
2010年-2014年猪流行性腹泻病毒S 基因遗传进化分析
2015-06-18 11:28刘艳成刘丹丹张文超杜雅楠
动物医学进展 2015年8期
刘艳成,芦 婷,刘丹丹,李 静,张文超,邓 侨,张 伟,杜雅楠
(内蒙古农业大学兽医学院,农业部动物疾病临床诊疗技术重点实验室,内蒙古呼和浩特 010018)
猪流行性腹泻(Porcine epidemic diarrhea,PED)是由猪流行性腹泻病毒(Porcine epidemic diarrhea virus,PEDV)引起的猪以呕吐、脱水、腹泻为主要症状的高度接触性肠道传染病[1]。各生长阶段的猪群均可发病,但以未断乳仔猪最为严重,病死率可高达100%。该病最早于1976年在比利时和英国的部分猪场被发现,此后,荷兰、德国、法国、匈牙利、前苏联、日本、韩国、越南、缅甸、中国台湾等国家和地区也陆续发生本病的流行[2]。我国内地自20世纪80年代初开始陆续有本病发生的报道。2010年以来,随着新毒株的出现,该病在我国的流行情况更趋严重[3]。美国自2013年5月PED首次暴发以来,已迅速蔓延至30多个州,波及4 000多个养殖场,累计死亡仔猪超过700万头。日本、加拿大、墨西哥、哥伦比亚等国也相继暴发此病,欧洲多国也开始研究防止PEDV传入的策略[4-5]。
PEDV是尼多病毒目(Nidovirales)冠状病毒科(Coronaviridae)冠状病毒亚科(Coronavirinae)的一员,为有囊膜的单股正链RNA病毒[6]。其S基因编码病毒纤突蛋白(S蛋白),根据其他冠状病毒的S蛋白可以将PEDV的S蛋白划分为S1(1-735aa)和S2(736-1 383aa)两个结构域[7-8]。S蛋白在感染宿主体内介导中和抗体产生的过程中发挥重要的生物学作用[9]。遗传进化分析结果表明,S基因存在较高变异性,不同分离株S基因之间存在不同程度的核苷酸插入、突变和删减等现象,进而改变病毒原始抗原特性,因此常被用来研究不同时间和不同地区流行毒株的亲缘关系[10]。
本研究通过DNA Star、MEGA6等软件对2010年-2014年NCBI上已收录的319株PEDV的S基因进行多序列比对,筛选出6个国家和地区的代表性流行毒株,并对各代表毒株、疫苗株、经典毒株进行比对分析,构建系统进化树。旨在通过对不同流行毒株的序列进化分析,明确PEDV S基因的遗传进化关系,为该病的分子流行病学研究及疫苗株的筛选奠定基础。
1 材料与方法
1.1 材料
1.1.1 基因序列来源 2010年-2014年NCBI上已登录的319株PEDV S基因序列,其中中国流行毒株181株、美国流行毒株98株、韩国流行毒株25株、加拿大流行毒株6株、日本流行毒株6株、越南流行毒株3株。
