
控制水稻红米性状相关基因分子标记的开发
2015-06-05 14:35:08张亭亭窦兰兰王英存李建粤
上海师范大学学报·自然科学版 2015年5期
张亭亭,窦兰兰,王英存,李建粤
(上海师范大学生命与环境科学学院,上海 200234)
控制水稻红米性状相关基因分子标记的开发
张亭亭,窦兰兰,王英存,李建粤
(上海师范大学生命与环境科学学院,上海 200234)
红米因其独特的营养价值日益受到人们关注.红米由两对非同源染色体上的Rc、Rd显性基因控制,且Rc、Rd基因同时存在时,水稻才表现为红米表型.本研究分别在Rc和rc等位基因以及Rd和rd等位基因影响性状的关键性位点上建立了分子标记:Rc(+5150)和Rd(+276).检测结果显示:这2个分子标记可以分别用来区别控制红米性状的Rc/rc和Rd/ rd基因型.开展本研究,为今后利用分子标记辅助快速有效地选育各类红米水稻新品种研究提供了重要的帮助.
水稻;分子标记;红米;Rc基因;Rd基因
随着生活水平的提高,目前人们对稻米的需求已不仅仅满足于解决温饱问题,同时希望吃得更营养更健康.在天然有色水稻糙米中,如红米表皮,含有类黄酮中的重要成员,花色素与糖基结合形成的花色苷.
类黄酮作为一类天然生物活性物质对人体具有多种保健功效,其在增强人体免疫力以及防治多种慢性病等方面的作用越来越受到人们的重视.早在2000年,Middleton等报道[1],黄酮类物质对抵制哺乳动物细胞炎症、心脏病以及癌症方面有较好的效果.黄酮类物质还具有抗氧化、降低血清胆固醇和血清脂质、抑制癌细胞生长和抗癌等生理功能[2-3].黄酮类食物的摄取可以降低由冠心病、心血管疾病和其他原因导致人的死亡危险[4].McCullough等[5]的研究表明,即使摄入极少量黄酮类富集的食物对人体也大有裨益.
红米的米色由两对基因,Rc和Rd决定.这两对基因都通过单基因进行遗传,但同时又具有相互作用,共同参与水稻谷粒种皮中红色色素的淀积.Rc基因编码一个含有bHLH基序的转录调节因子,Rd基因编码花色素合成路径中一个关键分子DFR蛋白.当Rc单独存在,即基因型为RcRcrdrd或Rcrcrdrd的水稻,米色为棕色.当Rd单独存在时,即基因型为rcrcRdRd或rcrcRdrd的水稻,米色为白色.当Rc和Rd两对基因都隐性纯合时,即基因型为rcrcrdrd,米色也为白色.绝大多数白米水稻都是rcrcrdrd基因型.只有当Rc和Rd同时存在,即基因型为Rc-Rd-,米色才能表现为红色[6-7].
本文作者根据Rc和Rd分别在红米和白米中的差异位点,建立了便于检测Rc和Rd两对等位基因的分子标记.开展本研究,可为利用分子标记辅助选育红米水稻新品种研究奠定基础.
1 材料和方法
1.1 水稻材料和总DNA提取
本试验用到4种水稻种子:“Kasalath”(基因型为RcRcRdRd)红米水稻种子、“日本晴”(基因型为rcrcrdrd)白米水稻种子、“上师大6号”(基因型为RcRcRdRd)红米水稻[8]与“上师大5号”(基因型为rcrcrdrd)白米巨胚水稻[9]杂交种子以及再自交收获的种子.分别在如上4种水稻种子长出幼苗后取叶片,采用CTAB法提取基因组DNA.
1.2 分子标记的建立与检测
比较从GenBank查找的“Kasalath”水稻(GenBank:AB250059.1)和“日本晴”水稻(GenBank:DQ885804.1)相应的Rc和rc等位基因序列.在Rc和rc两者具有差异位点的两端设计1对引物.Rc和rc等位基因的PCR检测反应程序为:94℃预变性5 min;94℃变性50 s、59℃退火30 s、72℃延伸20 s,32个循环;72℃保温10 min.
