
绝经后骨质疏松小鼠中 Hedgehog 信号对破骨细胞的作用
2015-04-25 05:19:34厉晓杰杨柳刘建罗卓荆
中国骨与关节杂志 2015年5期
厉晓杰 杨柳 刘建 罗卓荆
绝经后骨质疏松症是困扰绝经后妇女健康的一大难题,据统计,发展中国家中有约 30% 的绝经后妇女患有骨质疏松[1-2]。高发病率、严重的并发症以及为防治骨质疏松而付出的沉重经济支出更加突出了骨质疏松研究的紧迫性。
绝经后骨质疏松是高转换型骨质疏松,活跃的骨吸收超过骨形成是绝经后骨质疏松中骨量丢失的主要原因[3]。雌激素能抑制破骨细胞的分化并通过雌激素 α 受体促进成熟破骨细胞的凋亡[4]。因此,抑制因雌激素突然缺失而导致的破骨细胞活性过度增强成为治疗绝经后骨质疏松的一种途径。
Hedgehog 信号通路在骨组织中,能调节软骨细胞的增殖和肥大,能促进间充质干细胞 ( BMSC ) 向成骨方向分化[5-6]。在骨转移瘤中,它促进破骨细胞的分化,从而促进骨吸收[7]。而在绝经后骨质疏松中,Hedgehog 信号对破骨细胞的作用尚未研究清楚。
本研究采用体外模拟骨质疏松情况,探究在绝经后骨质疏松的条件下,Hedgehog 信号的改变情况及其对破骨细胞的作用。
材料与方法
一、主要试剂
RAW264.7 细胞购自 ATCC 公司;Murine M-CSF 购自 Peprotech 公司,以 50 ng / ml 浓度使用;RANKL 购自 California bioscience 公司,使用浓度为 100 ng / ml;Purmorphamine 购自 Merck Millipore公司,Vismdegib 购自 Selleck 公司,使用浓度均为1 μm;雌二醇购自 Sigma 公司,使用浓度为 10-8M;无酚红 α-MEM 培养基购自 Gibico 公司;活性炭处理的胎牛血清 ( FBS ) 购自 Hyclone 公司;TRIzol 购自 Invitrogen 公司;RNA 反转录试剂盒购自 Takara公司;SYBR green 染料购自 Takara 公司。
二、绝经后骨质疏松小鼠模型的建立
6 只 8 周大小雌性 C57BL / 6J 小鼠,根据来源不同随机数表法分成两组:绝经后骨质疏松模型( OVX,Ovariotomized ) 组和假手术组,每组 3 只。OVX 组,切开背部皮肤,暴露并切除小鼠双侧卵巢;假手术组,切开背部皮肤,暴露卵巢,切除附近脂肪组织[8]。切除卵巢后 8 周绝经后骨质疏松模型建立完成,处死后取股骨,取骨髓细胞培养。所有动物实验操作均通过第四军医大学伦理委员会通过 ( 20110405-5 )。
三、细胞培养及分组
小鼠股骨干来源的骨髓细胞添加于含 10% FBS和 50 ng / ml 的 M-CSF 的无酚红 α-MEM 培养基中,培养于含 5% CO2、37 ℃ 的恒温培养箱。培养后3 天,取贴壁细胞继续用含有 10% FBS、50 ng / ml的 M-CSF 和 100 ng / ml 的 RANKL 的无酚红 α-MEM培养基培养 6 天,获得破骨细胞[9]。提取两组细胞的总 RNA 并做实时定量 PCR 检测 Hedgehog 信号通路活性标志基因 Ptch1 和 Gli1 的表达水平 (表1)。

表1 实时定量 PCR 引物序列Fig.1 Real-time PCR primers sequence
