
干旱胁迫对鹿角杜鹃种子萌发和幼苗生理特性的影响
2015-06-27 05:51苏家乐刘晓青何丽斯陈尚平熊才法
西北植物学报 2015年7期
李 畅,苏家乐,刘晓青,何丽斯,陈尚平,肖 政,熊才法
(江苏省农业科学院园艺研究所/江苏省高效园艺作物遗传改良重点实验室,南京210014)
干旱胁迫对鹿角杜鹃种子萌发和幼苗生理特性的影响
李 畅,苏家乐*,刘晓青,何丽斯,陈尚平,肖 政,熊才法
(江苏省农业科学院园艺研究所/江苏省高效园艺作物遗传改良重点实验室,南京210014)
为探明鹿角杜鹃种子萌发和幼苗生长期的耐旱性,以鹿角杜鹃干种子和90d苗龄幼苗为材料,采用聚乙二醇(PEG-6000)模拟不同程度的干旱胁迫,研究干旱胁迫对其种子萌发、早期幼苗生长及幼苗的细胞膜透性、MDA含量、有机渗透调节物质和抗氧化酶活性的影响,并对种子萌发率、早期幼苗生长量与PEG胁迫浓度间进行了回归分析。结果表明:(1)5%~25%PEG胁迫范围内,随着干旱胁迫程度的增加,鹿角杜鹃种子的发芽启动时间推迟,发芽持续时间延长,发芽率、发芽势、发芽指数、活力指数和幼苗生长量显著降低;重度干旱胁迫(25%PEG)下,鹿角杜鹃种子完全未萌发。(2)发芽率、发芽势、发芽指数、活力指数以及幼苗生长量的变化均与干旱胁迫程度呈极显著负相关关系,回归分析求得鹿角杜鹃种子萌发的半致死PEG干旱胁迫浓度为15.68%、半致矮PEG干旱胁迫浓度为15.37%。(3)随着PEG胁迫浓度的增加,鹿角杜鹃幼苗叶片SOD活性呈先升后降的趋势,但各胁迫处理仍显著高于CK(0%PEG);细胞膜透性、MDA、脯氨酸、可溶性糖含量、POD和CAT活性则在中度(15%~20% PEG)和重度胁迫下显著升高,与干旱胁迫程度呈极显著正相关关系。研究表明,干旱胁迫显著抑制了鹿角杜鹃种子萌发和早期幼苗生长,使其细胞膜受到损伤,同时鹿角杜鹃可通过体内渗透调节物质和抗氧化酶活性的增加来适应干旱环境,使得自身受抑制、损伤程度降到最低。
鹿角杜鹃;干旱胁迫;种子萌发;早期幼苗生长;幼苗生理
鹿角杜鹃(Rhododendron latoucheae)是杜鹃花科(Ericaceae)杜鹃花属马银花亚属(subgenus Azaleastrum)常绿灌木或小乔木,作为杜鹃花属中垂直分布最宽的种类之一,其主要分布于长江中下游及其以南地区海拔160~2 670m的山坡林缘或杂木林内,生境阴凉湿润[1-2]。鹿角杜鹃叶片狭长浓绿、花色清雅有清香,花叶兼赏,生长势强,研究开发前景广阔[1-3]。但其在引种繁育过程中会受到诸多环境因素的影响,水分是极为重要的影响因子之一,尤其是干旱环境。实生繁殖是目前鹿角杜鹃主要繁殖方式,且科研工作者们从杜鹃花种子繁殖的实践认识到,从种子萌发获得的幼苗开始驯化,使之适应当地的气候,是一种有效的引种驯化手段[4-5]。而种子萌发成苗又是植物生活史中抗逆性最为脆弱的发育阶段[6],故开展种子萌发期抗旱特性、对干旱环境的适应性及响应机制研究尤为重要。
目前,关于鹿角杜鹃的报道主要集中在园林应用[3]、繁殖技术[7-8]、光合生理[9]、耐盐评价[10]、辐射育种[11]等方面,对其耐旱特性仅限于“喜湿润”的笼统描述,未见关于鹿角杜鹃种子耐旱方面的研究报道。聚乙二醇(polyethylene glycol,PEG)是一种渗透调节剂,近些年来作为水分胁迫剂被广泛应用于植物种子萌发期的耐旱性研究[6,12-17]。因此,本试验采用不同浓度PEG-6000溶液模拟不同程度的干旱胁迫状况,研究干旱胁迫对鹿角杜鹃种子萌发、早期幼苗生长、细胞膜透性、渗透调节能力、抗氧化系统等的影响,以期了解鹿角杜鹃对干旱胁迫的适应性及响应机制,为鹿角杜鹃的合理开发、引种栽培及耐旱育种提供理论依据。