从GenBank查找“Murasaki-ine”水稻(AB003495)[7]的Rd序列,再通过NCBI的blast寻找“日本晴”相应序列(AP004317.1)并进行比较.在两者有差异位点的两端设计一对引物.Rd和rd等位基因扩增的PCR程序为:94℃预变性5 min;94℃变性30s、64℃退火30 s、72℃延伸40 s,32个循环;72℃保温10 min.
各引物均由上海生工生物工程技术服务有限公司合成.
对于Rc和rc等位基因扩增产物,直接采用4%琼脂糖凝胶电泳进行检测.Rd和rd等位基因扩增产物,需要进一步采用Taq I限制性核酸内切酶酶切后,再利用2%琼脂糖凝胶电泳进行检测.Rd和rd等位基因扩增产物酶切体系:PCR产物17μL,Taq I内切酶(TaKaRa公司)1μL(10U),10×Taq IBuffer 2μL,0.1%BSA 2μL,在65℃水浴锅中酶切12 h.
2 结果与分析
2.1 Rc和Rd基因分子标记的建立
比较白米水稻“日本晴”rc基因和红米水稻“Kasalath”Rc基因包括外显子和内含子全长序列发现,不论在外显子和内含子,两者都存在差异.在第7外显子,距离Rc基因起始密码子第一个脱氧核苷酸+ 5150处,“日本晴”的rc基因比“Kasalath”Rc基因缺失14个脱氧核苷酸(图1).
根据Furukawa等研究认为,正是由于这14个脱氧核苷酸的缺失引起Rc基因移码突变,提前出现终止密码子,导致白米水稻的rc基因合成的是一个无功能的截短蛋白[7].因此,本试验将Rc和rc等位基因在第7外显子是否具有14个脱氧核苷酸缺失位点作为筛选Rc/rc的分子标记Rc(+5150),并在其两端设计一对上、下游引物Rc-F和Rc-R,相应的序列分别为:5′CAGTTACAGGGGAGCAGAAAC3′和5′GTACCAAAGATCGCAGAATTATG3′.
比较“日本晴”水稻rd基因和“Murasaki-ine”水稻Rd基因包括外显子和内含子全长序列发现,两者只有一个脱氧核苷酸位点差异.在距离Rd基因起始密码子第一个脱氧核苷酸+276处,“Murasaki-ine”水稻的Rd基因为C,与临近上游+275处脱氧核苷酸T和临近下游+277和+278两处脱氧核苷酸G和A,构成Taq I限制性核酸内切酶识别位点“TCGA”,而白米水稻rd基因+276处脱氧核苷酸为A,不能被Taq I限制性核酸内切酶识别(图2).本试验将该位点作为筛选Rd/rd的分子标记Rd(+276),并在两端设计一对上、下游引物Rd-F和Rd-R,相应的序列分别为:5′ATGGGCGAGGCGGTGAAGG3′和5′TCGTCGTGGTCGTAGGAGGG3′.
