
新疆春小麦光合生理特性及产量形成规律研究
2015-05-23 03:58:27新疆农业大学农学院乌鲁木齐市830052陈明
石河子科技 2015年6期
(新疆农业大学农学院,乌鲁木齐市,830052)陈明
新疆春小麦光合生理特性及产量形成规律研究
(新疆农业大学农学院,乌鲁木齐市,830052)陈明
通过田间试验研究适于新疆北部种植的不同品质类型的春小麦品种花后旗叶光合特性,探讨不同品质类型春小麦品种产量形成特点,以期为新疆专用优质小麦栽培以及品质生态区域的划分提供一定的理论基础。采用单因子随机区组设计,设8个品种处理,进行田间小区试验。从开花至成熟,三种品质类型春小麦叶面积指数由大到小依次为中筋>强筋>中强筋,虽然中强筋品种花后叶面积指数最低,但在灌浆后期光合速率、蒸腾速率以及气孔导度均大于中筋和强筋品种,并且从衰减速率上看,中强筋品种叶片衰老速度最慢,从而使其获得高产。中强筋品种穗粒数、千粒重和产量均大于强筋和中筋品种,而穗数却为最低。通过灰色关联度分析,产量构成因素与产量的关联序为穗粒数(r=0.3 190)>千粒重(r=0.3 156)>穗数(r=0.3 037)。中强筋品种在灌浆后期保持较高的光合速率,叶面积持续期较长,最终获得最高产量。此外,本试验得出增加穗粒数和千粒重可以有效地促进产量的提高。
北疆;春小麦;光合生理特性;产量
小麦是世界三大粮食作物之一,在全世界拥有最大的种植面积和分布范围,并且其总产量和贸易额均为最高。小麦不仅为人类提供了丰富的营养物质,而且小麦作为国家重要的的储备粮食,对满足人们的粮食需求,提高城乡居民物质生活水平,保证社会安定,促进国民经济发展,都有着十分重要的意义[1,2]。小麦品种高产的形成是一系列生理生化过程相互作用的结果[3],其中光合作用及其碳氮代谢是小麦产量形成的主要生理基础。已有大量研究证明了叶片光合速率与小麦籽粒产量的呈显著正相关,在小麦生育后期,由于经常受到高温、干热风等不利因素的影响,容易造成小麦功能叶片的提前衰老,降低叶片光合速率,进而影响小麦产量[4]。薛香等认为[5],小麦旗叶宽度与光合速率呈正相关,而它的着生角度和长度对冠层光合特性没有明显影响。我国小麦优质生产还是一个十分薄弱的环节,新疆由于其特殊的地理位置和气候条件,使其具备了生产优质小麦的良好的资源条件,成为西北地区重要的小麦优势产区[6,7],但在小麦生产上仍存在一些问题,比如品种布局不合理、缺乏系统的高产栽培技术理论等等[8]。因此,研究新疆不同类型春小麦品种花后干物质、可溶性糖及氮素积累动态、光合特性变化以及品质特性,探讨其产量形成及品质特点,对不同类型春小麦合理栽培措施的选择以及品质生态区域的划分具有重大的意义。
1 材料与方法
1.1 试验区基本概况
试验于2010~2011年在昌吉州奇台县进行。奇台县位于新疆维吾尔自治区东北部,昌吉回族自治州东部,地理坐标为东经89°13′~91°22′,北纬42°25′~45°29′,地势南北高、中间低,呈条状马鞍形,地貌类型可分为南部山区(丘陵)、中部平原、北部沙漠、东北部山地丘陵四大部分。属中温带大陆性干旱半干旱气候,冬季长而严寒,夏季短而炎热,春秋季节不明显,气候干燥。中部为冲积平原和沙漠戈壁区,宜农耕地有200万亩,实际耕地面积为150万亩。年平均气温4.7℃,年平均降水量176mm,蒸发量2 141mm,无霜期约为156d,年总日照时数2 280~3 230h。
1.2 试验设计
