
43份割手密资源农艺性状遗传多样性评价
2015-04-29 00:44:03刘建乐白昌军严琳玲贾庆麟罗灿张瑜
热带作物学报 2015年2期
刘建乐 白昌军 严琳玲 贾庆麟 罗灿 张瑜


摘 要 为揭示割手密种质资源的遗传多样性,对野外采集的43份材料的株高、叶长、叶宽、茎径、节数等15个重要农艺性状进行检测和分析。结果表明,参试材料在这15个性状上都表现出较大的遗传变异,变异系数在9.87%~206.37%之间。主成分分析显示7个主成分对变异的累计贡献率达86.66%,可作材料选择的重要依据。聚类分析将参试材料分为3类,其遗传关系与地理来源没有必然联系。
关键词 割手密;遗传多样性;主成分分析;相关性分析
中图分类号 S566.1 文献标识号 A
割手密(Saccharum spontaneum L.)即甘蔗细茎野生种,又名甜根子草、小巴茅,为禾本科(Gramineae)蜀黍族(Andropogoneae)甘蔗属(Saccharum)多年生草本植物[1]。割手密的适应性强,分布范围广,在热带、亚热带两大气候区均有其踪迹[2]。割手密在甘蔗品种改良中起着重要的作用,是栽培甘蔗种的原始亲本之一,生产种植的甘蔗品种中几乎都有割手密的血缘[3],研究结果表明,甘蔗栽培种中大约有 10%的染色体来源于割手密[4]。因而世界各主要甘蔗生产国都很重视割手密种植资源的收集和保护,同时也开展了许多遗传多样性及核心种质构建的研究[5-6]。
中国是世界上重要的甘蔗原产地和甘蔗种植国,一直非常重视割手密资源的收集、保育及其在甘蔗品种改良上的应用。目前甘蔗育种存在亲本资源匮乏,品种间遗传背景狭窄的困境,利用甘蔗的野生资源割手密,拓展甘蔗亲本的遗传背景,是提高甘蔗亲本的遗传多样性的有效途径[7]。齐永文[8]对广东甘蔗种质资源库中的64份割手密材料的遗传多样进行研究,结果发现,9个性状上都表现出了较高的遗传差异,变异系数介于0.17~0.58,均超过10%。王丽萍等[9]利用割手密和甘蔗杂交、回交手段,获得了高糖后代。李奇伟[3]研究发现利用割手密进行杂交前对其糖分进行目的选择有利于提高其杂交后代的蔗糖分。张革民等[10]对割手密的蔗糖分变异及聚类分析发现糖分遗传变异具有丰富多样性。海南甘蔗育种场利用陵水、崖城、云南的割手密的3个无性系,育成了23个含我国本土割手密血缘的甘蔗品种,甘蔗中10%的遗传物质来源于割手密[4]。邱崇力等[11]、杨荣仲等[12]等分别对我国云南和四川等地割手密种质资源遗传特点进行了研究,各省割手密类型多样,遗传多样性丰富。黄忠兴[13]对国内外456份割手密材料的主要农艺性状进行了多样性分析,虽然材料广泛,但其材料生长在75 cm的水泥圈中,实验设计未设重复。张革明[14]对94份割手密材料的主要数量性状进行了主成分及聚类分析,但是材料来源狭窄(90份采自广西,4份引自云南),实验设计为桶栽。本研究对采自华南8省的43份割手密材料进行大田实验观察,旨在从15个重要农艺性状中筛选出对变异的累计贡献率较大的7个主成分,为将来割手密种质资源的收集保育和创新利用提供参考依据。
1 材料与方法
1.1 材料
1.1.1 自然概况
(1)气候条件。试验田位于海南省儋州市中国热带农业科学院牧草基地,处于北纬19°30′,东经109°30′,海拔134 m;年平均气温23.4 ℃,5~11月高温多雨,12月至来年4月低温干旱,干湿季节明显。试验田年均气温27.3 ℃,历年绝对高温为38.9 ℃,绝对低温6.7 ℃;,年均降水量1 766.2 mm,年平均日照时数1 996.2 h。
(2)土壤条件。试验地土壤为酸性砖红壤,土壤质地较差。经测定,pH5.29,全氮0.069%,有机质1.38%,速效磷1.8 mg/L,速效钾37 mg/L。
