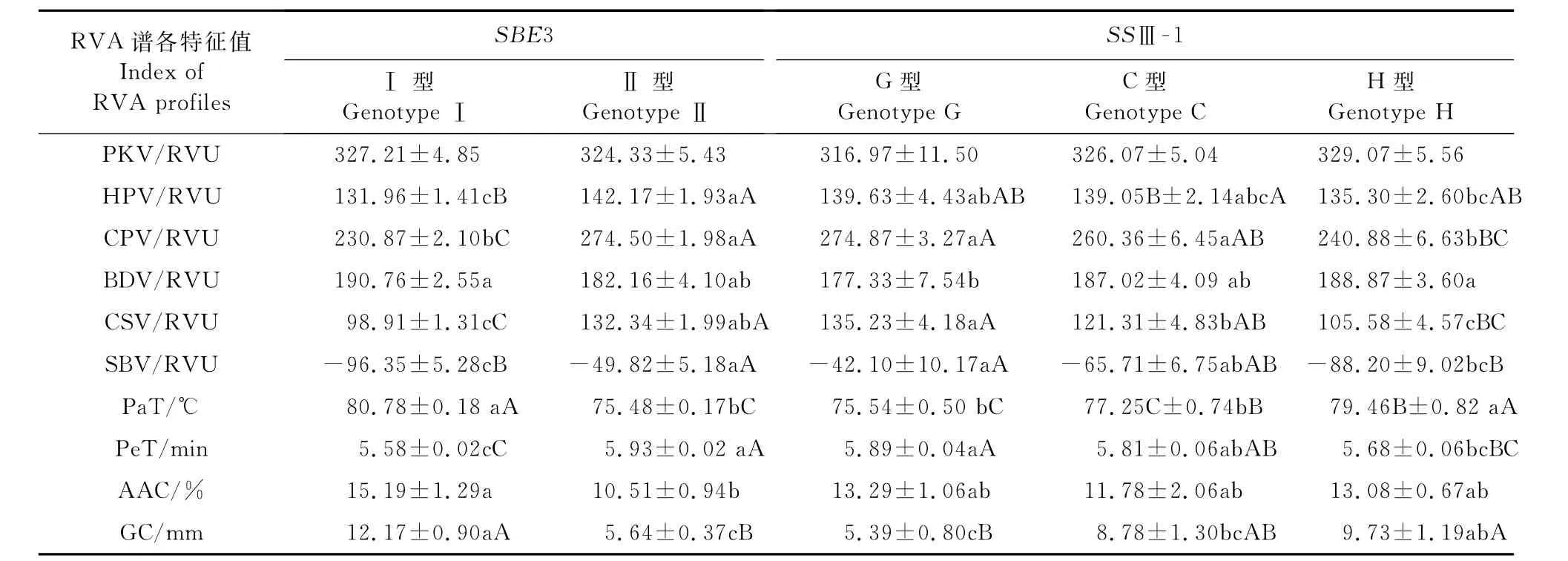
表4 SBE3和SSⅢ-1不同基因型稻米品质性状的均值及差异显著性Table 4 The means of quality traits and their significant test of different SBE3 and SSⅢ-1 genotypes
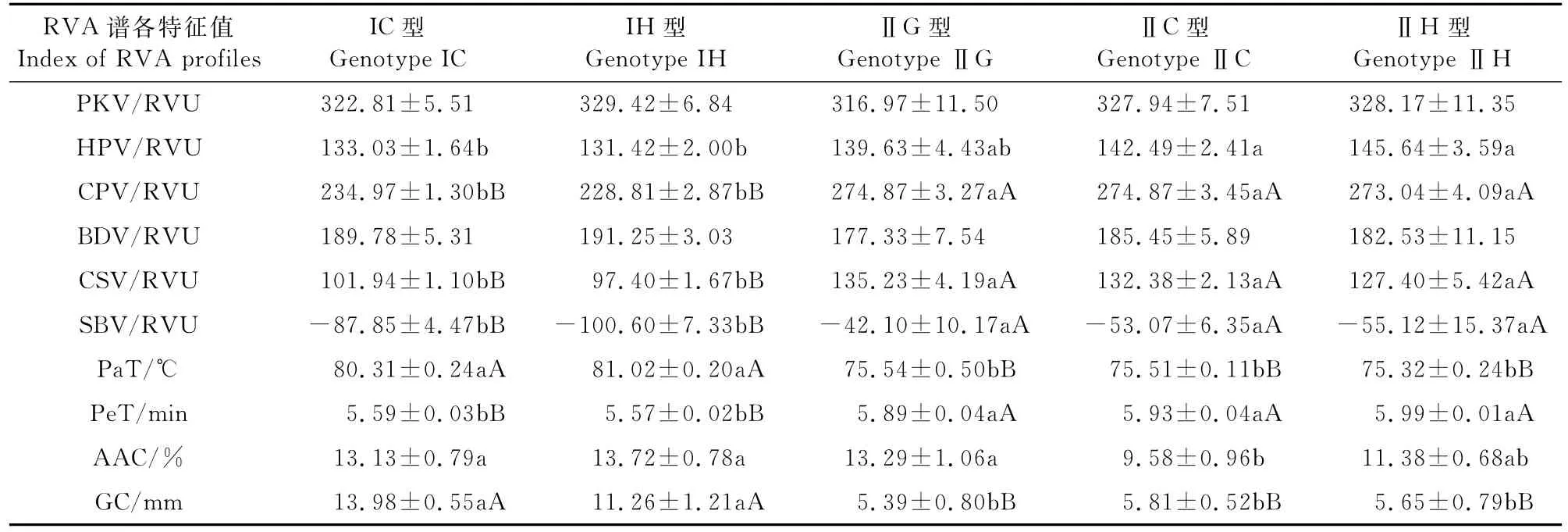
表5 不同SSⅢ-1和SBE3基因型组合稻米品质的均值及差异显著性Table 5 The means of quality traits and their significant test of different combinations of SBE3 and SSⅢ-1 genotypes
进一步将SBE3基因型作为主因素,SSⅢ-1基因作为副因素,进行裂区试验分析,探究SBE3基因和SSⅢ-1基因的互作效应,结果见表6。由表6可知,SBE3基因对HPV、BDV和AAC的效应达到极显著水平。SSⅢ-1基因对SBV、Pa T、CSV、AAC的效应达显著性水平。SBE3基因和SSⅢ-1基因的互作对AAC的效应达到极显著水平,对Pe T、Pa T和GC的效应达到显著水平。
2.2.3 PUL和SSⅢ-1基因对稻米品质的影响
(1)PUL和SSⅢ-1基因同时分离时单基因对稻米品质的影响 株系S52中各单株在Wxb、SSⅡ-3、AGPlar、SSⅠ和SBE3基因位点的基因型相同,在SSⅢ-1和PUL位点同时存在分离。由表7可知,不同的PUL基因在SBV和Pe T上存在显著性差异,而且在SBV指标上其均值为2<1,在Pe T指标上其均值为1<2。