1 材料和方法
1.1 材 料
供试的鹿角杜鹃种子采集于浙江天目山自然保护区野生植株,由浙江农林大学鉴定提供,自然晾干,除去干瘪空粒和杂质,室温干燥器内保存。鹿角杜鹃种子净度97%,种子千粒重(0.1003±0.0084)g,TTC法[18]测定种子活力为87%。试验用鹿角杜鹃幼苗由种子萌发得到。
1.2 方 法
1.2.1 聚乙二醇模拟干旱胁迫浓度的设置 采用PEG-6000模拟不同程度的干旱胁迫。根据预备试验,共设6个胁迫浓度,其PEG-6000浓度(M/V)分别为0%(去离子水,CK)、5%、10%、15%、20%和25%,与之相对应的溶液水势分别约为0、-0.05、-0.20、-0.40、-0.60、-0.86MPa[19]。其中,-0.05和-0.20MPa水势为轻度干旱胁迫,-0.40和-0.60MPa水势为中度干旱胁迫,-0.86MPa水势为重度干旱胁迫[13]。
1.2.2 种子萌发胁迫试验 体式显微镜Olympus SZX10下挑取籽粒饱满、大小均匀的鹿角杜鹃种子,经0.3%(M/V)KMnO4溶液浸种消毒15min,去离子水充分洗净,吸水纸吸干种子表面水分后,将种子均匀排布于培养皿内浸透了不同浓度PEG-6000溶液的滤纸上,置于光照培养箱中进行萌发测定。培养条件为:温度25℃,光照强度120μmol· m-2·s-1,光照时间12h·d-1。每天向滤纸加PEG-6000溶液数滴,以浸透滤纸并稍有剩余为度,每4d更换1次滤纸,以减少水势变动。每处理100粒,重复3次。以胚根长度达种子长度的1/2为发芽标准,连续5d无萌发种子视为萌发结束,每24h观察记录种子发芽数;计算发芽启动时间、发芽持续时间、发芽率、发芽势、发芽指数,并结合幼苗生长量测量结果计算活力指数;种子发芽后第2~10天测量幼苗生长量(生长点至根尖总长),每2d测量1次,每处理测量10株,重复3次。
发芽启动时间:从发芽试验开始到第1粒种子开始萌发所需时间(天数)
发芽持续时间:种子开始萌发到最后一个萌发的总天数
发芽率(GR)=种子发芽总数/供试种子总数(100粒)×100%
发芽势(GE)=日发芽种子数达到高峰时,发芽种子总数/供试种子总数(100粒)×100%
发芽指数(GI)=∑(在不同时间的发芽数/相应的发芽天数)
活力指数(VI)=发芽指数×第10天幼苗高度(mm)
1.2.3 幼苗生长胁迫试验及生理指标测定 试验在智能玻璃温室中进行,挑选生长一致的90d苗龄的鹿角杜鹃幼苗移植于72孔穴盘中,每孔1株,基质为泥炭和珍珠岩等比例混合而成。缓苗7d后进行干旱胁迫试验,胁迫梯度设置6个水平(见1.2.1),每处理水平72株(1穴盘),3次重复。胁迫7d后,取幼苗叶片进行生理指标的测定。其中,丙二醛(MDA)含量的测定采用硫代巴比妥酸法[20],细胞膜相对透性采用电导仪法[20],游离脯氨酸含量测定采用酸性茚三酮法[20],可溶性糖测定采用蒽酮比色法[20]。超氧化物歧化酶(SOD)活性、过氧化氢酶(CAT)、过氧化物酶(POD)活性采用试剂盒法,试剂盒由南京建成生物有限公司提供[21]。所需上清液均按李合生[20]的方法制备,按试剂盒说明书操作、比色,运用试剂盒提供的公式计算出SOD、POD、CAT活性。
1.3 数据处理
所有测定结果取平均值±标准差,采用SPSS 18.0软件进行单因素方差分析和相关性分析,使用最小显著差数法(LSD)比较差异显著性,Excel 2003下绘制图表并拟合回归方程。
2 结果与分析
2.1 干旱胁迫对鹿角杜鹃种子萌发的影响