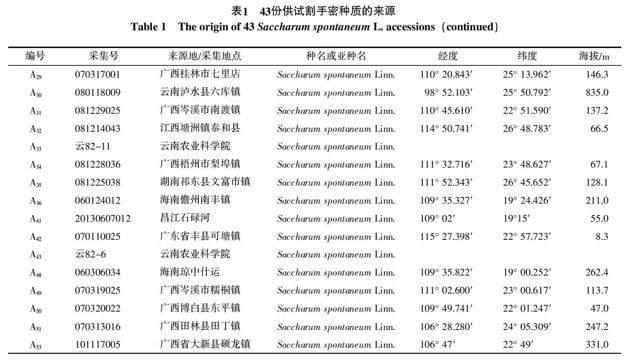
1.1.2 供试材料 供试的43份割手密材料见表1。
1.2 方法
1.2.1 试验方法 试验采用随机区组设计,设3个重复,小区面积为2.0 m×2.0 m,小区间隔1 m,株行距0.4 m×0.4 m,2013年9月26日种植大田,按照常规方式管理,2014年6月16日进行指标调查。每个指标均重复9次求其平均值。
1.2.2 测定项目与方法
株高(cm):测定植株的生理株高。
叶长(cm):叶片肥厚带到叶尖的长度。
叶宽(cm):叶片最宽处的长度。
叶面积(cm2):参照叶面积指数法[15]进行测定,叶面积=0.75×叶长×叶宽。
节数:参照《甘蔗种质资源描述数据规范和数据标准》[16]。
五节长:茎中部最长节连续五节长度。
分蘖率:分蘖率=总分蘖数/基本苗。
茎径:测定植株中部节间的直径。
单株干物质积累(干物质)(g):每个小区选取3株健康完整的植株,齐泥砍下,将茎叶鞘分去,用报纸包好,105 ℃杀青1 h,然后在80 ℃烘至恒重,分别称重。
生长势、黄叶症、眼点病、螟虫、金龟子和嵌纹病参照《甘蔗种质资源描述数据规范和数据标准》[16],略作改动,具体如下:
生长势:4=很好,3=较好,2=中等,1=较差;
黄叶症:3=易感染,2=中抗,1=高抗,0=免疫;
眼点病:3=易感染,2=中抗,1=高抗,0=免疫;
螟虫:3=易感染,2=中抗,1=高抗,0=免疫;
金龟子:3=易感染,2=中抗,1=高抗,0=免疫;
嵌纹病:3=易感染,2=中抗,1=高抗,0=免疫。
遗传多样性指数计算公式为:
H′=-∑Pi×LnPi [17]
1.3 数据处理与分析
采用Microsoft office Excel 2003对数据进行处理,使用SAS 9.0软件计算相关系数,并进行主成分分析和聚类分析。
2 结果与分析
2.1 农艺性状间的遗传多样性分析
从43份割手密资源的15个性状结果可知,割手密种质资源性状差异明显,变异范围广,遗传多样性丰富(表2)。
15个性状的平均变异系数为60.98%,株高、叶长、节数和五节长的变异系数均小于20%,以株高的变异系数最低;干物质、黄叶病、眼点病、螟虫、金龟子和嵌纹病的变异系数均高于50%,以螟虫的变异系数最大。各指标的变异系数大小为:螟虫>黄叶病>金龟子>眼点病>干物质>嵌纹病>分蘖率>生长势>叶面积>茎径>叶宽>节数>叶长>五节长>株高。这表明参试割手密的株高、叶长、节数和五节长等一致性较强,变异的范围不大,相比之下,干物质、黄叶病、眼点病、螟虫、金龟子和嵌纹病等抗虫、抗病性的变异范围更广。
15个性状的平均多样性指数为1.56,其中,螟虫的多样性指数最小(0.70),株高的多样性指数最大(1.98),两者相差1.28,说明参试材料不同性状间变异幅度较大,其余各性状的多样性指数在1.00~1.94之间,结果表明,参试割手密种质的主要农艺性状遗传多样性丰富,在育种方面有巨大的潜力。
2.2 农艺性状间的简单相关分析
利用SAS 9.0软件,对株高、叶长、叶宽及嵌纹病等15个农艺性状指标进行相关分析(表3),结果表明:生长势均和株高呈显著正相关,叶长、叶宽、叶面积、节数、五节长、茎径和干物质均与株高呈极显著正相关,分蘖率和株高呈极显著负相关。这和大田观察结果相似,一般来说,高大植株的材料其叶子比较长,比较宽,干物质积累也比较多,但是分蘖率往往比较低。