试验于2010~2011年在昌吉州奇台县进行,土壤地力均匀一致,前茬作物为保护性耕作小麦,试验采用单因素随机区组试验设计,供试春小麦为新疆主栽品种“新春15号(中筋)、新春12号(中筋)、Y-20(强筋)、新春21号(强筋)、新春17号(中强筋)、新春26号(中强筋)、新春27号(中强筋)、新春6号(中强筋)”,每个品种设3次重复,共计24个小区,小区面积3×4=12m2,播种量525万粒/hm2;春小麦行距20cm,播深3~4cm,播种期为4月中旬。试验区边行设置保护行。大田常规管理。
1.3 测定项目与方法
1.3.1 试验测定指标
1.3.1.1 春小麦主要光合特性测定
自开花期开始,对标记的小麦植株测定其旗叶的净光合速率、蒸腾速率、气孔导度、胞间CO2浓度,每个小区测定3株;同时测定旗叶SPAD值以及植株叶面积,每个小区测定10株。
1.3.1.2 春小麦产量以及经济性状测定
在每个小区的样点外随机选取植株10株,在室内进行考种。测定株高、穗长、每穗小穗数、小穗粒数、穗粒数、结实小穗数和千粒重。小区产量实打实收。
1.3.2 春小麦主要光合特性测定方法
旗叶净光合速率(Pn)、胞间CO2浓度(Ci)和气孔导度(Gs)等指标,使用CIRAS-2型便携式光合仪(英国PPSystems公司)测定,测定过程中光强为自然光,测定时间为上午10∶00~13∶30。旗叶SPAD值的测定采用日本产SPAD-502叶绿素仪;采用长宽系数法进行单株叶面积测定。
1.4 数据处理
采用Excel2003,DPS6.50对试验数据进行处理分析,并用Origin8.5作图。
2 结果与分析
2.1 不同品质类型春小麦品种光合生理特性差异
2.1.1 花后叶面积指数变化动态
由图1可知,不同品质类型春小麦品种花后单株叶面积和叶面积指数变化趋势基本一致。不同类型春小麦品种之间进行比较,从开花至成熟,三种品质类型春小麦叶面积指数由大到小依次为中筋>强筋>中强筋,从叶片衰减速率来看,强筋>中筋>中强筋。各品种之间比较,随着花后时间的推移,新春21号、新春12号、新春17号、新春26号和新春27号单株叶面积和叶面积指数均呈不断下降的趋势,而Y-20、新春15号和新春6号的叶面积高峰值出现时间略有推迟,大约出现在花后12d左右,其原因可能是这三个品种对氮肥反应较为敏感,生育后期追施氮肥促进了其叶面积的增长。各品种从开花至花后24d,叶面积指数最大值由大到小依次为新春21号>新春12号>新春26号>新春17号>新春6号>新春27号>新春15号>Y-20;从叶片衰减速率来看,新春17号叶片衰老最慢,Y-20叶片衰老最快。

图1 花后单株叶面积及叶面积指数变化动态
2.1.2 花后旗叶SPAD值变化动态
三种品质类型春小麦品种花后SPAD值均呈“低-高-低”的变化趋势(图2),除新春17号之外,其余品种均在花后12d达到最大值,新春17号则在花后18d左右达到最大值,几乎所有品种在花后24dSPAD值开始迅速下降,其中新春6号下降幅度最大;从开花至成熟,三种品质类型春小麦品种SPAD值大体表现为中筋>中强筋>强筋。

图2 花后旗叶SPAD值变化动态
2.1.3 花后旗叶净光合速率(Pn)和蒸腾速率(Tr)动态变化
由图3可知,不同类型春小麦品种花后净光合速率(Pn)花后变化动态存在一定程度的差异,并且同一类型不同品种间变化趋势也不同,在花后12d之前,不同类型春小麦品种间净光合速率从大到小依次为中筋>中强筋>强筋,花后12d之后,则表现为中强筋>中筋>强筋,但总体呈下降的趋势。强筋各品种自花后6d开始净光合速率(Pn)持续下降;中强筋品种除新春6号之外,其余各品种花后净光合速率(Pn)均呈单峰曲线变化,其中新春17号、新春26号在花后18d净光合速率(Pn)达到最大值,之后开始下降,但新春17号下降相对缓慢,新春27号则在花后12d达到最大值;中筋品种新春12号花后净光合速率(Pn)变化规律同强筋品种,新春15号在花后12d达到最大值。