2.1.1 发芽进程 如图1所示,鹿角杜鹃干种子正常条件下(0,CK)发芽启动时间为7d,发芽持续时间为8d。PEG胁迫浓度≤10%时,鹿角杜鹃发芽持续时间较CK延长1d,发芽启动时间不变,但第7天的发芽率较CK显著降低;当PEG浓度≥15%时,鹿角杜鹃种子发芽启动时间较CK延后,且随着PEG胁迫浓度的增加,鹿角杜鹃种子发芽启动时间延后时间越长,如20%PEG胁迫下的鹿角杜鹃种子,其发芽启动时间(10d)延迟3d,发芽持续时间(11d)延长3d;25%PEG胁迫下的鹿角杜鹃种子未见萌发。说明轻度干旱胁迫(5%~10%PEG)对鹿角杜鹃种子萌发已具有延缓作用,且随着干旱胁迫程度的加强(PEG浓度的增加),种子吸收水分越发困难,发芽启动时间延后,发芽持续时间延长。
2.1.2 萌发指标 发芽率、发芽势、发芽指数和活力指数是反映种子发芽能力的主要指标。由表1可以看出,随PEG胁迫浓度的增加,鹿角杜鹃种子发芽率、发芽势、发芽指数和活力指数均呈现明显降低趋势,且各测试萌发指标均与CK差异显著,其中的活力指数在各处理间差异均达显著水平(P<0.05)。20%PEG中度干旱胁迫下,发芽率、发芽势、发芽指数和活力指数4个萌发指标仅为CK的13.72%、12.73%、10.55%和2.94%,25%PEG重度干旱胁迫则完全抑制了鹿角杜鹃种子萌发。以上结果说明干旱胁迫对鹿角杜鹃种子萌发抑制作用明显,整体表现为浓度依赖规律;同时也可以看到,在15%PEG中度干旱胁迫下,鹿角杜鹃种子的发芽率、发芽势、发芽指数和活力指数仍能达到CK的58.41%、46.36%、51.10%和24.66%,又说明鹿角杜鹃具有一定的抗旱性。
另外,进一步分析发现,鹿角杜鹃种子发芽率、发芽势、发芽指数和活力指数均与PEG胁迫浓度呈极显著负相关;建立发芽率与PEG胁迫浓度的回归方程[y=-769.05x2-122.21x+74.845(R2=0.967 5)],推算得出鹿角杜鹃种子的半致死PEG模拟干旱胁迫浓度为15.68%。

图1 干旱胁迫下鹿角杜鹃种子发芽进程Fig.1 The seed germinating progress of R.latoucheae under drought stress
2.2 干旱胁迫对鹿角杜鹃早期幼苗生长的影响
早期幼苗生长对干旱胁迫较为敏感,在5%~20%PEG浓度范围内,随胁迫时间的延长,不同PEG浓度下的鹿角杜鹃早期幼苗生长量均逐渐增加,但生长量增加的幅度则随PEG胁迫浓度增加而呈现降低趋势(图2)。其中,20%PEG胁迫下鹿角杜鹃第10天幼苗生长量较第2天幼苗生长量增加了1.11倍,而CK第10天幼苗生长量则较第2天幼苗生长量增加了4.63倍,二者差异显著。种子发芽4d后,5%~25%PEG浓度胁迫下的鹿角杜鹃早期幼苗生长量均显著低于CK,且随着胁迫时间的延长,处理间差异增大,种子发芽第6天后处理间差异均达到显著水平;种子发芽第10天,轻度干旱(5%和10%)胁迫下幼苗生长量分别比CK降低8.33%和26.85%,而中度干旱胁迫(15%和20%)下则比CK分别降低51.65%和72.02%。进一步相关性分析表明,幼苗生长量与PEG胁迫浓度呈极显著负相关。说明5%~20%PEG胁迫浓度范围内,干旱胁迫可抑制鹿角杜鹃早期幼苗生长,且抑制作用随胁迫时长的增加及胁迫浓度的增加而增大。建立幼苗生长量(第10天)与PEG胁迫浓度的回归方程[y=-103.09x2-30.115x+13.895(R2=0.996 8)],推算得出鹿角杜鹃种子的半致矮PEG模拟干旱胁迫浓度为15.37%。
2.3 干旱胁迫对鹿角杜鹃幼苗细胞膜相对透性和MDA含量的影响