不同的SSⅢ-1基因同样在SBV和Pe T上存在显著性差异;在SBV上其均值大小为H<C,在PeT上其均值大小为H<C。由于SSⅢ-1和PUL基因在株系S52中同时存在基因分离,可能在两者之间存在互作效应。
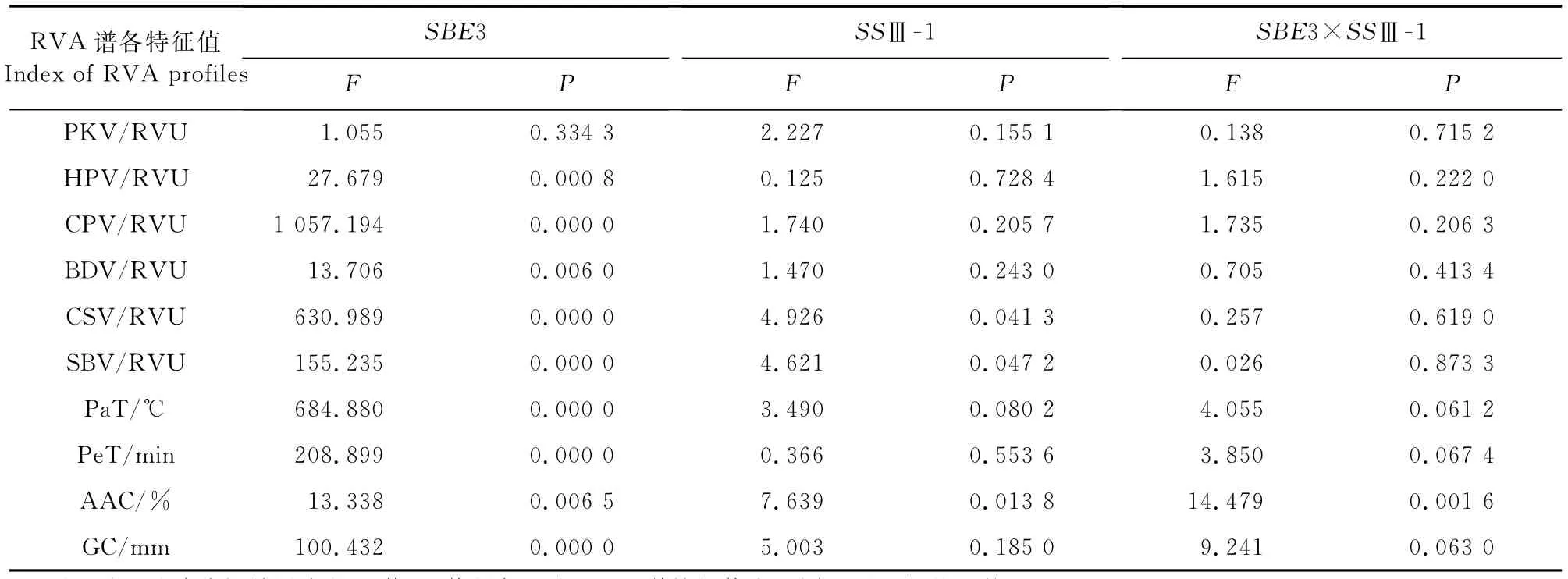
表6 SBE3基因与SSⅢ-1基因的裂区设计分析结果Table 6 The results of split block design for SBE3 gene and SSⅢ-1 gene
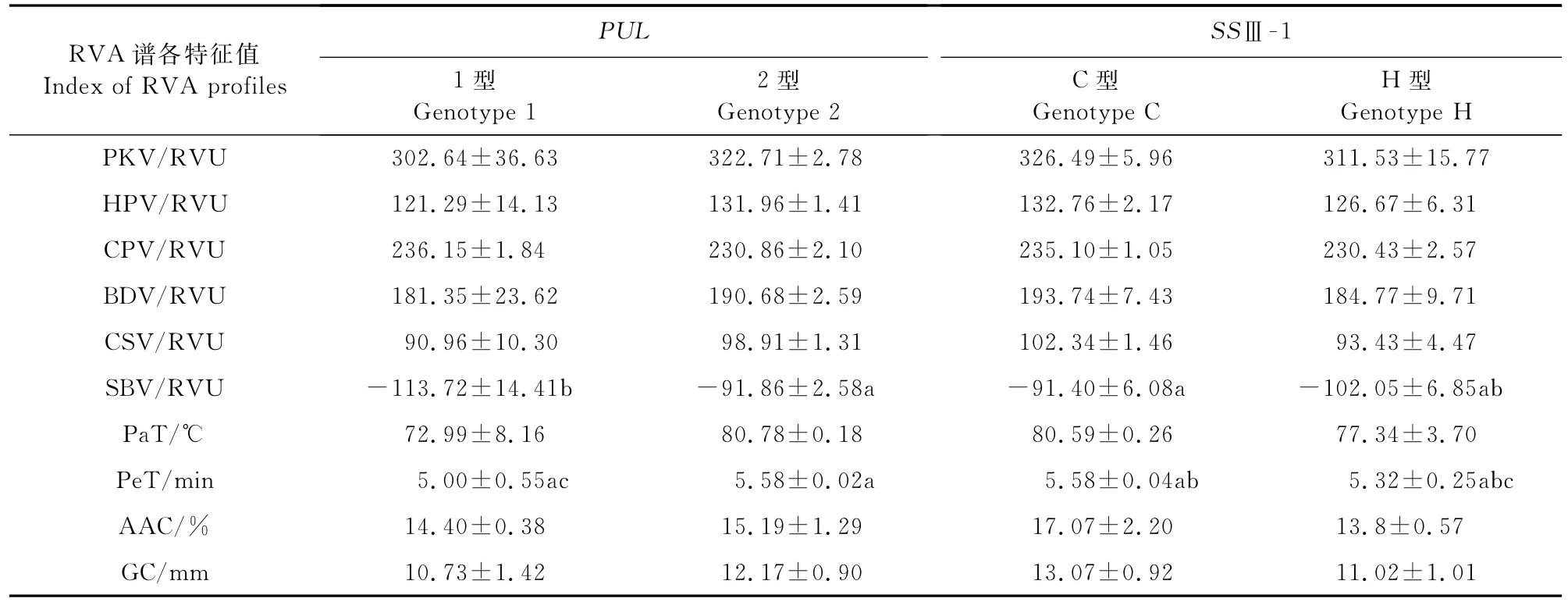
表7 PUL和SSⅢ-1不同基因型稻米品质性状的均值及差异显著性Table 7 The means of quality traits and significance of difference for different PUL and SSⅢ-1 genotypes
(2)PUL和SSⅢ-1基因的互作效应分析 根据株系中SSⅢ-1和PUL基因型组合方式不同,将此株系参试材料分为4类(表8),在不同组合的情况下,Pe T、Pa T和CSV各特征值在各基因型间均存在显著性差异。在相同PUL基因型的遗传背景下,Pe T、Pa T和CSV各指标的C型的均值均大于H型,也即SSⅢ-1基因的CG173R基因型的均值均大于杂合型的均值。
进一步将PUL基因型作为主因素,SSⅢ-1基因作为副因素,进行裂区试验分析,探究PUL基因和SSⅢ-1基因的互作效应,结果见表9。由表9可知,PUL基因对Pe T的效应达到极显著水平,对HPV、CPV、PaT、CSV和AAC的效应达到显著水平。