1.1.2 疫苗株及经典毒株 疫苗株包括中国的CV777细胞株、CH/S沪毒株,韩国的 DR13、KPEDV-9弱毒株,日本的83P-5细胞株。
经典毒株包括欧洲 CV777、Br1/87,韩国的SM98、Chinju99,中国北方毒株 LJB/03、南方毒株JS-2004-2和JS-HZ2012、西部毒株DX和LZC(表1)。
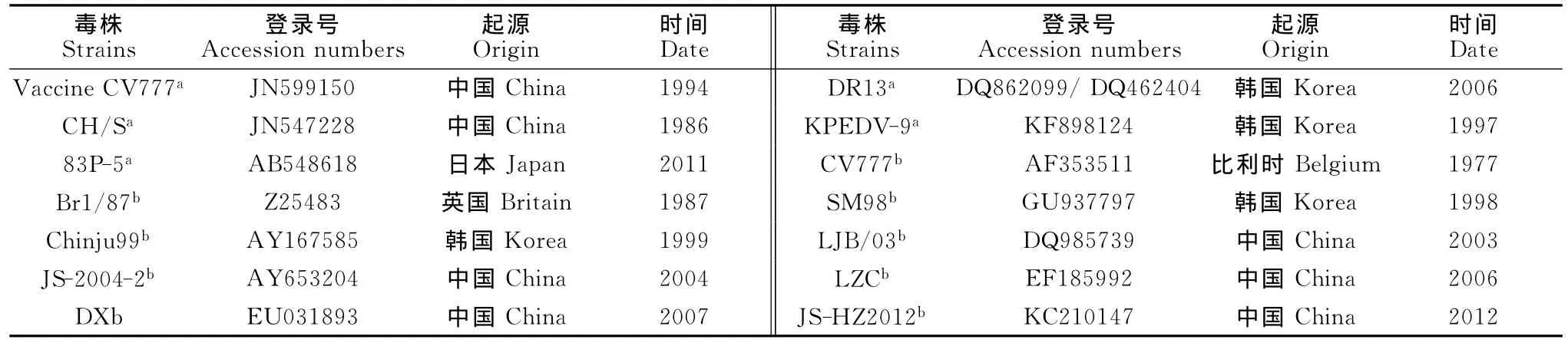
表1 疫苗株和经典毒株的基本信息Table 1 The information of vaccine strains and classical strains from different countries
1.2 方法
1.2.1 代表毒株的筛选 利用MEGA6的Clustal W功能对各国流行毒株进行多序列比对,并应用Neighbor-Joining Tree方法构建系统进化树,根据遗传距离差异性筛选出各个国家和地区的代表性流行毒株。
1.2.2 代表毒株、疫苗株、经典毒株的遗传进化关系 根据1.2.1所确定的各国代表性流行毒株,与临床上常用的疫苗株及经典毒株进行多序列比对,并应用Neighbor-Joining Tree法构建系统进化树。根据遗传进化结果分析2010年-2014年各国PEDV S基因与疫苗株及经典毒株的遗传进化关系。
1.2.3 S基因的序列分析 利用DNA Star中的MegAlign功能对代表毒株、疫苗株、经典毒株的S基因进行序列分析,了解各国代表性流行毒株与经典毒株及疫苗株相比是否存在核苷酸和氨基酸的突变、插入和缺失位点,并明确主要高变区域的结构位置。
2 结果
2.1 代表性流行毒株的筛选
根据不同国家PEDV S基因的遗传进化分析结果,筛选出各国具有代表性的流行毒株。其中中国的181株流行毒株中筛选出42株作为代表毒株,美国的98株流行毒株中筛选出18株作为代表毒株,韩国的25株流行毒株中筛选出8株作为代表毒株,加拿大6株流行毒株中筛选出4株作为代表毒株,日本6株流行毒株中筛选出4株作为代表毒株;越南的3株流行毒株中筛选出2株作为代表性毒株,总计78株,各代表毒株的基本信息见表2。
2.2 代表毒株、疫苗株、经典毒株的遗传进化分析
利用所筛选的78株代表毒株、6株疫苗株、9株经典毒株的S基因构建遗传进化树(图1)。进化树可分为G1、G2两大分支,其中G1可分为G1-1、G1-2两小分支,G2也可分为G2-1、G2-2两小分支。