逆境条件下,植物叶片的细胞膜相对透性通常用叶片的相对电导率表示。如图3所示,随PEG胁迫浓度的增加,鹿角杜鹃幼苗叶片的相对电导率呈现逐渐上升的趋势;且在5%~25%PEG胁迫浓度范围内,相对电导率均与CK呈显著性差异;25% PEG重度干旱胁迫下,相对电导率升至CK的2.93倍。相关性分析表明,相对电导率和PEG胁迫浓度之间的相关系数为0.969,并达到极显著水平。说明不同程度PEG胁迫对鹿角杜鹃幼苗细胞膜透性均产生伤害。

表1 干旱胁迫下鹿角杜鹃种子萌发指标的变化Table 1 The seed germination of R.latoucheae under drought stress
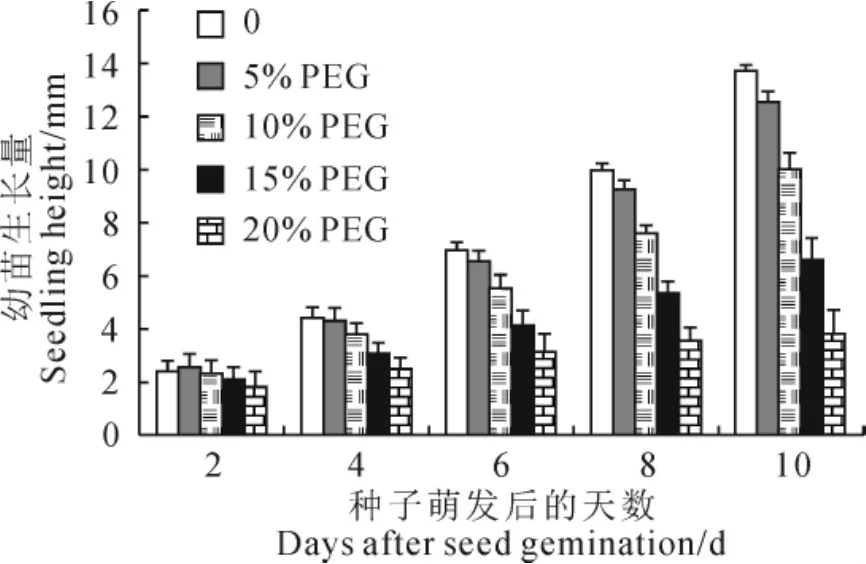
图2 干旱胁迫下鹿角杜鹃幼苗生长的变化不同小写字母表示不同PEG浓度处理间在0.05水平差异显著;下同Fig.2 The seedling growth of R.latoucheae under drought stress The different normal letters indicate the significant difference among the different PEG treatments at 0.05level;The same as below
MDA是膜脂过氧化的主要产物之一,因其在细胞中的积累在一定程度上反映了植物体内自由基的动态和细胞的受损程度,在衡量植物对逆境条件反应的强弱中广泛应用[22]。与相对电导率变化趋势相同,5%~25%PEG浓度范围内,鹿角杜鹃幼苗MDA含量亦随PEG胁迫浓度的增加呈现增加趋势;PEG胁迫浓度≥15%时,MDA含量与CK差异显著(图3)。相关性分析表明MDA含量与PEG胁迫浓度之间的相关系数为0.949,呈极显著正相关。可见,随干旱胁迫程度的增强,鹿角杜鹃幼苗细胞膜系统已受到不同程度的损伤。
2.4 干旱胁迫对鹿角杜鹃幼苗有机渗透调节物质含量的影响
游离脯氨酸和可溶性糖作为植物体内2种有机渗透调节物质,在干旱胁迫中起重要作用[23]。如图4所示,鹿角杜鹃幼苗游离脯氨酸和可溶性糖含量对干旱胁迫的响应特征一致,均随PEG胁迫浓度的增加而持续上升;PEG胁迫浓度≥15%时,2指标含量与相应CK差异均达到显著水平。其中,游离脯氨酸含量在中度干旱胁迫(15%PEG)下较对照增加了39.57%,在重度干旱胁迫(25%PEG)下则显著增加到CK(139.81μg·g-1)的2.13倍;可溶性糖含量随PEG胁迫浓度的增加而产生的积累量的增幅则较游离脯氨酸小,在15%PEG中度干旱胁迫下较CK增加了28.00%,25%PEG重度干旱胁迫下达到最大值,为CK(1.52μg·g-1)的1.47倍。所测的游离脯氨酸含量和可溶性糖含量均与PEG胁迫浓度呈正相关关系,且均达到极显著水平。可见,PEG胁迫下鹿角杜鹃幼苗可通过增加自身有机渗透胁迫调节物质的含量来抵御干旱胁迫。
2.5 干旱胁迫对鹿角杜鹃幼苗抗氧化酶活性影响
SOD、POD和CAT是植物体内3种重要的抗氧化酶,逆境条件下可在一定程度内降低活性氧对植物的伤害。鹿角杜鹃幼苗3种抗氧化酶对PEG干旱胁迫的响应不尽相同(图5)。其中,在5%~25%PEG胁迫浓度范围内,SOD活性均显著高于CK,且随PEG胁迫浓度的升高呈先升高后降低的趋势,并于15%PEG胁迫下升至最高,为CK的1.42倍;POD和CAT活性则随PEG胁迫浓度的升高而呈持续上升趋势,其中POD活性在PEG胁迫浓度≥10%较CK显著升高,CAT活性则在PEG胁迫浓度≥15%时与CK差异显著,POD和CAT活性均在25%PEG胁迫浓度下升至最高,分别为CK的2.24倍和1.62倍。3种抗氧化酶活性均与PEG胁迫浓度呈正相关关系,其中POD和CAT活性与PEG胁迫浓度相关性达到极显著水平。说明干旱胁迫下鹿角杜鹃幼苗可通过抗氧化酶活性增强来抵御过量的活性氧自由基造成的损伤。