不同类型春小麦品种蒸腾速率(Tr)变化动态与净光合速率(Pn)基本一致。新春15号,Y-20和新春21号花后蒸腾速率(Tr)自花后6d开始持续下降;新春6号,新春12号和新春26号在花后12d蒸腾速率(Tr)达到最大值,而新春17号和新春27号在花后18d蒸腾速率(Tr)达到最大值。不同品质类型品种之间比较,在灌浆后期中强筋品种(Tr)最高,其次为中筋品种,强筋品种最低。
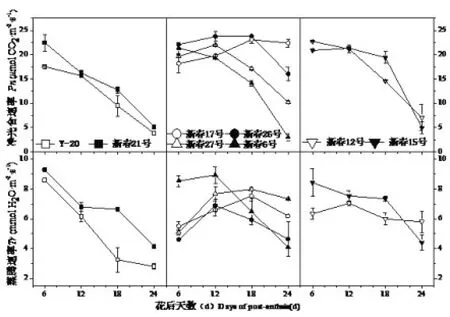
图3 花后旗叶净光合速率(Pn)和蒸腾速率(Tr)动态变化
2.1.4 花后旗叶胞间CO2浓度(Ci)和气孔导度(Gs)动态变化
从图4可以看出,三种品质类型春小麦品种花后气孔导度(Gs)随时间推移逐渐下降,而胞间CO2浓度(Ci)却呈“高-低-高”的变化趋势,不同类型春小麦品种进行比较,花后12d,气孔导度(Gs)基本表现为中强筋>中筋>强筋,而胞间CO2浓度(Ci)在不同类型品种间则未呈现出明显规律。由以上分析可以说明,生育后期光合速率下降的原因并非气孔因素造成,可能是因为叶肉细胞自身光合能力下降,灌浆后期SPAD值降低,Gs也降低,必然导致光合能力下降,进入气孔的CO2并不能被完全利用,这可能也是胞间CO2浓度(Ci)后期升高的原因。
通过对旗叶花后光合特性的分析可以看出,在灌浆后期中强筋小麦之所以能保持较高的净光合速率(Pn),可能是因为在花后12d后中强筋小麦气孔导度(Gs)较高,促进了蒸腾速率(Tr)的加快,而蒸腾速率(Tr)提高有利于叶片表面温度降低,进而促进了净光合速率(Pn)的提高。
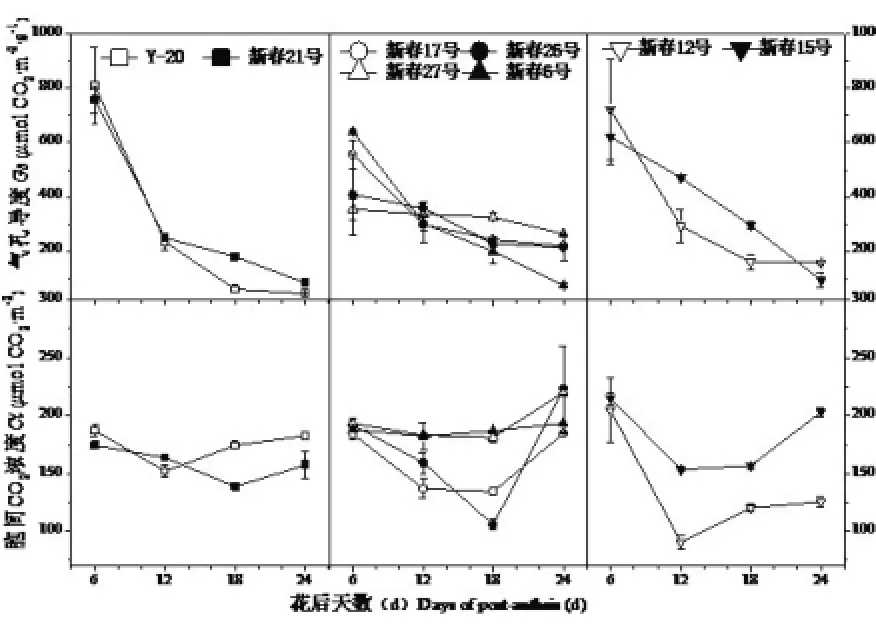
图4 花后旗叶气孔导度(Gs)和胞间CO2浓度(Ci)动态变化
2.2 不同品质类型春小麦品种产量及产量构成因素
由表1可知,强筋、中筋、中强筋三种品质类型穗粒数均值分别为41.43、41.37、45.53,千粒重均值分别为36.77g、40.77g、43.37g,穗数均值分别为480.5、492.0、413.0,产量均值分别为4 403.55kg/ hm2、5 483.61kg/hm2、5 548.61kg/hm2,由以上数据可以看出,中强筋品种穗粒数、千粒重和产量均大于强筋和中筋品种,而穗数却为最低。仅从表1并不能直观的看出产量构成因素间的差异以及与产量的关系大小,因此,可将原始数据进行均值化,使各因素的单位统一以方便进行关联度分析,从关联矩阵(表2)可以看出,产量构成因素与产量的关联序为穗粒数(r=0.3 190)>千粒重(r=0.3 156)>穗数(r=0.3 037),通过以上分析表明,增加穗粒数和千粒重可以有效地促进产量的提高。
3 讨论与小结
小麦产量提高的先决条件是具有良好功能的源,即以叶面积动态变化为主要性状指标。在小麦品种改良过程中,旗叶光合能力和光合面积的同步协调改良是小麦产量持续增加的主要原因,而非光合能力的简单提高[10]。本研究表明,从开花至成熟,三种品质类型春小麦叶面积指数由大到小依次为中筋>强筋>中强筋,虽然中强筋品种花后叶面积指数最低,但最后却获得最高产量,说明不能单一地根据叶面积动态变化来判断小麦产量的高低。

表1 不同类型春小麦品种产量及构成因素

表2 不同类型春小麦品种产量及构成因素关联矩阵