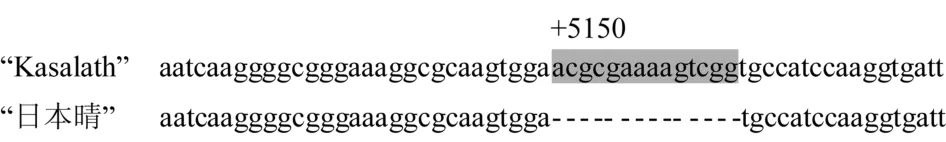
图1 "Kasalath"和"日本晴"水稻部分Rc和rc等位基因序列比较(阴影部分显示为差异位点)
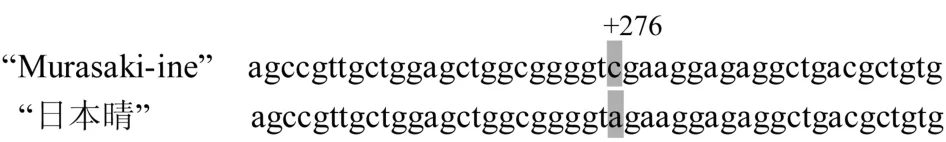
图2 "Murasakiine"和"日本晴"水稻部分Rd和rd等位基因序列比较(阴影部分显示为差异位点)
2.2 Rc和Rd基因分子标记的检测
利用Rc-F和Rc-R一对引物,对分别含有Rc和rc等位基因的“Kasalath”和“日本晴”水稻基因组DNA进行扩增,预计两种水稻都只能扩增出一条带,分子量分别为167 bp和153 bp,而杂交植株同时具有与“Kasalath”和“日本晴”两种水稻相同分子量的两条带.经检测显示,电泳结果与预计完全相同(图3A).由此表明,本试验设计的分子标记及检测方法能够明显地区分出红米水稻和白米水稻分别具有的Rc和rc等位基因.
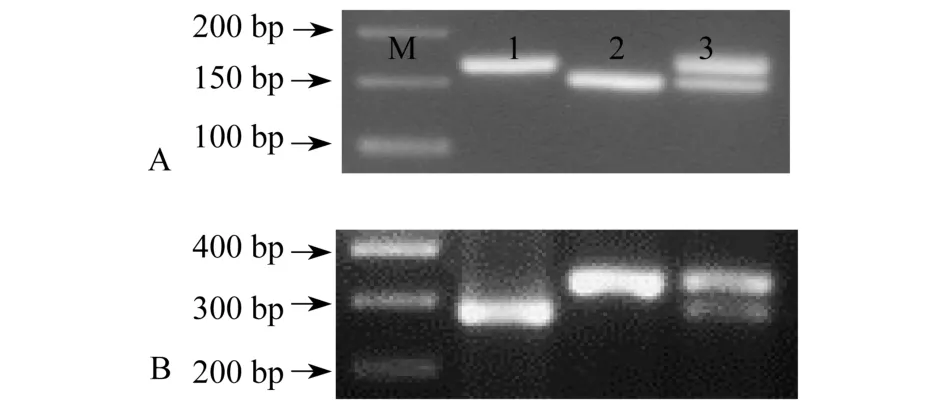
图3 Rc(A)和Rd(B)基因分子标记检测
Rd和rd基因PCR产物预期都为553 bp.Rd基因的扩增产物经Taq I限制性核酸内切酶酶切后,预计会产生分子量分别为275、52、57、132和37 bp 5个片段.由于52 bp和57 bp条带比较接近,预计用2%的琼脂糖凝胶电泳往往会分不开,37 bp条带片段较小,电泳后在凝胶中有可能会看不到;rd基因的扩增产物经Taq I限制性核酸内切酶酶切后,预计会产生分子量分别为327、57、132和37 bp 4个片段.因此,Rd和rd基因PCR产物经Taq I限制性核酸内切酶酶切,再经过2%的琼脂糖凝胶电泳检测后,两者之间的主要差异是在275 bp和327 bp两个条带.
以“Kasalath”红米水稻、“日本晴”白米水稻以及红米和白米杂合植株为材料,对本试验设计的Rd基因分子标记及相应的Rd-F和Rd-R引物和检测方法进行验证.结果显示,“Kasalath”水稻的PCR产物经Taq I酶切后具有275 bp的特征条带(图3B泳道1),“日本晴”水稻的PCR产物经Taq I酶切后具有327 bp的特征条带(图3B泳道2),而杂合植株同时具有275 bp和327 bp的特征条带(图3B泳道3).因此,本试验设计的分子标记及检测方法能够明显地区分出红米水稻和白米水稻分别具有的Rd和rd等位基因.