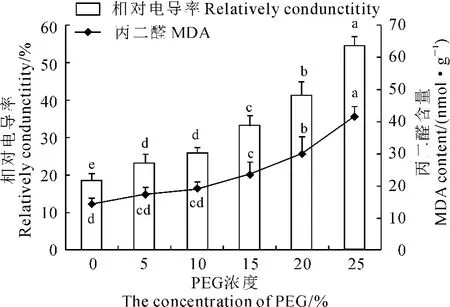
图3 干旱胁迫下鹿角杜鹃幼苗相对电导率和丙二醛含量Fig.3 The relatively conductivity and MDA content of R.latoucheae seedlings under drought stress

图4 干旱胁迫下鹿角杜鹃幼苗脯氨酸和可溶性糖含量Fig.4 The proline content and soluble sugar content of R.latoucheae seedlings under drought stress
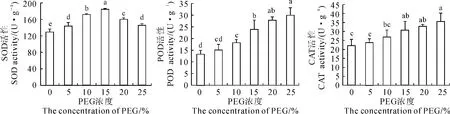
图5 干旱胁迫下鹿角杜鹃幼苗抗氧化酶活性Fig.5 The antioxidant enzyme activities of R.latoucheae seedlings under drought stress
3 讨 论
3.1 鹿角杜鹃种子萌发及早期幼苗生长对PEG模拟干旱胁迫的响应特征
植物种子的萌发指标直接反映种子耐逆境能力[16]。在本研究中,从5%PEG胁迫浓度开始,各浓度PEG干旱胁迫处理对鹿角杜鹃种子的萌发及早期幼苗的生长就具有明显的延缓和抑制作用,表现为随PEG胁迫浓度的增加,种子的发芽率、发芽势、发芽指数和活力指数显著降低,发芽启动时间推迟,发芽持续时间延长,幼苗的生长量和生长速度下降,这与前人报道的干旱胁迫下五爪金龙(Ipomoea cairica)[12]、麻楝(Chukrasia tabularis)[14]、凤仙花(Impatiens balsamina)[16]、宿根天人菊(Gaillardia aristata)[17]等的种子萌发和幼苗生长规律一致。但与李鹏鹏等[6]对杠柳(Periploca sepium)、李志萍等[15]对栓皮栎(Quercus variabilis)种子研究报道的低浓度PEG对种子萌发有一定的引发作用有所不同,这可能与植物本身的特性及其环境的异质性有关。本试验结果表明轻度(5%~10%PEG)和中度(15%~20%PEG)干旱胁迫下,鹿角杜鹃种子萌发和幼苗生长受到抑制,但对干旱胁迫仍具有一定的忍耐性;在25%PEG重度干旱胁迫下,种子萌发完全受抑制;发芽率、发芽势、发芽指数、活力指数以及幼苗生长量的变化均与干旱胁迫程度呈极显著负相关,回归分析求得鹿角杜鹃种子萌发的半致死PEG干旱胁迫浓度为15.68%、半致矮PEG干旱胁迫浓度为15.37%,即鹿角杜鹃种子萌发的耐旱临界值[15]介于15.37%~15.68%之间。试验结果为人工直播栽培鹿角杜鹃时土壤水分管理提供指导,同时也为鹿角杜鹃耐旱育种提供了适宜的PEG胁迫浓度。
3.2 鹿角杜鹃幼苗对PEG模拟干旱胁迫的生理生化响应特性