分蘖率与叶长呈显著负相关,叶面积、五节长、茎径、干物质和生长势均和叶长呈极显著正相关;茎径和叶宽呈显著正相关,叶面积、节数和干物质均与叶宽呈极显著正相关;分蘖率与叶面积呈显著负相关,节数、五节长、茎径、干物质和生长势均与叶面积呈极显著正相关;生长势和节数呈显著正相关,分蘖率和节数呈显著负相关,五节长、茎径和干物质均与节数呈极显著正相关;生长势和五节长呈显著正相关,干物质和茎径与五节长呈极显著正相关,分蘖率和五节长呈极显著负相关;干物质和茎径与分蘖率均呈极显著负相关,一般来说,分蘖率高的植物,不利于植物干物质的积累。生长势和茎径呈显著正相关,干物质与茎径呈极显著正相关,对于禾本科植物,其干物质主要贮存在茎中,茎越粗,其干物质的积累也越大。此外,相关分析还表明生长势和干物质呈极显著正相关。
2.3 农艺性状间的主成分分析
主成分分析结果表明(表4),在所有的主成分构成中,入选的前7个特征值和相应的特征向量累计贡献率达86.66%。其中第1主成分贡献率39.96%为最大,第2、3、4、5、6、7主成分的贡献率分别为11.25%、10.68%、8.57%、7.17%、4.74%、4.30%。因此可以应用这几个主成分对其农艺性状进行概括分析,提取的7个主成分基本上代表了15个指标的绝大部分信息。
从表4、5可知,决定第1主成分大小的主要是株高、叶面积、五节长、茎径和干物质5个性状分量,主成分1相当于5.993 4个原始指标的作用,它反映了原始数据39.96%的信息量。这几个都属于产量性状,是产量性状因子。
在第2主成分的特征向量中,特征向量为正且值较高的性状有黄叶症、螟虫和金龟子。特征向量为负且较高的性状有叶长、叶面积和生长势。说明这一主成分大的材料,抗病、抗虫性较强,叶长、叶面积较小,植物生长势也较慢。因此,选择叶长小,生长势慢的材料,可以有效防止病虫害。甘蔗育种中需要考虑生长势大,栽培上需要考虑降低材料的病虫害。第2主成分主要包含病虫害性状,是抗性性状因子。
在第3主成分的特征向量中,特征向量为正且值较高的性状有分蘖率、生长势、叶宽和叶面积。特征向量为负且较高的性状有螟虫、五节长、眼点病和黄叶症。说明这一主成分大的材料,其生长茂盛,生物质产量高,但其抗虫抗病性较弱。第3主成分是茂盛性因子。
在第4主成分的特征向量中,特征向量为正且值较高的性状有嵌纹病、黄叶病和五节长。特征向量为负且较高的性状有叶宽、叶面积和干物质。说明这一主成分大的材料,其抗病性强,但其干物质产量低。第4主成分是产量性状因子。
第5、6、7主成分的大小主要由螟虫、金龟子、嵌纹病、叶长、五节长和生长势决定。在这3个主成分中,其抗病性越强,五节长和生长势越弱。因此可称其为主茎节数性状因子。
综上所述,入选材料应是第4、5、6、7主成分偏低,第3主成分适中偏低,第1、2主成分较高。即应选择植株高大、茎粗、干物质积累多的材料。经依此筛选,A1和A36两份材料具有较好的综合农艺性状。
2.4 农艺性状的聚类分析
以15个指标作为对割手密农艺性状评价的标准,利用SAS 9.0软件,运用类平均法进行聚类分析(图1),从图1可以看出,通过聚类可将43份种质分为3大类。第Ⅰ类只有A1(云南)一份种质,第Ⅱ类有41份种质,其中广东6份、海南6份、广西11份、云南8份、湖南3份、福建4份、江西2份、贵州1份,第Ⅲ类只有A36(海南)一份种质。从图看出它们的聚类结果与其地理来源没有严格的一致性。从各类群的农艺性状看(表6),第Ⅰ类群植株生长迅速,植株高大,茎粗,叶子长且宽,单株干物质积累量多,但是分蘖率低;第Ⅱ类群植株分蘖率高,抗病、抗虫性强,但是植株矮小,节数较少,不利于单株干物质的积累;第Ⅲ类群的特点大多处于第Ⅰ类群和第Ⅱ类群之间,表现为节数多,植株高大,干物质积累丰富,易染病。
3 讨论与结论