2.3 Rc和Rd基因分子标记对F2植株的检测
以“Kasalath”和“日本晴”水稻为对照,利用Rc和Rd基因分子标记,检测了124棵由“上师大6号”红米水稻[8]与“上师大5号”白米巨胚水稻[9]杂交再自交获得F2植株的基因型.在F2植株中,如Rc或Rd带型与“Kasalath”水稻相同,表明该植株具有纯合显性的Rc或Rd基因,如带型与“日本晴”水稻相同,表明该植株具有纯合隐性的rc或rd基因,如带型同时具有与“Kasalath”和“日本晴”相同条带,表明该植株的Rc或Rd基因为杂合状态(图4A、B).在124棵自交植株中,对于Rc/rc等位基因,筛选到三种带型植株数分别为:33∶58∶33;对于Rd/rd等位基因,筛选到三种带型植株数分别为:36∶54∶34.经χ2检测显示,Rc基因和Rd基因的分离比,都符合孟德尔一对基因F2植株基因型1∶2∶1的分离比.根据孟德尔的分离规律和自由组合规律,两对基因F2代植株对应4种表型(Rc-Rd-、rcrcRd-、Rc-rdrd、rcrcrdrd)的分离比为9∶3∶3∶1.从分子标记检测显示,4种表型对应的基因型实际观察值为67∶24∶24∶9.χ2检测显示,实际值与理论值也相符.
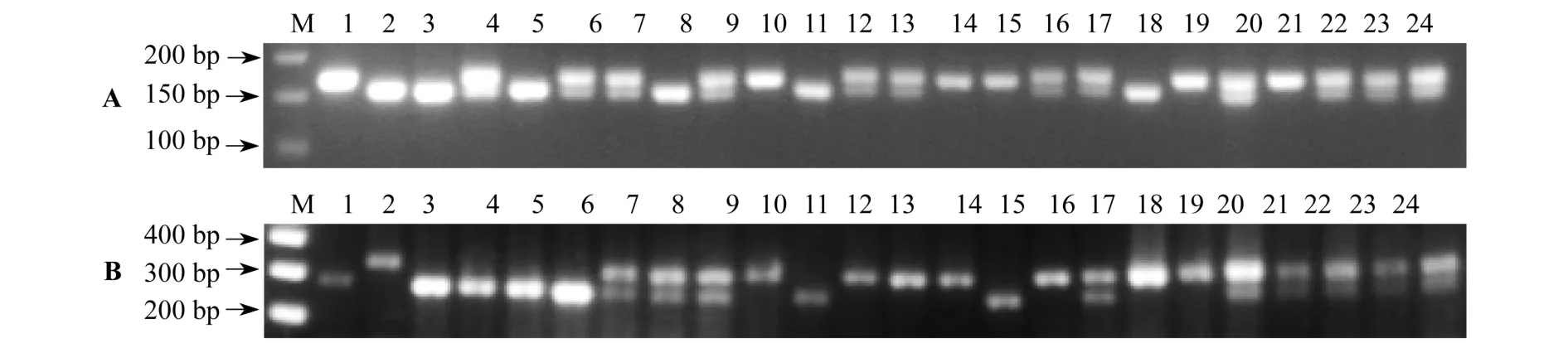
图4 部分F2植株Rc(A)和Rd(B)基因检测
对应比较Rc和Rd两个基因的检测结果,本试验共筛选到9棵与纯合红米水稻品种完全相同基因型的植株.
3 讨 论
在水稻传统育种中,主要根据田间表型对植株进行筛选.由于表型往往受到季节、环境及基因表达与否的影响,以至于个体的表型有可能与其基因型之间存在较大的差异.而且,有些性状,如米色的表型,由于存在母性效应而延迟至下一代才会与基因型一致.所以在水稻常规育种中,通过表型进行筛选,准确性相对较差.分子标记辅助育种选择,是利用分子标记对目标基因进行筛选[10].由于直接针对DNA分子水平上的差异进行筛选,因此,分子标记辅助育种选择,不受环境和目标基因表达的影响,可提高筛选的准确率,还能够缩短育种年限;在另一方面,由于没有引入异源基因,不存在目前争议较大的转基因安全问题[11].