干旱胁迫下,植物体内活性氧水平升高,引发膜脂过氧化,使得其产物MDA含量增加,最终导致膜结构及生理完整性的破坏,而植物体在长期进化过程中形成了包括SOD、CAT、POD在内的抗氧化酶防御系统,可清除活性氧,维持自由基生成与消除的动态平衡[21]。干旱胁迫下,本研究中鹿角杜鹃幼苗细胞膜透性及MDA含量的变化与五爪金龙[12]、文冠果(Xanthoceras sorbifolia)[24]等基本一致,均表现为持续升高,且在≥15%PEG胁迫下,二指标与CK差异显著,说明中度和重度干旱胁迫下鹿角杜鹃幼苗的细胞膜受到明显损伤。同时,本试验表明鹿角杜鹃幼苗SOD、CAT、POD对干旱胁迫的响应不尽相同。随PEG胁迫浓度的升高,SOD活性表现出先升后降趋势:轻度干旱胁迫下SOD活性就显著高以清除活性氧;20%~25%PEG胁迫下,SOD活性虽然下降但仍显著高于CK,即中度和重度干旱胁迫下鹿角杜鹃幼苗细胞膜虽然受到一定损伤但未超过SOD活性的忍耐限度,SOD仍具有较高的活性来清除氧自由基造成的伤害。CAT和POD活性则在5%~25%PEG浓度范围内随胁迫程度的加深持续上升,中度和重度干旱胁迫下显著增高。说明鹿角杜鹃幼苗在干旱胁迫下具有较好的清除活性氧的能力,轻度干旱胁迫下主要靠SOD活性的提高来清除自由基,而在中度和高度干旱胁迫下则主要靠3种抗氧化酶协同作用,共同抵御逆境伤害。
植物在遭受干旱胁迫后会使叶片细胞膜透性和MDA含量增加,同时植物体会通过积累一些可溶性糖、脯氨酸等可溶性物质来进行渗透调节[13,15]。本研究中,鹿角杜鹃幼苗可溶性糖和脯氨酸含量均随干旱胁迫程度的加深而增加,这与周玲等[24]对文冠果幼苗、柯世省等[22]对云锦杜鹃(Rhododendron fortunei)的研究结果一致,说明干旱胁迫下鹿角杜鹃幼苗自身可以积累较多的有机渗透调节物质,来调节植物细胞内渗透压与外界平衡,维持渗透平衡和体内水分,以减轻胁迫所造成的伤害。此外,试验中发现可溶性糖和脯氨酸含量的上升晚于细胞膜透性的升高,推测2种渗透调节物质是受细胞膜透性这个指标的诱导而产生的,具体相关机制尚有待进一步探究。
综上所述,干旱胁迫对鹿角杜鹃种子萌发和幼苗生长均具有抑制作用,但同时鹿角杜鹃也通过一系列生物学、生理学的改变来主动适应干旱逆境,使得干旱胁迫对鹿角杜鹃的抑制损伤降到最低。在鹿角杜鹃引种栽培过程中,应充分考虑水分因素,防止干旱损伤。当然,植物在不同生长阶段抗旱性不尽相同,鹿角杜鹃在其生活史其它阶段抗旱性如何,尚待进一步研究。
参考文献:
[1] 中国科学院中国植物志编委会.中国植物志(第57卷第2分册)[M].北京:科学出版社,1994:358.
[2] GAO L M(高连明),LI D ZH(李德铢).Five new synonyms in the genus Rhododendronsubgen.Azaleastrum(Ericaceae)from China[J].Acta Phytotaxonomica Sinica(植物分类学报),2006,44(5):604-607(in Chinese).
[3] HUANG L Q(黄丽群),XIE L SH(谢禄山),YAN L H(颜立红),et al.Initial research on the development of landscape value of wild Rhododendron latoucheae resources in Hunan[J].Journal of Hunan Environment Biological Polytechnic(湖南环境生物职业技术学院学报),2008,14(4):8-10(in Chinese).
[4] ZHANG CH Q(张长芹),FENG B J(冯宝钧),ZHAO G Y(赵革英),et al.Seed propagation of Rhododendron[J].Acta Botanica Yunnanica(云南植物研究),1992,14(1):87-91(in Chinese).