种质资源是作物育种的物质基础,甘蔗育种史上的重要突破均与优异种质资源的发现和利用有关。因此对割手密资源农艺性状的研究,发掘其优异的基因资源,对甘蔗遗传改良具有重要的意义。采用聚类分析方法,将43份供试割手密材料划分为各具特征的三类。说明供试材料表现出多样性和遗传分歧的多向性,这与齐永文[8]对广东割手密品种农艺性状的聚类分析结果一致。本研究认为以植株高大、单株干物质积累量多为目的育种选择亲本应从I类的材料中进行筛选;分蘖率高,抗病、抗虫性强育种选择亲本,应从Ⅱ类的41份材料中进行筛选;要利用割手密节数多的特点,应从Ⅲ类的材料中进行筛选。同时还可根据聚类所划分的类群并结合来源地选择一些育种所需的目标类型的少数材料来构建割手密的核心种质资源库。
遗传变异系数是遗传变异潜力大小的标志,表示群体中直接选择的范围。变异系数大的性状说明从该群体中选出具有该性状的优良个体的几率大,反之则小,因此在选择时,可参考性状的变异情况进行选择。本研究对43份材料的农艺性状进行了分析,结果表明,在所调查的分蘖率、干物质黄叶症等多个性状中,除株高外,其他指标均表现出了较高的遗传变异,变异系数介于14.65%~206.37%之间,其中螟虫和黄叶病的变异系数最高,表明参试材料对螟虫和黄叶症的抗性有较高的变异潜力,在育种利用过程中需要注意选择抗虫、抗病的种质。齐永文等[8]对来自广东地区的64份割手密种质的农艺性状遗传多样性进行评价的研究中,与本研究相同的节数、叶长、叶宽等指标的变异系数均低于0.50,远低于本研究的结果,这可能是因为其实验材料均来自于广东地区,具有较近的遗传距离。另外,黄忠兴等[13]对国内外456份不同地理来源的割手密的农艺性状进行了分析,其中眼点病和金龟子的遗传多样性指数分别为0.97和1.50,与本研究结果相似;而本研究中株高、叶长、叶宽、叶面积、茎径和五节长的遗传多样性指数均处于1.90
张革明等[10]认为,甘蔗育种的最佳途径是要求品种各性状之间的协调发展,不能片面追求单一目标性状的提高。本研究相关分析结果表明,割手密农艺性状间相互关系复杂,彼此间或相互制约,或协调发展,各性状间存在不同程度的相关。因此,选择割手密作为亲本材料时应根据育种目标综合考虑多个性状进行选择。本试验利用主成分分析将割手密主要农艺性状转化为较少的几个主成分,提供了原性状85%以上的信息,且是综合的、相对独立的指标体系,数值直观,容易分析。将主成分分析用于割手密农艺性状的评价和筛选,既能把握其综合性状表现,又能简化选择程序,且更具有科学性。按入选的7个主成分因子分析所得的相应标准筛选,来自云南的A1和海南的A36两份材料符合选择标准,这些材料需要改良的性状相对较少,分别具有各自的优点,可以在甘蔗育种中加以利用。从供试材料间的聚类分析可以看出,43份割手密的遗传聚类与其地理来源没有严格的一致性,产生这种现象的原因可能有2个:一是地区之间存在种质资源的交换,二是不同地方品种来源地的小生境可能存在一定的遗传相似性。特别值得注意的是,A1(云南)与其他供试材料差异明显,单独为一类。与同为云南的A19并没有聚为一类,其原因可能是地理位置导致了基因的变异,A19靠海边,A1靠近山区。
割手密不仅可以为甘蔗育种提供原材料,而且可以用来固土护坡,保持水土。最近几年将割手密作为能源植物研制酒精的报道也越来越多。本研究收集的43份材料中,不同省份材料在数量上差异也较大,如贵州、江西和福建的材料太少。因此,要准确掌握我国割手密分布及遗传多样性的情况,应建立全国性质的割手密种质资源圃,在摸清其遗传多样性规律的基础上加快甘蔗育种和新能源开发新途径。
参考文献
[1] 刀志学, 白史且, 常 丹, 等. 割手密的分子标记应用进展[J]. 草业与畜牧, 2012, 203(10): 16-19.