福建株CH/FJND-2/2012为G2-1分支上的唯一成员;美国的3株流行株 (USA/Indiana12.83/2013、USA/Ohio126/2014、USA/Iowa23.57/2013),中国的8株流行株 (CH/FJXM-2、CH13/GX、CH/JLGZL、CH-AHHF、CH/BJSY 、CH/JL 、AH-M 、SD-M/2012),加拿大的 SK/030 株,日本的 MK、83P-5 100th流行株,经典毒株JS-2004-2、LJB/03、DX、CV777、LZC、SM98、Br1/87,疫苗株Vaccine CV777、CH/S、KPEDV-9、DR13、DR13、83P-5同位于G2-2分支上。经典毒株Chinju99和JS/HZ/2012与剩余的63条代表毒株同位于G1上,其中韩国经典毒株Chinju99与韩国的NJ02/2012、AS01/2012、AD0/2012流行毒株,日本的NK、KH野猪毒株同位于G1-2分支上。而较新的经典毒株JSHZ2012与其余的流行毒株同位于G1-1分支上。
2.3 S基因的序列分析
部分毒株氨基酸比对结果表明,S蛋白共存在218处氨基酸位点的突变,其中S1区(1-735aa)存在142个氨基酸突变位点,占突变位点总数的65.60%(143/218),仅S1区的 N′端(1-370aa)就存在86处氨基酸突变位点,占S1区突变位点的60.14%(86/143)。此外,S1区的58-62、140、159-160aa位均存在氨基酸的插入和缺失现象(图2)。综上所述,S蛋白氨基酸位点的突变、插入和缺失现象主要发生于S1区,特别是其N′端。
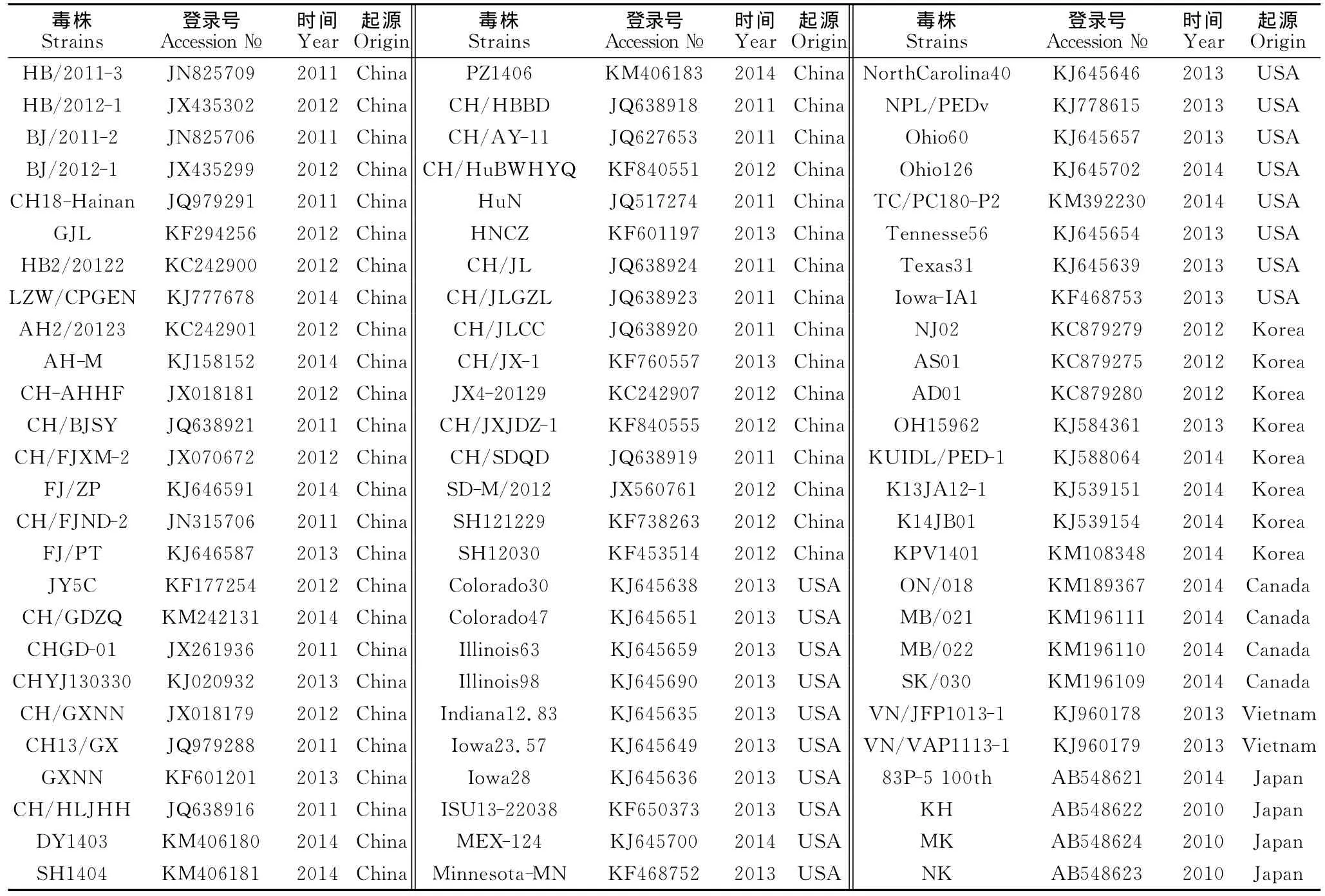
表2 代表毒株的筛选结果Table 2 The screening results of representative strains
3 讨论
大量试验研究表明,PEDV在流行的过程中为更好地适应环境,不同地区PEDV流行毒株的S蛋白会发生不同程度的变异,因此,有必要进一步对PEDV流行毒株的遗传背景进行调查,了解不同国家和地区PEDV S基因的遗传进化关系。本试验通过对2010年-2014年多个国家PEDV流行毒株的S基因进行遗传进化与序列分析,明确近年来PEDV的遗传进化关系,为病原诊断和现地疫苗候选株筛选提供依据。
遗传进化分析结果显示,78株代表性流行毒株中仅有15株流行毒株与疫苗株及经典毒株同位于G2分支上,且没有明显的国籍及时间特征,表明近几年大多数流行毒株与疫苗株及经典毒株的亲缘关系较远。其中流行毒株SD-M/2012、CH/JL/2011、AH-M/2014与中国现用疫苗株CV777亲缘关系最近,位于同一小分支上,笔者推测这3株流行毒株可能为现用疫苗株CV777的变异株。