[5] ZHANG Y H(张乐华),LIU X P(刘向平),WANG K H(王凯红),et al.Factors influencing seed germination and seedling survival of Rhododendron Subgenus Hymenanthes[J].Journal of Wuhan Botanical Research(武汉植物研究),2007,25(2):178-164(in Chinese).
[6] LI P P(李鹏鹏),ZHANG W H(张文辉).Seed germination of Periploca sepiumender drought stress and different storage time[J].Bulletin of Botanical Research(植物研究),2012,32(5):567-572(in Chinese).
[7] ZHANG Y H(张乐华),WANG SH SH(王书胜),SHEN W(单 文),et al.Influences of growth media,and hormone types and concentrations on cutting propagation of Rhododendron latoucheae[J].Scientia Silvae Sinicae(林业科学),2014,50(3):45-54(in Chinese).
[8] WANG SH SH(王书胜),LI X H(李晓花),ZHANG Y H(张乐华),et al.Effects of hormone types and concentrations on cutting propagation of Rhododendron latoucheae and its evaluation[J].Guihaia(广西植物),2014,34(2):227-234(in Chinese).
[9] LIAO J Y(廖菊阳),YAN W D(闫文德),WANG G J(王光军),et al.Diurnal changes in photosynthesis of Rhododendron latoucheae[J].Journal of Central South University of Forestry &Technology(中南林业科技大学学报),2011,31(5):117-120(in Chinese).
[10] LI CH(李 畅),SU J L(苏家乐),LIU X Q(刘晓青),et al.Effect of NaCl stress on seed germination of Rhododendron latoucheae[J].Jiangsu Agricultural Sciences(江苏农业科学),2012,40(12):190-192(in Chinese).
[11] LIU X Q(刘晓青),LI CH(李 畅),SU J L(苏家乐),et al.Effect of60Coγ-ray irradiation on Rhododendron latoucheae seed vigor and seedling growth[J].Jiangsu Journal of Agricultural Sciences(江苏农业学报),2014,30(2):458-460(in Chinese).
[12] ZHU H(朱 慧),MA R J(马瑞君),WU SH T(吴双桃),et al.Seed germination and seedling growth of invasive weed Ipomoea cairica under drought stress[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2009,29(2):344-349(in Chinese).
[13] SHI W(史 微),XU H L(徐海量),ZHAO X F(赵新风),et al.Physiological and biochemical responses to drought stress during seed germination of Glycyrrhiza inflata[J].Acta Ecologica Sinica(生态学报),2010,30(8):2 112-2 117(in Chinese).
[14] WU CH(武 冲),ZHONG C L(仲崇禄),MU ZH Q(牟振强),et al.Effect of water stress simulated by PEG on germination ability of Chukrasia tabularis seeds from different provenances[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2012,32(4):774-780(in Chinese).
[15] LI ZH P(李志萍),ZHANG W H(张文辉),CUI Y CH(崔豫川),et al.Effect of PEG simulated drought stress on seed germination and growth physiology of Quercus variabilis[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2013,33(10):2 043-2 049(in Chinese).
[16] ZHANG F Y(张凤银),LI F L(李方良),TAN Y(谭 英).Effects of PEG Imposed drought stress on the seed germination and seeding growth of Impatiens balsamina[J].Journal of South China Normal University(Nat.Sci.Edi.)(华南师范大学学报·自然科学版),2014,46(2):114-118(in Chinese).
[17] ZHANG F Y(张凤银),CHEN CH Y(陈禅友),HU ZH H(胡志辉).Effect of PEG-6000stress on seed germination and seedling physiological characteristics of Gaillardia aristata Pursh.[J].Journal of Northwest Agriculture and Forestry University(Nat.Sci.Edi.)(西北农林科技大学学报·自然科学版),2014,42(1):132-136(in Chinese).
[18] 张志良.植物生理学实验指导[M].北京:高等教育出版社,2003:226-227.
[19] MICHEL B E,KAUFMANN M R.The osmotic potential of polyethylene glycol 6000[J].Plant Physiology,1973,51(5):914-916.
[20] 李合生.植物生理生化试验原理和技术[M].北京:高等教育出版社,2006.
[21] BAI P(白 鹏),RAN CH Y(冉春艳),XIE X Y(谢小玉).Influence of drought stress on physiological characteristics and agronomic traits at bud stage of rapeseed(Brassica napus L.)[J].Scientia Agricultura Sinica(中国农业科学),2014,47(18):3 566-3 576(in Chinese).