最近几年,利用分子标记辅助选育水稻新品种研究越来越受到育种家的重视,并在水稻分子标记开发[12-15]、水稻食味品质改良[8,16]、提高水稻的抗病性[8]、培育香型巨胚水稻[17]等方面已有较多研究报道.红米水稻因其糙米表面含有对人体具有保健功能的黄酮类物质,属于特种功能水稻.尽管目前已有关于控制红米性状基因相关的研究报道[6-7],但至今为止国内外还未见关于控制红米性状相关基因分子标记开发方面的研究报道.
在目标基因编码序列与决定性状直接相关的位点上建立分子标记,是针对目标基因筛选最为有效的标记类型.本研究建立的红米性状筛选的两个分子标记,都是直接涉及两个目标基因的关键性位点.在本研究中,已利用这样的分子标记筛选获得了Rd和Rc基因都显性纯合的植株,再结合常规育种对纯合植株进行田间农艺及产量性状观察和筛选,即可快速培育出红米水稻新品种.
虽然红米的营养成分含量比普通白米高,但目前采用常规育种培育的红米水稻产量一般比普通白米水稻低.红米性状来源于野生稻,以常规育种选育的红米水稻,其口味一般不如普通栽培稻.如果将本研究建立的分子标记在育种中加以应用,同时结合前人在稻米优质食味品质、抗病、香味等性状建立的分子标记,一同进行辅助选择,并以高产水稻品种作为回交亲本,就有望选育出产量高、口味优良的红米水稻新品种.
Megan等[18]通过比对红米“O.rufipogon”与白米“Jefferson”的Rc/rc等位基因发现,外显子单核苷酸差异位点有8处,还有2处片段的缺失和1处片段插入.Furukawa等也发现红米“Kasalath”水稻和白米“日本晴”水稻,在Rc基因除了单核苷酸碱基差异外,第7外显子处有14个核苷酸缺失,第8外显子有12个核苷酸的缺失和6个核苷酸插入[7].比较白米“日本晴”水稻rd基因和“Murasaki-ine”水稻的Rd基因包括外显子和内含子全长序列显示,两者只有一个核苷酸的碱基位点差异.从进化的角度考虑,对于共同控制水稻红米性状的两个互作基因,Rd比Rc保守.
[1] MIDDLETON E J,KANDASWAMIC,THEOHARIEDSTC.The effects of plant flavonoids on mammalian cells:implications for inflammation,heart disease,and cancer[J].Pharmacol Rev,2000,52(4):673-751.
[2] LAZZW M C,PIZZALA R,SAVIOM,etal.Anthocyanins protectagainst DNA damage induced by tertbutyl-hydroperoxide in rat smooth muscle and hepatoma cells[J].Mutat Res,2003,535(1):103-115.
[3] RUSSO A,FAUCIL L,ACQUAVIVA R,etal.Ochratoxin A-induced DNA damage in human fibroblast:protective effectof cyanidin 3-O-β-D-glucoside[J].JNutr Biochem,2005,16(1):30-37.
[4] MINK P J,SCRAFFORD CG,BARRAJ L M,et al.Flavonoid intake and cardiovascular disease mortality:a prospectivestudy in postmenopausal women[J].Am JClin Nutr,2007,85(3):895-909.
[5] MCCULLOUGH M L,PETERSON J J,PATEL R,etal.Flavonoid intake and cardiovascular diseasemortality in aprospective cohort of US adults[J].Am JClin Nutr,2012,95(2):454-464.
[6] EIGUCHIM,SANO Y.Evolutionary significance of chromosome 7 in an annual type ofwild rice[J].Rice Genetics Newsletter,1995,12:87-189.
[7] FURUKAWA T,MAEKAWA M,OKIT,etal.The Rc and Rd genes are involved in proanthocyanidin synthesis in rice pericarp[J].Plant J,2007,49(1):91-102.
[8] 郭亮,徐申中,张建中,等.聚合蜡质和抗条纹叶枯病基因水稻的分子标记辅助选择[J].西北植物学报,2011,31(8):1537-1542.