[2] 杨清辉, 李富生, 肖凤回. 割手密RAPD指纹图谱分析[J]. 云南农业大学学报, 1998, 13(4): 348-350.
[3] 李奇伟. 现代甘蔗改良技术[M]. 广州:华南理工大学出版社, 2000.
[4] Simmonds N W. Sugarcanes in Evolution of Crop Plants[M]. London: Longmans, 1976: 104-108.
[5] Tai P Y P, Miller J D. A Core Collection for Saccharum spontaneum L. from the World Collection of Sugarcane[J]. CropScience, 2001, 41: 879-885.
[6] Amalraj V A, Balakrishnmn R, Jebadhas A W, et al. Constituting a core collection of Saccharum spontaneum L. and comparison of three stratified random sampling procedures[J]. Genetic Resources and Crop Evolution, 2006, 53: 1 563-1 572.
[7] 何顺长, 杨清辉, 肖凤迴, 等. 全国甘蔗野生种质资源采集和考察[J]. 甘蔗, 1994, 1(1): 11-17.
[8] 齐永文, 樊丽娜, 何慧怡, 等. 广东割手密资源农艺性状遗传多样性评价[J]. 甘蔗糖业, 2009(3): 7-10.
[9] 王丽萍, 马 丽, 夏红明, 等. 甘蔗细茎野生种(S. spontaneum)在杂交育种中的利用[J]. 中国糖料, 2006(1): 1-4.
[10] 张革民, 廖江雄, 黄宏套. 广西高糖割手密遗传多样性的表型分析和RAPD分析[J]. 西南大学学报, 2007, 29(8): 83-88.
[11] 邱崇力, 何顺长, 杨世清. 云南割手密(S. spontaneum L.)的研究Ⅱ、 酯酶同工酶的研究[J]. 云南农业大学学报, 1987(2): 83-88.
[12] 杨荣仲, 吴才文, 黄久凯. 四川割手密资源聚类分类研究[J]. 甘蔗, 2001, 8(2): 4-9.
[13] 黄忠兴, 周 峰, 王勤南, 等.国内外割手密资源农艺性状表型遗传多样性分析[J]. 植物遗传资源学报, 2012, 13(5):825-829.
[14] 张革民, 杨荣仲, 刘海斌, 等. 割手密主要数量性状的主成分及聚类分析[J]. 西南农业学报, 2006(6): 1 127-1 131.
[15] 李轩然, 刘 琪, 蔡 哲, 等. 千烟洲针叶林的比叶面积及叶面积指数[J]. 植物生态学报, 2007, 31(1): 93-101.
[16] 范源洪. 甘蔗种质资源描述数据规范和数据标准[M]. 北京: 中国农业出版社, 2006.
[17] 孔繁玲. 植物数量遗传学[M]. 北京: 中国农业大学出版社, 2006.
猜你喜欢
长江蔬菜·学术版(2016年12期)2017-01-12 20:57:59
中国科技博览(2016年25期)2016-12-20 20:04:30
农家科技下旬刊(2016年9期)2016-12-15 10:42:25
现代营销·学苑版(2016年9期)2016-12-08 00:08:32
时代金融(2016年29期)2016-12-05 16:15:54
中国市场(2016年40期)2016-11-28 03:35:20
商(2016年33期)2016-11-24 22:04:19
大学教育(2016年11期)2016-11-16 20:33:18
中小企业管理与科技·上旬刊(2016年10期)2016-11-15 10:22:56
考试周刊(2016年84期)2016-11-11 23:57:34