[22] KE SH SH(柯世省),YANG M W(杨敏文).Effects of water stress on antixxidant system and lipid peroxidation in leaves of Rhododendron fortunei[J].Acta Horticulturae Sinica(园艺学报),2007,34(5):1 217-1 222(in Chinese).
[23] ASHRAF M,IRAM A.Drought stress induced changes in some organic substances in nodules and other plant parts of two potential legumes differing in salt tolerance[J].Flora,2005,200(6):535-546.
[24] ZHOU L(周 玲),WANG N J(王乃江),ZHANG L N(张丽楠).Effect of PEG treatment on seed germination and growth of seedlings of Xanthoceras sorbifolia[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2012,32(11):2 293-2 298(in Chinese).
(编辑:裴阿卫)
Effects of Drought Stress on Seed Germination and Seedling Physiological Characteristics of Rhododendron latoucheae
LI Chang,SU Jiale*,LIU Xiaoqing,HE Lisi,CHEN Shangping,XIAO Zheng,XIONG Caifa
(Institute of Horticulture Jiangsu Academy of Agricultural Sciences/Jiangsu Key Laboratory for Horticultural Crop Genetic Improvement,Nangjing 210014,China)
In order to explore the drought tolerance of Rhododendron latoucheae in seed germination and seedling growth stage,we studied the effects of drought stress on seed germination,growth,cell membrane permeability,malondialdehyde(MDA)content,the organic osmoregulation substances and antioxidant enzyme activities by choosing dry seeds and seedlings with 90dage and using different concentrations of polyethylene glycol(PEG-6000)simulated drought stress.The regression analyses were done between seed germination,early seedling growth and PEG stress.The results showed that:(1)the initial germination time of R.latoucheae seeds was delayed with the increasing of drought stress degree in 5%-25%PEG range,and the germination duration was prolonged.The seed germination rate,germination potential,germination index,vigor index and early seedling growth were all significantly reduced.The seed could notgerminate absolutely at severe drought stress(25%PEG).(2)The changes of seed germination rate,germination potential,germination index,vigor index and early seedling growth were showed very significant negative correlation with the degree of drought stress.The regression equation between seed germination,early seedling growth and PEG stress showed that the half lethal PEG drought stress concentration was 15.68%,while the semi dwarfing PEG drought stress concentration was 15.37%.(3)With the increase of PEG concentration,SOD activity of R.latoucheae seedling leaves increased firstly and then decreased,but were significantly higher in all the stress treatments than that of CK(0%PEG).Cell membrane permeability,MDA content,proline content,soluble sugar content,POD and CAT activities increased significantly in moderate(15%-20%PEG)and severe drought stress,and the changes of these indexes were showed very significantly positive correlation with the degree of drought stress.The research found that seed germination and seedling growth of R.latoucheae were significantly inhibited by drought stress,and cell membrane was also damaged in different degrees.Thus it could be seen that R.latoucheaeincreased contents of the osmoregulation substances and antioxidant enzyme activities in vivo to adapt to drought stress environment at the same time,making the inhibition and damage degree to the minimum.
Rhododendron latoucheae;drought stress;seed germination;early seedling growth;seedling physiology
Q945.78
A
10.7606/j.issn.1000-4025.2015.07.1421
1000-4025(2015)07-1421-07
2015-03-12;修改稿收到日期:2015-04-20
国家科技支撑计划(2013BAD01B070403);江苏省自然科学基金(BK2012789);江苏省农业科技自主创新项目[CX(12)2016]
李 畅(1982-),女,硕士,助理研究员,主要从事花卉种质资源与遗传育种研究。E-mail:changli529@foxmail.com
*通信作者:苏家乐,研究员,主要从事花卉育种及栽培研究。E-mail:sujl66@aliyun.com
猜你喜欢
江西农业(2022年8期)2022-05-13
绿色科技(2021年21期)2021-11-26
心声歌刊(2021年3期)2021-08-05
内蒙古林业调查设计(2021年3期)2021-07-06
好孩子画报(2020年4期)2020-05-14
小聪仔(幼儿版)(2020年12期)2020-02-01
阅读(快乐英语中年级)(2019年2期)2019-09-10
民族音乐(2018年5期)2018-11-17
乡村地理(2018年3期)2018-11-06
现代园艺(2018年1期)2018-03-15