[9] 任永刚,张建中,张红梅,等.通过成熟胚离体培养获得巨胚水稻新品种及性状和稻米品质分析[J].上海师范大学学报:自然科学版,2011,40(3):289-294.
[10] 秦延春.DNA分子标记及其在水稻遗传育种研究中的应用[J].广东农业科学,2012(7):145-147.
[11] 王彤彤.水稻分子标记辅助育种研究进展[J].黑龙江农业科学,2012(2):142-145.
[12] 徐小龙,赵国超,李建粤.24种香稻品种甜菜碱醛脱氢酶2基因突变位点的分析及分子标记开发[J].植物分类与资源学报,2011,33(6):667-673.
[13] 许言福,郭亮,沈卫平,等.抗条纹叶枯病基因qSTV11KAS分子标记检测方法的建立以及50种水稻qSTV11KAS基因的分析[J].上海农业学报,2013,29(5):32-35.
[14] SHIY Q,ZHAO GC,XU X L,etal.Discovery of a new fragrance allele and developmentof functionalmarkers for identifying diverse fragrant genotypes in rice[J].Mol Breed,2014,33(1):701-708.
[15] 谢米雪,郭亮,窦兰兰,等.22种上海市重要水稻10个食味品质相关基因的基因型分析[J].上海师范大学学报:自然科学版,2013,42(4):384-391.
[16] 周屹峰,赵霏,任三娟,等.具中等直链淀粉含量的籼型优质不育系浙农3A的选育[J].杂交水稻,2010,25(4):14 -17.
[17] 朱映东,时亚琼,周锋利,等.分子标记辅助选育香型巨胚水稻[J].上海师范大学学报:自然科学版,2013,42(6):623-628.
[18] MEGAN T S,MICHAEL JT,BERNARD E P,etal.Caught red-handed:Rc encodes a basic Helix-Loop-Helix protein conditioning red pericarp in rice[J].The Plant Cell,2006,2(18):283-294.
Development of themolecular markers for identifying red pericarp trait in rice
ZHANG Tingting,DOU Lanlan,WANG Yingcun,LIJianyue
(College of Life and Environmental Sciences,Shanghai Normal University,Shanghai200234,China)
Red rice attracts people′s attention greatly for its special nutritional value.Red pericarp phenotype of rice is controlled by two pairs of dominant genes,Rc and Rd,on non-homologous chromosomes,and rice pericarp will be red only while Rc and Rd genes are both present.In this study we developed twomolecularmarkers,namely Rc(+5150)and Rd(+276),directly on key loci of Rc/rc allele and Rd/rd allele affecting the rice pericarp color.The results show that the twomolecularmarkers could discriminate the different genotypes of Rc/rc and Rd/rd.This study provides an important help for breeding red rice new varieties quickly and efficiently by usingmolecularmarker-assisted selection.
rice;molecularmarker;red pericarp rice;Rc gene;Rd gene
Q 341;S 336
A
1000-5137(2015)05-0485-05
(责任编辑:顾浩然)
10.3969/J.ISSN.1000-5137.2015.05.005
2014-07-26
上海师范大学产学研项目(DCL201103)
李建粤,中国上海市徐汇区桂林路100号,上海师范大学生命与环境科学学院,植物种质资源开发中心,邮编:200234,E-mail:lijianyue01@aliyun.com
猜你喜欢
南方农业(2021年32期)2022-01-14 05:39:30
诗林(2021年2期)2021-11-09 03:46:51
保健医苑(2021年7期)2021-08-13 08:47:58
智慧健康(2021年17期)2021-07-30 14:38:32
现代妇女(2021年1期)2021-02-08 08:34:35
电脑报(2019年8期)2019-09-10 07:22:44
通信产业报(2018年16期)2018-08-20 10:25:40
现代检验医学杂志(2016年5期)2016-08-20 03:16:54
中国粮油学报(2016年5期)2016-01-23 02:45:03
法医学杂志(2015年4期)2016-01-06 12:36:40