SSⅢ-1基因对CPV、Pe T、Pa T、CSV和GC的效应达到极显著水平,对PKV、HPV和BDV的效应达到显著水平。PUL基因和SSⅢ-1基因的互作效应对PeT、Pa T和CSV的效应达到极显著水平,对PKV、HPV、BDV、CPV、SBV、AAC和GC的效应达到显著水平。
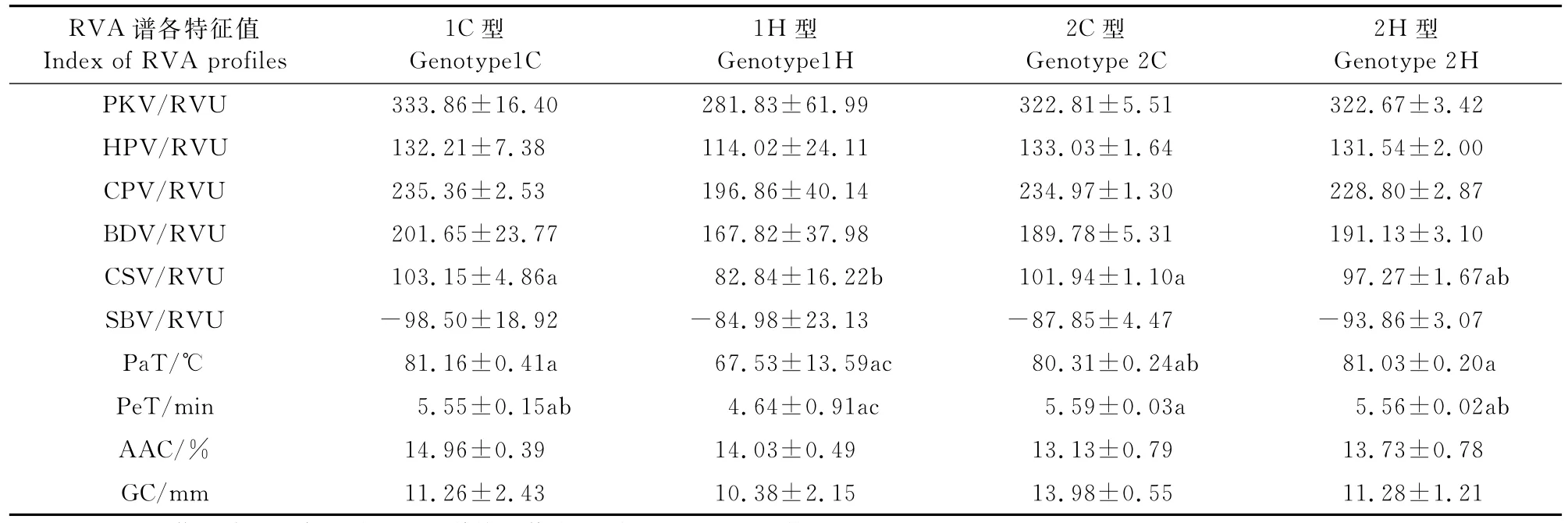
表8 PUL和SSⅢ-1不同基因型组合品质性状的均值及差异显著性Table 8 The means of quality traits and significance of difference for different combinations of PUL and SSⅢ-1 genotypes
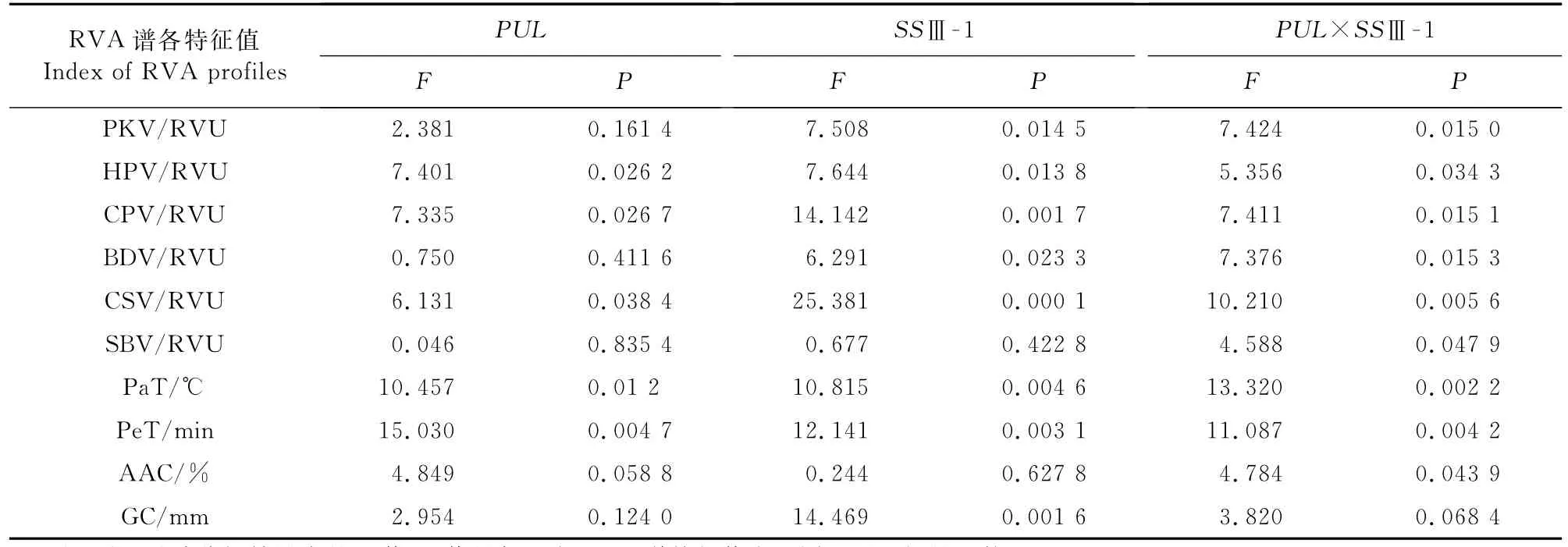
表9 PUL基因与SSⅢ-1基因的列区设计分析结果Table 9 The rsults of split block design for PUL gene and SSⅢ-1 gene
3 讨 论
水稻品质改良是水稻育种的一个重要目标,稻米淀粉合成通路由多个基因共同协作调控完成,解析各基因在调控过程中的功能是水稻品质改良的前提。因此深入研究稻米淀粉合成代谢途径中的一系列基因对稻米理化特性的影响及其遗传规律具有十分重要的意义。
SSⅢ-1基因是可溶性淀粉合成酶基因家族中的一种类型,主要参与分支链的延长。王芳等构建了以‘苏御糯'为供体亲本,‘桂朝2号'为轮回亲本的淀粉合成相关基因的近等基因系,研究表明SSⅢ-1基因对RVA谱特征值HPV、SBV和CSV有显著影响[33]。本研究也发现在S10和S134株系中仅有SSⅢ-1单基因分离的情况下,SSⅢ-1基因对稻米的AAC有极显著性的影响。