而部分美国毒株(USA/Indiana12.83/2013、USA/Ohio126/2014、USA/Iowa23.57/2013),中 国 的 CH/FJXM-2、CH13/GX、CH/JLGZL、CH-AHHF、CH/BJSY 流行毒株和加拿大的SK/030毒株则与JS-2004-2、DX、LJB-03这3个中国经典毒株亲缘关系较近,推测其可能由这3个经典毒株演变而来。剩余的58株代表性流行毒株与JS-HZ/2012株亲缘关系较近,同位于G1-1分支上,没有明显的国籍及时间特征。但整体观察可知,2010年-2014年的流行毒株与较早的疫苗株及经典毒株遗传距离差异较大,亲缘关系较远,但与近几年的经典毒株亲缘关系较近,说明尽管各毒株间没有时间上的明显规律,但其变异过程仍需一定时间的积累。
通过对PEDV S基因的核苷酸、氨基酸序列进行比对,发现S基因变异性较大,且集中发生在N′末端区域,这一结果与 Lee D K 等[11]对7株韩国PEDV毒株的S基因进行序列分析的结果一致。郑逢梅等[10]通过对12株PEDV流行毒株S、M 和ORF3基因部分核苷酸及氨基酸的序列分析,发现其关键序列明显不同于以往毒株,其中有11株流行毒株的S基因比现用疫苗株CV777多9个核苷酸,且在其S1区N′端存在大量的氨基酸突变。Chen J等[3]通过对除西藏和青海外中国的29个省(市、区)577份腹泻样品进行分子流行病学调查分析,证实33个分离株在S基因N′末端1-370aa存在不同程度碱基的删减或插入现象。
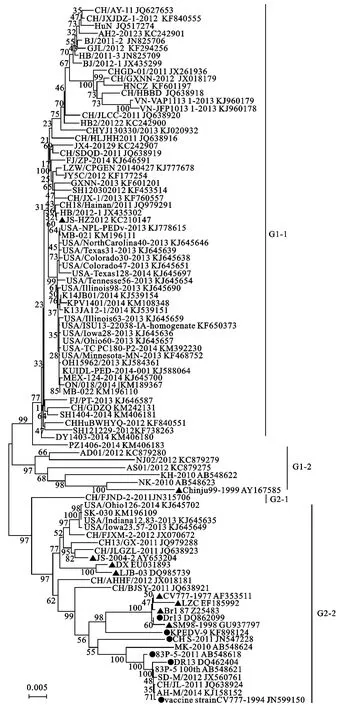
图1 流行毒株、经典毒株、疫苗株的遗传进化分析Fig.1 Phylogenetic analyses of prevailing strains,classic strains,vaccine strains
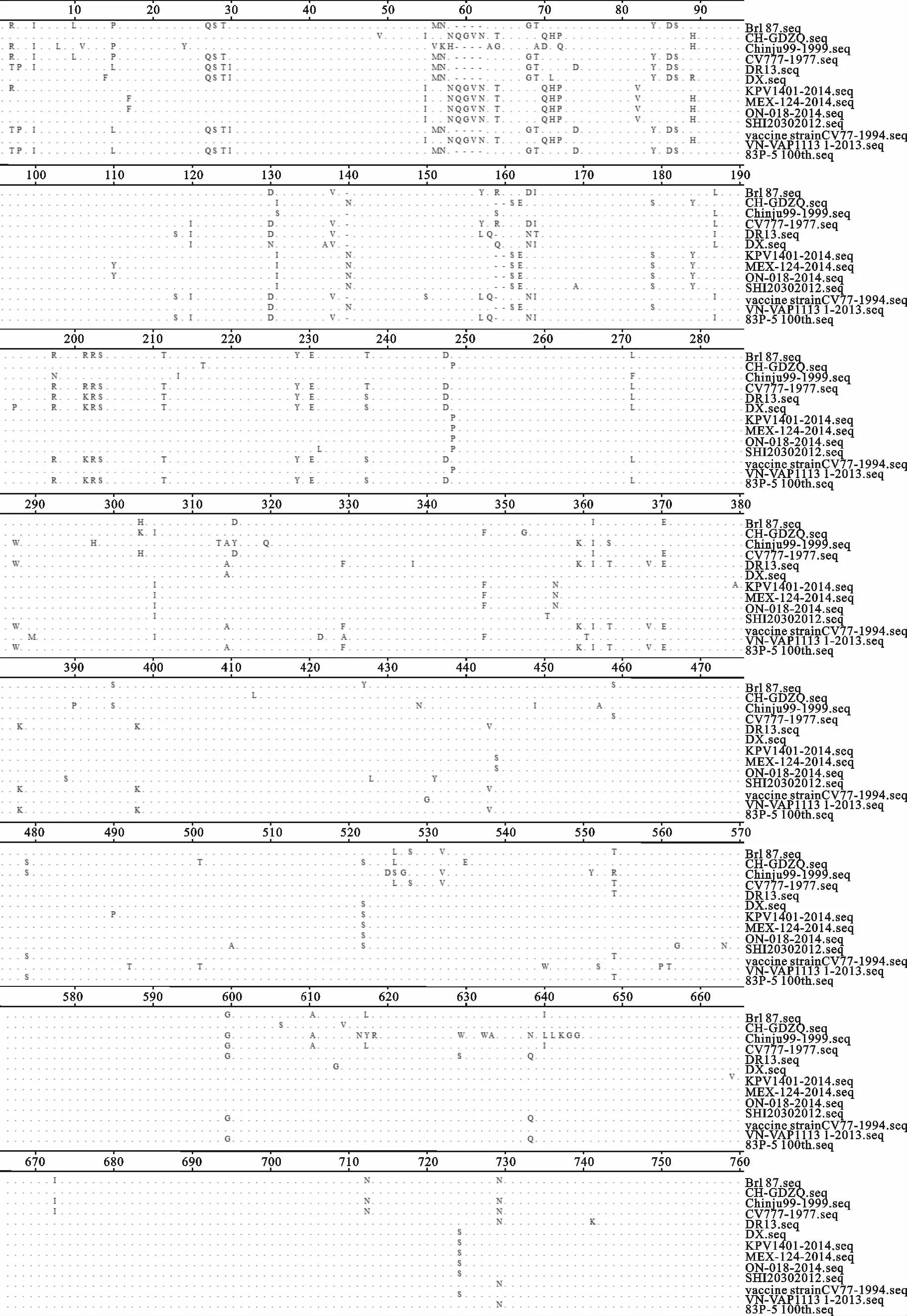

图2 部分PEDV S蛋白氨基酸比对结果Fig.2 Amino acid sequence alignment of the S protein of part PEDV strains
PED是目前危害养猪业最重要的传染病之一,虽未被列入国家农业部重大动物疫病名录,但却严重影响了养猪业的健康发展。自1995年猪传染性胃肠炎(华毒株)与PED(CV777疫苗株)二联弱毒苗及灭活苗在我国投入使用以来,显著降低了该病对我国养殖业造成的损失。但自2006年起,PED开始在免疫猪群中流行[12],特别是从2010年10月开始,中国南部10多个省份暴发大规模的PED,导致超过100万头仔猪的死亡。此后,该病迅速蔓延至全国各个省份,由此造成的发病和死亡率至今仍然无法得到有效的控制[13]。根据各地报道可以看出用于试验研究的猪群大多使用了PED灭活苗或弱毒苗(CV777株),但发病率和未断乳仔猪的病死率依然很高,由此说明疫苗免疫或母源抗体并不足于保证猪只免受PEDV的侵袭。