保持较长的光合速率高值持续期,是提高小麦产量的重要途径[11]。本试验结果表明,中强筋品种在灌浆后期光合速率、蒸腾速率以及气孔导度均大于中筋和强筋品种,并且从衰减速率上看,中强筋品种叶片衰老速度最慢,从而使其获得高产。这与前人的研究结果基本一致。
戴明宏等研究表明[12],穗粒数和单位面积穗数是决定产量高低的主要因素,而粒重不能反应出产量的高低。而本研究结果表明,中强筋品种穗粒数、千粒重和产量均大于强筋和中筋品种,而穗数却为最低。进一步进行灰色关联度分析,产量构成因素与产量的关联序为穗粒数(r=0.3 190)>千粒重(r=0.3 156)>穗数(r=0.3 037),由以上分析表明,增加穗粒数和千粒重可以有效地促进产量的提高。
通过以上分析可知,中强筋品种在灌浆后期保持较高的光合速率,叶面积持续期较长,最终获得最高产量。此外,本试验得出增加穗粒数和千粒重可以有效地促进产量的提高。
[1]崔金梅,郭天财等.小麦的穗[M].北京:中国农业出版社,2008:1-15.
[2]于振文.小麦产量与品质生理及栽培技术[M].北京:中国农业出版社,2006:1-12.
[3]高海涛,王育红,孟战赢等.超高产小麦产量及旗叶生理特性的研究[J].麦类作物学报,2010,30(6):1080-1084.
[4]李宏伟,王淑霞,李滨等.早衰和正常小麦近等基因系旗叶光合特性与产量比较研究[J].作物学报,2006,32(11):1649-1655.
[5]薛香,吴玉娥,杨中强等.小麦旗叶性状与光合特性的相关性分析[J].陕西农业科学,2007(4):22-23,44.
[6]叶玉香,何建军,史丰疆.新疆小麦生产发展的限制因素及对策[J].新疆农业科技,2009(2):5.
[7]钟永玲.2011年新疆小麦生产形势及展望[J].农业展望,2011(7):39-41.
[8]李寿山,赵奇,陈兴武等.新疆小麦生产存在的问题及其解决的途径与措施[J].新疆农业科学,2007,44(S1):1-4.
[9]高凤梅,姜东峰,邵立刚.不同品质类型春小麦品种产量形成的动态分析[J].河南科技学院学报(自然科学版),2008,36(1):4-6,21.
[10]王士红,荆奇,戴廷波等.不同年代冬小麦品种旗叶光合特性和产量的演变特征[J].应用生态学报,2008,19(6):1255-1260.
[11]隋娜,李萌,韩伟等.超高产小麦生育后期旗叶生理特性的研究[J].麦类作物学报,2009,29(6):1039-1042.
[12]戴明宏,赵久然,杨国航等.不同生态区和不同品种玉米的源库关系及碳氮代谢[J].中国农业科学,2011,44(8):1585-1595.
S512.1+2
B
1008-0899(2015)12-0013-04
猜你喜欢
作物学报(2022年1期)2022-11-05 08:06:34
河北科技师范学院学报(2022年2期)2022-08-26 08:55:32
农业工程学报(2022年7期)2022-07-09 06:16:32
上海农业科技(2022年3期)2022-06-11 00:50:14
今日农业(2021年3期)2021-12-05 01:46:23
美食(2021年8期)2021-08-18 08:49:18
西藏农业科技(2019年3期)2019-11-04 00:35:12
饮食科学(2019年8期)2019-08-26 02:16:48
今日农业(2019年11期)2019-08-15 00:56:32
现代农业(2016年5期)2016-02-28 18:42:39