SBE3(SBEⅡb)基因主要在种子中特异性表达,主要影响淀粉A链B1链的合成[34]。王芳等研究认为SBE3被‘苏御糯'相应基因置换后使得PKV、HPV和CPV极显著下降,对其他特征值没有显著的影响,但在BC3F2代,SBE1和SBE3基因被‘苏御糯'相应基因置换后,对近等基因系的PKV和HPV没有显著的影响,但使CPV显著下降,SBV、Pe T和Pa T极显著下降。在汪结明等利用RVA干扰技术干涉水稻SBE3基因的表达,发现SBE3活性的下降也导致转基因株系AAC显著提高,同时还发现转基因株系成熟籽粒千粒重显著下降[35]。在S63株系中存在SBE3和SSⅢ-1双基因分离的情况下,本研究发现SBE3基因对HPV和BDV的效应达到极显著水平,SSⅢ-1基因对SBV、Pa T、CSV和AAC的效应达显著性水平;SBE3和SSⅢ-1的互作效应对AAC及RVA谱的部分特征值有显著或极显著性的影响,其中对AAC达极显著水平,对PeT、Pa T和GC的效应达到显著水平。
PUL以极限糊精为底物,仅在胚乳中表达,在淀粉合成中起最后修饰作用,能切去支链淀粉的不恰当分支[36]。房玉伟等研究发现,PUL基因对GC的变异有一定的影响,并且在糯稻背景下,PUL基因对RVA谱的影响最为显著,包括PKV、HPV、CPV、SBV、Pe T和Pa T在内的多个特征值都受到PUL基因影响[37]。He等利用‘南京11'和Balilla为亲本构建加倍单倍体群体,结果认为PUL基因对AAC、GC和GT没有遗传效应;在‘苏御糯'ב扬辐糯4号'衍生的F2群体中PUL基因对AAC和GC等指标的变异没有影响,却发现PUL与SSⅡa之间存在互作,对PaT的变异有显著的影响[38]。