原因可能有两个:①流行毒株发生变异,从而导致免疫失败;②流行毒株毒力增强,导致疫苗或其他途径产生的免疫力并不足以保护猪只免于感染而发病。由于RNA聚合酶缺乏矫正功能,所以RNA病毒容易发生变异。PEDV灭活苗及弱毒疫苗的使用一度使该病的感染显著减少,但可能正是由于疫苗的长期使用从而使得PEDV在免疫压力下发生变异,导致出现免疫逃逸。本试验通过对各代表毒株、疫苗株、经典毒株的遗传进化关系及序列变化进行分析,证实目前全球范围内广泛流行的PED的流行毒株与经典毒株及疫苗株的基因序列发生了很大程度的变异。因此,尽快从当前流行毒株中筛选出新的疫苗候选株将成为防控该病继续蔓延的关键措施之一。
[1]尹宝英,吴旭锦,熊忙利.猪流行性腹泻诊断方法研究进展[J].动物医学进展,2013,34(12):156-159.
[2]Song D,Park B.Porcine epidemic diarrhea virus:a comprehensive review of molecular epidemiology,diagnosis,and vaccines[J].Virus Genes,2012,44:167-175.
[3]Chen J F,Sun D B,Wang C B,et al.Molecular characterization and phylogenetic analysis of membrane protein genes of porcine epidemic diarrhea virus isolates in China[J].Virus Genes,2008,36:355-364.
[4]Jung K,Wang Q H,Scheuer K A,et al.Pathology of US porcine epidemic diarrhea virus Sstrain PC21Ain gnotobiotic pigs[J].Emerg Infec Dis,2014,20(4):662-665.
[5]Wang L Y,Beverly B,Zhang Y.New variant of porcine epidemic diarrhea virus,United States[J].Emerg Infec Dis,2014,20(5):917-919.
[6]Pensaert M B,de Bouck P.A new coronavirus-like particle associated with diarrhea in swine[J].Arch Virol,1978,58:243-247.