RAW264.7 细胞以 1.5×104/ cm2密度接种于细胞培养板并随机分为 6 组:雌激素组,给予雌二醇以 10-8M 浓度干预;雌激素+激动剂组,给予10-8M 浓度雌二醇和 1 μm 浓度的 Hedgehog 信号激动剂 Purmorphamine 进行干预;雌激素+拮抗剂组,给予 10-8M 浓度雌二醇和 1 μm 浓度的 Hedgehog 信号拮抗剂 Vismdegib 进行干预;对照组,不给予任何干预;激动剂组,给予 1 μm 浓度的 Hedgehog 信号激动剂 Purmorphamine 进行干预;拮抗剂组,给予 1 μm 浓度的 Hedgehog 信号拮抗剂 Vismdegib 进行干预。各组细胞用含有 100 ng / ml RANKL 的无酚红α-MEM 培养基进行培养 5 天,获得破骨细胞,并进行 RNA 提取和定量 PCR[10]。
RAW264.7 细胞用含有 100 ng / ml RANKL 的无酚红 α-MEM 培养基进行培养 5 天,以进行 TRAP染色分析。随机分为 3 组:雌激素组,给予雌二醇以 10-8M 浓度干预;拮抗剂组,给予 1 μm 浓度的 Hedgehog 信号拮抗剂 Vismdegib 进行干预;对照组,不给予任何干预。
四、RNA 提取和定量 PCR
用 TRIzol 提取细胞总 RNA,然后用反转录试剂盒将 RNA 反转录为单链 cDNA。定量 PCR 采用SYBR Green 染料法以 SYBR Green 染料 12.5 μl、双蒸水 9.5 μl、前后引物各 1 μl、cDNA 样品 1 μl 配成反应体系。利用 real time PCR 反应仪 ( BIO-RAD 公司 ) 检测各组样品中各种基因表达量,并以 GAPDH基因表达量为标准定量。
五、TRAP 染色
培养完成的细胞用 TRAP 试剂盒 ( Sigma ) 进行染色,固定细胞后,蒸馏水冲洗,加入 TRAP 染色液孵育 1 h,蒸馏水冲洗后用苏木精复染,干燥后观察。
六、F-actin 染色
细胞培养完成后弃培养基,用 PBS 冲洗后加入 4% 多聚甲醛固定细胞 10 min,PBS 冲洗,0.1%Triton X-100 PBS 溶液处理细胞 10 min,PBS 冲洗,加入鬼笔环肽 ( Cytoskeleton ) 工作液常温孵育30 min,PBS 冲洗后加入 DAPI 常温孵育 5 min,PBS冲洗后甘油封片。
表2 各组细胞中 Ptch1 和 Gli1 的表达 (±s)Fig.2 The expression levels of Ptch1 and Gli1 in different groups of cells (±s)

表2 各组细胞中 Ptch1 和 Gli1 的表达 (±s)Fig.2 The expression levels of Ptch1 and Gli1 in different groups of cells (±s)
基因 基因表达OVX 组 假手术组Ptch1 0.72400±0.04272 0.44196±0.06822 Gli1 0.66794±0.07331 0.45229±0.05750
表3 各组细胞中 Ptch1 和 Gli1 的表达 (±s)Fig.3 The expression levels of Ptch1 and Gli1 in different groups of cells (±s)
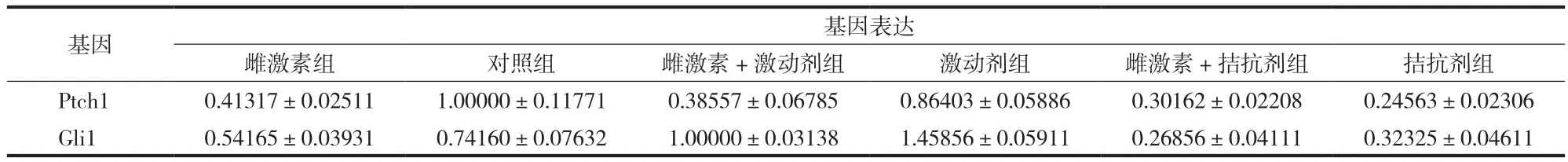