康翠芳等[27]研究发现在BC1F9代的305株系中,仅有PUL基因发生分离时,PUL基因对稻米的AAC有显著性的影响。本研究发现,在BC1F10代的S52株系中存在PUL和SSⅢ-1双基因分离的情况下,PUL基因对HPV、CPV、Pa T、CSV和AAC等指标有显著影响,对PeT有极显著影响。SSⅢ-1基因对Pe T、HPV和BDV有显著影响,对CPV、PeT、Pa T、CSV和GC有极显著影响,并且PUL和SSⅢ-1的互作对PKV、HPV、BDV、CPV、SBV、AAC、GC有显著影响,对PeT、Pa T和CSV有极显著影响。
在Wxb和SSⅡ-3基因背景下,利用回交重组自交系通过对参与淀粉合成的SSⅢ-1、SBE3和PUL等微效基因效应的研究,表明SSⅢ-1单基因效应及SBE3和SSⅢ-1的互作效应对水稻的AAC有极显著影响,SBE3和SSⅢ-1的互作效应及PUL和SSⅢ-1的互作效应对水稻的GC有显著影响,PUL和SSⅢ-1的互作效应对水稻的Pa T有极显著影响,这些发现将对改良稻米品质和加快水稻优质育种具有重要意义。
[1] WU H K(吴洪恺),LIANG G H(梁国华),GU Y J(顾燕娟),et al.The effect of the starch-synthesizing genes on RVA profile characteristics in rice(Oryza sativa L.)[J].Acta Agronomica Sinica(作物学报),2006,32(11):1 597—1 603(in Chinese).