[7]Jackwood M W,Hilt D A,Callison S A,et al.Spick glycoprotein cleavage recognition site analysis of infectious bronchitis virus[J].Avian Dis,2001,45(2):366-372.
[8]陈弟诗,任玉鹏,张 斌,等.猪流行性腹泻病毒S基因研究进展[J].动物医学进展,2014,35(7):77-81.
[9]Kang T,Seo J,Kim D,et al.Cloning and sequence analysis of the Korean strain of spike gene of porcine epidemic diarrhea virus and expression of its neutralizing epitope in plants[J].Protein Expres Purif,2005,41(2):378-383.
[10]郑逢梅,霍金耀,赵 军,等.2010年~2012年华中地区猪流行性腹泻病毒分子特征和遗传进化分析[J].病毒学报,2013,29(2):197-205.
[11]Lee D K,Park C K,Kim S H,et al.Heterogeneity in spike protein genes of porcine epidemic diarrhea viruses isolated in Korea[J].Virus Res,2010,149(2):175-182.
[12]Chen J,Wang C,Shi H,et al.Molecular epidemiology of porcine epidemic diarrhea virus in China[J].Arch Virol,2010,155(9):1471-1476.
[13]Sun R Q,Cai R J,Chen Y Q,et al.Outbreak of porcine epidemic diarrhea in suckling piglets,China[J].Emerg Infec Dis,2012,18(1):161-163.
猜你喜欢
区域治理(2022年40期)2022-11-27
科学大观园(2022年2期)2022-01-23
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
中国洗涤用品工业(2019年4期)2019-05-11
中成药(2018年1期)2018-02-02
中成药(2017年3期)2017-05-17
动物医学进展(2015年10期)2015-12-07
动物医学进展(2015年10期)2015-12-07