表3 各组细胞中 Ptch1 和 Gli1 的表达 (±s)Fig.3 The expression levels of Ptch1 and Gli1 in different groups of cells (±s)
基因表达雌激素组 对照组 雌激素 + 激动剂组 激动剂组 雌激素 + 拮抗剂组 拮抗剂组Ptch1 0.41317±0.02511 1.00000±0.11771 0.38557±0.06785 0.86403±0.05886 0.30162±0.02208 0.24563±0.02306 Gli1 0.54165±0.03931 0.74160±0.07632 1.00000±0.03138 1.45856±0.05911 0.26856±0.04111 0.32325±0.04611基因
七、统计学处理
所有数值均采用表示,用 SPSS 16.0t检验进行两两组间比较,P<0.05 为差异有统计学意义。
结 果
一、OVX 组和假手术组小鼠成熟破骨细胞中的Ptch1 和 Gli1 表达情况
OVX 组小鼠来源的破骨细胞 Ptch1 和 Gli1 基因表达水平均较假手术组小鼠来源的破骨细胞高,差异有统计学意义 (P<0.05 ) ( 表 2,图 1 )。说明在OVX 组中,Hedgehog 信号活性水平较高。
二、破骨细胞定量 PCR 结果定量 PCR
与对照组相比,雌激素刺激抑制了破骨细胞中Gli1 和 Ptch1 的表达 (P<0.05 )。与激动剂组相比,雌激素+激动剂组中 Gli1 和 Ptch1 表达水平较低(P<0.01 )。以上结果发现,雌激素在破骨细胞中对Hedgehog 信号有抑制作用 ( 表 3,图 2 )。
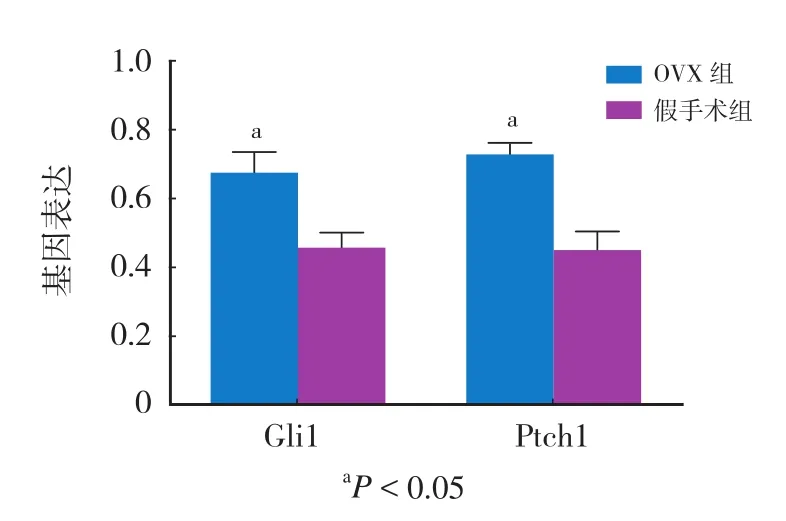
图1 Ptch1 和 Gli1 在不同组细胞中的表达水平Fig.1 Expression levels of Ptch1 and Gli1 in different cell groups

图2 Ptch1 和 Gli1 在不同组细胞中的表达水平Fig.2 Expression levels of Ptch1 and Gli1 in different cell groups
三、TRAP 和 F-actin 染色观察破骨细胞数目
对照组破骨细胞数目较雌激素组多,而拮抗剂组破骨细胞数目较对照组明显减少,且恢复到雌激素存在情况下的正常生理数目。F-actin 染色结果发现,对照组中具有环状细胞骨架的成熟破骨细胞的数目较雌激素组多,拮抗剂组中破骨细胞数目较对照组少。这说明,Hedgehog 信号抑制剂能将雌激素缺乏时过多的破骨细胞数目恢复到雌激素存在时正常水平 (图 3)。
讨 论
绝经后骨质疏松以其高发病率、严重并发症和沉重的社会医疗费用负担成为世界健康难题[11]。与其它类型的骨质疏松不同的是,绝经后骨质疏松为高转换型骨质疏松。雌激素在骨中能维持成骨和破骨活动的平衡,绝经后雌激素水平骤然下降,成骨与破骨平衡打破,破骨细胞介导的骨吸收过度增强成为导致绝经后骨质疏松的重要因素。因此,破骨细胞成为治疗绝经后骨质疏松的重要靶点。