[2] 韩月澎.稻米淀粉合成相关基因对品质的影响及分支酶基因Sbe1、Sbe3克隆与分析[D].江苏扬州:扬州大学,2004.
[3] OHDANT,FRANCISCO PB JR,SAWADA T,et al.Expression profiling of genes involved in starch synthes is in sink and source organs of rice[J].Journal of Experimental Botany,2005,56(422):3 229—3 244.
[4] PANDEY M K,RANI N S,MADHAV M S,et al.Different isoforms of starch-synthesizing enzymes controllingamylose and amylopectin content in rice(Oryza sativa L.)[J].Biotechnology Advances,2012,30(6):1 697—1 706.
[5] GAO Z Y,ZENG D L,CUI X,et al.Map-based cloning of the ALK gene,which controls the GT of rice[J].Sciance China(Ser C),2003,46:661—668.
[6] UMEMOTO T,AOKI N.Single-nucleotide polymorphisms in rice starch synthaseIIa that alter starch gelatinization and starch as-sociation of the enzyme[J].Functional Plant Biology,2005,32:763—768.
[7] UMEMOTO T,YANO M,SATOH H,et al.Map-ping of a gene responsible for the difference in amylopectinstructure between japonicatype and indica-type rice varieties[J].Theoretical and Applied Genetics,2002,104:18.
[8] BAO J S,CORKE H,SUN M.Nucleotide diversity in starch synthaseIIa and validation of single nucleotide polymorphisms in relationto starch gelatinization temperature and other physicochemicalproperties in rice(Oryza sativa L.)[J].Theoretical and Applied Genetics,2006,113:171—1183.
[9] NAKAMURA Y,FRANCISCO JR P B,HOSAKA Y,et al.Essential amino acids of starch synthaseIIa differentiate amylopect in structure and starch quality between japonica and indica rice varieties[J].Plant Molecular Biology,2005,58:213—227.
[10] TAN Y F,LI J X,YU S B,et al.The three important traits for cooking and eating quality of rice grains arecontrolled by a single locus inan elite rice hybrid,Shanyou 63[J].Theoretical and Applied Genetics,1999,99:642—648.
[11] BAO J S,ZHENG X W,XIA Y W,et al.QTL mapping for the paste viscosity characteristics inrice(Oryza sativa L.)[J].Theoretical and Applied Genetics,2000,100:280—284.
[12] WANG L Q,LIU W J,XU Y,et al.Genetic basis of 17 traits and viscosity parameters characterizing the eating and cooking quality of rice grain[J].Theoretical and Applied Genetics,2007,115:463—476.
[13] WAN X Y,WAN J M,SU C C,et al.QTL detection for eating quality of cooked rice in a population of chromosome segment substitution lines[J].Theoretical and Applied Genetics,2004,110:71—79.
[14] FAN C C,YU X Q,XING Y Z,et al.Themain effects,epistatic effects and environmental interactions of QTLs on the cooking and eating quality of rice in a doubled-haploid line population[J].Theoretical and Applied Genetics,2005,110:1 445—1 452.
[15] HE Y,HAN Y P,JIANG L,et al.Function alanalysis of starch-synthesis genes in determining rice eating and cooking qualities[J].Molecular Breeding,2006,18:277—290.
[16] YAN CH J(严长杰),FANG Y W(房玉伟),LI M(李 敏),et al.Effect of PUL allelic variation on rice cooking and eating quality[J]. Acta Agronomica Sinica(作物学报),2010,6(5):728—732(in Chinese).