Hedgehog 信号通路对于胚胎发育和多种组织代谢稳态维持都有重要作用[12]。在骨骼系统中,它能促进间充质干细胞向成骨方向分化,能调控软骨细胞的增殖和肥大,在骨转移瘤的情况下,它还能促进破骨细胞的分化。本研究发现,Hedgehog 信号的标志性基因 Ptch1 和 Gli1 在绝经后骨质疏松破骨细胞中表达水平升高。因此,绝经后骨质疏松破骨细胞中 Hedgehog 信号活性增强。为探究 Hedgehog信号增强是否与雌激素水平降低有关,笔者使用RAW264.7 细胞体外模拟绝经后骨质疏松条件,即人为去除或加入雌激素模拟绝经后条件和正常条件。Purmorphamine 和 Vismdegib 分别是 Hedgehog 信号的激动剂和拮抗剂,作用与 Hedgehog 信号的 SMO受体,分别激动和抑制下游转录因子的活化。
本研究结果显示,与雌激素存在时相比,去除雌激素时 Hedgehog 信号活性较高,而且雌激素能减弱 Hedgehog 激动剂对 Ptch1 和 Gli1 表达的促进作用,因此雌激素在破骨细胞中能抑制 Hedgehog信号。以上结果表明,破骨细胞中,雌激素对Hedgehog 信号有抑制作用。这可能是由于正常生理状态下,雌激素维持在正常水平,而正常水平的雌激素对 Hedgehog 信号有一定的抑制作用,进而抑制了破骨细胞的分化成熟。绝经后,雌激素水平骤然下降,雌激素对 Hedgehog 信号的抑制被解除,Hedgehog 活化,进而促进破骨细胞的形成,促进骨吸收,引起骨量下降。当采用 Vismdegib 处理雌激素缺乏情况下的破骨细胞后发现,原过度激活的破骨细胞在 Vismdegib 的作用下被抑制,恢复到雌激素正常情况下的水平。
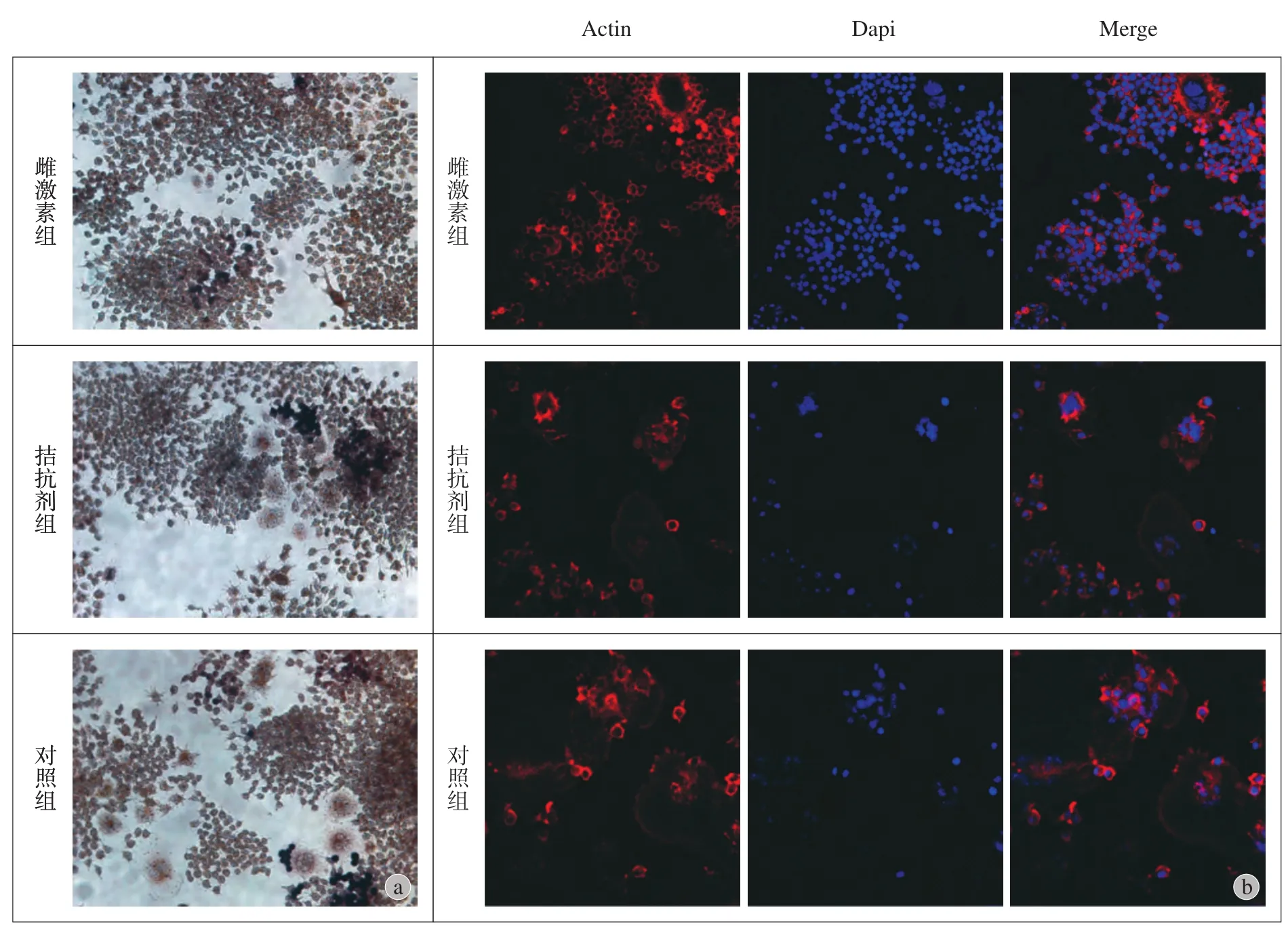
图3 破骨细胞数目比较 a:不同组细胞 TRAP 染色结果;b:不同细胞 F-actin 染色Fig.3 Osteoclasts number in different groups a: TRAP staining; b: F-actin staining
绝经后骨质疏松小鼠破骨细胞中 Hedgehog 信号活性较高;破骨细胞中雌激素可抑制 Hedgehog 信号的活性,使破骨细胞活性恢复至正常情况,从而减少骨吸收,改善绝经后骨质疏松中破骨细胞数目过多的状况;同时,Hedgehog 信号通路抑制剂也具有相同的作用。因此,抑制 Hedgehog 信号通路可以作为治疗绝经后骨质疏松的新靶点。
[1] Reginster JY, Burlet N. Osteoporosis: a still increasing prevalence. Bone, 2006, 38(2 Suppl 1):S4-9.
[2] Sweet MG, Sweet JM, Jeremiah MP, et al. Diagnosis and treatment of osteoporosis. Am Fam Physician, 2009, 79(3):193-200.
[3] Rodan GA, Martin TJ. Therapeutic approaches to bone diseases. Science, 2000, 289(5484):1508-1514.
[4] Nakamura T, Imai Y, Matsumoto T, et al. Estrogen prevents bone loss via estrogen receptor alpha and induction of Fas ligand in osteoclasts. Cell, 2007, 130(5):811-823.
[5] Lanske B, Karaplis AC, Lee K, et al. PTH/PTHrP receptor in early development and Indian hedgehog-regulated bone growth.Science, 1996, 273(5275):663-666.