[17] NAKAMURA Y,TAKWICHI T,KAWAGUCHI K.Changes in enzyme activities associated with carbohydrate metabolism during the development of rice endosperm[J].Plant Physiology,1992,84:329—335.
[18] NAKAMURA Y.Towards a better understanding of the metabolic system for amylopection biosynthesis in plants:rice endosperm as a model tissue[J].Plant Cell Physiology,2002,43:718—725.
[19] KRISHNAN H B,REEVES C D,OKITA T W.ADP glucose pyrophosphorylase is encoded by different mRNA transcripts in leaf and endosperm of cereals[J].Plant Physiology,1986,81(2):642—645.
[20] MU FORSTER C,HUANG R,POWERS J R,et al.Physical association of starch biosynthetic enzymes with starch graules of maizy endosperm[J].Plant Physiology,1996,111:821—829.
[21] BOYER C D,PREISS J.Properties of citrate-stimulated starch synthesis catalyzed by starch synthaseI of developing maize kernels[J]. Plant Physiology,1979,64(6):1 039—1 042.
[22] KREIS M.Primer dependent and independent forms of soluble starch synthetase from developing barle endosperms.[J].Planta,1980,148(4):412—416.
[23] NISHI A,NAKAMURA Y,TANAKA N,et al.Biochemical and genetic analysis of the effects of amylose-extender mutation in rice endosperm[J].Plant Physiology,2001,127(2):459—472.
[24] JAMES M G,ROBERTSON D S,MYERS A M.Characterization of themaize gene sugary1,a determinant of starch composition in kernels[J].Plant Cell,1995,7:417—429.
[25] KUBO A,FUJITA N,HARADA K,et al.The starch debraching enzymes isoamylase and pullulanase are both involved in amylopectin biosynthesis in rice endosperm[J].Plant Physiology,1999,121:399—409.
[26] FUJITA N,TOYOSAWA Y,UTSUMI Y,et al.Characterization of pillulanase(PUL)-deficient mutants of rice(Oryza sativa L.)and the function of PUL on starch biosynthesis in the developing rice endosperm[J].Journal of Experimental Botany,2009,60:1 009—1 023.
[27] KANG C F(康翠芳),XIANG X CH(向珣朝),LONG X L(龙小林),et al.Effects of the starch-synthesizing Genes SSⅠ,SSⅢ-1 and PUL on rice(Oryza sativa L.)Quality[J].Journal of Agricultural Biotechnology(农业生物技术学报),2015,23(3):311—319(in Chinese).
[28] TIAN ZH X(田志喜),YAN CH J(严长杰),QIAN Q(钱 前),et al.Development of gene-tagged molecular markers for starch synthesis-related genes in rice[J].Chinese Science Bulletin(科学通报),2010,55(26):2 591—2 260(in Chinese).
[29] CAI X L(蔡秀玲),LIU Q Q(刘巧泉),TANG S Z(汤述翥),et al.Development of a molecular marker for screening the rice cultivars with intermediate amylose content in Osyza sative subsp.indica[J].Journal of Plant Physiology and Molecular Biology(植物生理与分子生物学学报),2002,28(2):137—144(in Chinese).
[30] XIANG X CH(向珣朝),LI J H(李季航),HE L B(何立斌),et al.Marker 2 assisted selection of ZmC4Ppc transformed rice and their performance of yield components[J].Chinese Journal of Rice Science(中国水稻科学),2007,21(1):25—30(in Chinese).
[31] 中华人民共和国国家标准.GB/T15683—2008.大米―直链淀粉含量的测定[S].北京:中国标准测定出版社,2008.
[32] American Association of Cereal Chemists(AACC).Approved methods of the AACC.10thed.Methods 61-02 for RVA.St.Paul,MN[S],USA:AACC,2000.
[33] 王 芳.淀粉合成相关基因近等基因系的构建及其对稻米蒸煮品质影响的初步研究[D].江苏扬州:扬州大学,2007.
[34] 闵绍楷,熊振民.水稻遗传与品质改良[M].杭州:浙江科学技术出版社,1984.
[35] WANG J M(汪结明),ZHANG J(张 建),JIANG H Y(江海洋),et al.Effects of RNA interference of SBE3 gene expression on starch accumulation and key enzymes activities involved in starch synthesis in transgenic rice grain[J].Acta Agronomica Sinica(作物学报),2010,36(2):313—320(in Chinese).
[36] LOYD J R,LANDSCHUTZE V,KOSSMARM J.Simultaneous antisense inhibition of two starch-synthase isoforms in potato tubers leads to accumulation of grossly modified amylopectin[J].The Biochemical Journal,1999,338(2):515—521.