[6] Hojo H, Ohba S, Yano F, et al. Gli1 protein participates in Hedgehog-mediated specification of osteoblast lineage during endochondral ossification. J Biol Chem, 2012, 287(21):17860-17869.
[7] Das S, Samant RS, Shevde LA. Hedgehog signaling induced by breast cancer cells promotes osteoclastogenesis and osteolysis.J Biol Chem, 2011, 286(11):9612-9622.
[8] 宋佳峻. 糖皮质激素与卵巢摘除后对大鼠骨量及骨代谢影响的对比研究. 中国骨与关节杂志, 2013, (2):84-89.
[9] Heller E, Hurchla MA, Xiang J, et al. Hedgehog signaling inhibition blocks growth of resistant tumors through effects on tumor microenvironment. Cancer Res, 2012, 72(4):897-907.
[10] Sun YL, Chen ZH, Chen XH, et al. Diamagnetic levitation promotes osteoclast differentiation from RAW264.7 cells. IEEE Trans Biomed Eng, 2014, 62(3)900-908.
[11] Hoerger TJ, Downs KE, Lakshmanan MC, et al. Healthcare use among U.S. women aged 45 and older: total costs and costs for selected postmenopausal health risks. J Womens Health Gend Based Med, 1999, 8(8):1077-1089.
[12] Briscoe J. Agonizing hedgehog. Nat Chem Biol, 2006, 2(1):10-11.
猜你喜欢
中国骨质疏松杂志(2024年2期)2024-03-19 09:30:14
中国骨质疏松杂志(2021年9期)2021-10-08 10:07:40
中国临床医学(2019年3期)2019-01-04 09:12:32
中成药(2018年10期)2018-10-26 03:41:22
天然产物研究与开发(2018年6期)2018-07-09 06:01:46
海南医学(2016年8期)2016-06-08 05:43:00
中国社区医师(2015年14期)2015-12-24 00:37:31
医学研究杂志(2015年5期)2015-06-10 06:43:26
生殖医学杂志(2015年11期)2015-02-28 16:32:16
中华皮肤科杂志(2014年4期)2014-12-19 12:55:43