[37] 房玉伟.糯稻背景下淀粉合成相关基因遗传网络初步探析及功能验证[D].江苏扬州:扬州大学,2009.
[38] HE Y,HAN Y P,JIANG L,et al.Functional analysis of starch-synthesis genes in determining rice eating and cooking qualities[J].Molecular Breeding,2006,18(4):277—290.
(编辑:宋亚珍)
Effects of SSⅢ-1,SBE3 and PUL on Eating and Cooking Qualities of Rice under the Background of Backcross Inbred Lines
XU Shunju,XIANG Xunchao*,KANG Cuifang,LONG Xiaolin,SU Wenli,YANG Bowen,WU Jiafu
(Lab of Plant Molecular Genetics and Breeding,Southwest University of Science and Technology,Mianyang,Sichuan 621010,China)
Rice is one of the major cereal crops and eating and cooking qualities are very important for improving commodification of rice and peopleˊs life.We used backcross inbred lines(BILs)of BC1F10generation as materials which were constructed by two parents of indica photo-thermo-sensitive gene male sterile(PTGMS)line Guangzhan 63S and rice potential restorer line CG173R and they contained the same alleles of starch synthaseⅡa gene(SSⅡ-3)and granule bound starch synthase gene(Wxb),and their hybrid by backcrossing and selfing for several generations.Furthermore,the genotypes,the eating and cooking qualities(ECQs)and RVA profiles were measured among the lines of BILs so that genetic effects of the minor geneswere analyzed.The results showed that:1)Five different starch synthesis-related genes ADP-glucose pyrophos-phorylase large subunit ADPG(AGPlar),starch branching enzymeⅢ(SBE3),Pullulanase gene(PUL),Soluble Starch SynthaseⅠgene(SSⅠ)and Soluble Starch SynthaseⅢ-1 gene(SSⅢ-1)existed difference for two parents.2)SSⅢ-1,SBE3 and PUL genes had single gene separation,and two genes separation of SSⅢ-1 and SBE3 genes,SSⅢ-1 and PUL genes in lines of BC1F10.3)Their different effect of genotypes,and interaction had significant influence at 0.05 level or at 0.01 level on apparent amylase content(AAC),the gel consistency(GC)and part indices of RVA profiles.4)Effects of SSⅢ-1 had significant influence on AAC at 0.01 level.Interaction effect of SBE3 and SSⅢ-1 had significant influence on AAC at 0.01 level and had significant influence on Peak time(Pe T),Pasting temperature(Pa T)and GC at 0.05 level.Interaction effect of PUL and SSⅢ-1 genes had significant influence on Pe T,Pa T and consistence value(CSV)at 0.01 level and had significant influence on peak viscosity(PKV),hot paste viscosity(HPV),Breakdown value(BDV),Cool paste viscosity(CPV),Setback value(SBV),AAC and GC at 0.05 level.In the background of Wxband SSⅡ-3 genes,the results demonstrated that effects of SSⅢ-1 and the genes interaction between SBE3 and SSⅢ-1 have significant influence on AAC at 0.01 level;Interaction effects between SBE3 and SSⅢ-1 and between PUL and SSⅢ-1 have significant influence on GC at 0.05 level;Interaction effect between PUL and SSⅢ-1 has significant influence on Pa T at 0.01 level.The studies have importance for improving the rice eating and cooking qualities and accelerating the research on rice quality breeding.
rice(Oryza sativa L.);backcross inbred lines(BILs);soluble starch synthaseⅢ-1 gene(SSⅢ-1);starch branching enzyme 3 gene(SBE3);pullulanase gene(PUL)
Q789
A
1000-4025(2015)10-1978-11
10.7606/j.issn.1000-4025.2015.10.1978
2015-07-10;修改稿收到日期:2015-08-29
四川省教育厅重点项目(13ZA0272);西南科技大学重点科研平台专职科研创新团队建设基金(14tdgc07);四川省生物质资源利用与改性工程技术研究中心开放基金(12ZXSK08)
许顺菊(1989—),女,在读硕士研究生,主要从事分子标记辅助选择改良稻米品质研究。E-mail:505525398@qq.com
*通信作者:向珣朝,博士,教授,硕士生导师,主要从事分子遗传与育种研究。E-mail:xiangxunchao@swust.edu.cn
猜你喜欢
亚热带植物科学(2022年1期)2022-05-17
美食(2022年5期)2022-05-07
早期教育(家庭教育)(2021年11期)2021-12-17
少儿科学周刊·儿童版(2021年21期)2021-12-11
四川农业科技(2019年5期)2019-07-01
中国粮食经济(2018年5期)2018-12-27
浙江柑橘(2016年1期)2016-03-11
现代检验医学杂志(2015年6期)2015-02-06
作物研究(2014年6期)2014-03-01
中国糖料(2013年1期)2013-01-22
