
水肥耦合对巨能草生长和光合色素的影响
2015-04-15 08:04张明锦陈良华胡相伟徐睿张健
草业学报 2015年5期
张明锦,陈良华,胡相伟,徐睿,张健,2*
(1.四川农业大学生态林业研究所,林业生态工程重点实验室,四川 成都 611130;2.四川农业大学林学院,四川 雅安 625014)
水肥耦合对巨能草生长和光合色素的影响
张明锦1,陈良华1,胡相伟1,徐睿1,张健1,2*
(1.四川农业大学生态林业研究所,林业生态工程重点实验室,四川 成都 611130;2.四川农业大学林学院,四川 雅安 625014)
采用盆栽控制实验的方式,利用控水、控肥的实验方法,设计了干旱(25% FC)、对照(75% FC)和水淹(100% FC)3个水分梯度以及低、中、高3个养分水平的完全随机组合实验,研究了不同水肥耦合条件对巨能草生长与光合色素的影响。结果表明,巨能草的地上生物量、地下生物量、生物总量和根冠比都受到了水肥交互作用的显著影响。在正常水分条件下,高肥处理下的分蘖数、株高、地下生物量、地上生物量和生物总量都是最大的,是9种水肥配比中巨能草生物量积累最佳的水肥配比。可见,良好的水肥条件是巨能草获得高产的前提。水淹和干旱都不利于植株的分蘖和高生长,但干旱条件下可通过施肥提高植株的分蘖能力。水淹环境下,不宜施用过多肥料,中等施肥量最有利于巨能草地下生物量的积累,其根冠比显著增大,有利于植物根系适应水淹条件下的缺氧环境。与水淹条件相比,干旱条件更不利于巨能草地上生物量的积累,为了适应干旱环境,巨能草会把更多的同化物质分配给地下部分,进而增大根冠比,从而表现出较高的生理可塑性以适应极端的干旱环境。有趣的是,水分胁迫下的光合色素含量显著高于正常水分,且随着施肥量的增加,光合色素的含量都有所增加,干旱处理下的增加尤为显著。由此可见,在水分胁迫环境下,巨能草会通过其各种形态和生理适应机制来适应环境,表现出一定的耐涝性和抗旱性,且施肥能够在一定程度上降低水分胁迫对植物生长的影响。
巨能草;水肥耦合;生长;光合色素
在实际生产过程中,人们常常把施肥和灌溉作为主要的农艺措施来管理农作物。但具体施多少肥,浇多少水才能满足植物的生长需求,人们往往凭经验来判断。有时会造成过多的施肥,引起一系列的环境恶化,如营养失衡、土壤板结、水体富营养化等,反而限制了作物的正常生长;施肥太少又不能满足植物的正常生长需求,出现营养不良和缺素症状,影响农作物正常生长和发育,进而影响植物生物量的积累。而在土壤水分方面,则更多地受自然条件的影响,其中干旱和水涝是两种常见的自然逆境。水肥耦合效应概括了水与肥之间的关系,其核心是强调植物生长的两大环境因素“水”和“肥”之间的有机联系。水肥对植物可产生3种不同的耦合效应:协同效应、叠加效应和拮抗效应[1]。因地制宜地调节水分和肥料,使它们处于合理的范围,产生协同作用,达到“以水促肥”和“以肥调水”的目的,这对植物的生长和环境的保护都会有重要的意义。
植物的生长对水分胁迫高度敏感,特别是叶片,轻度的水分亏缺就足以使叶生长显著降低。水分亏缺对植物生长有直接和间接两种效应,直接效应将导致细胞紧涨度降低,使细胞不能增大和正常分裂,间接效应则是通过对光合作用的不利效应而影响生长。而在水涝环境下,其危害不在于水分本身,而在于水分过多引起缺氧,从而产生一系列危害,如乙烯含量增加,植物的有氧呼吸受到抑制,无氧呼吸加强,ATP合成减少,同时积累大量的无氧呼吸产物,如丙酮酸、乙醇和乳酸等导致植物呼吸代谢紊乱[2]。同时,植物体内氧气和二氧化碳等气体扩散和植物对光能的获取受到限制,使水淹成为植物遭受的重要非生物胁迫之一[3]。过度干旱或水涝都会影响植物对养分的吸收,从而影响植物生长和发育。有研究表明,施肥有明显的调水作用,灌水也有显著的调肥作用:作物对矿质离子的吸收是有选择性的,适宜的土壤水肥条件能促进根系发育,扩大根系与土壤的接触面积,有利于增加养分吸收量并通过质流及扩散作用而运输,从而提高作物吸收土壤矿质养分的强度和数量[1]。
巨能草(Pueliasinese)为多年生直立丛生的禾本科植物,由二倍体美洲狼尾草(又称御谷、珍珠谷)作母本和四倍体象草(又称紫狼尾草)杂交选育而成,故又称杂交狼尾草,属三倍体C4植物[4]。我国最早于1982年从哥伦比亚引种到海南岛,引种成功后逐步在全国各地推广[5]。巨能草一年种、多年生,根系生长迅速,可长达3 m以上,在较短时间内形成须根网络,牢固地锁住水分和泥土,可防止水土流失,对绿化荒山荒坡、防风固沙都具有积极作用。目前对巨能草的研究多为其在生态环境治理中的应用推广研究[6],治理的生态环境多为荒山、荒坡、河滩、沙地,但对巨能草在这些极端生态环境下(如干旱的荒山、荒坡、沙地和淹水的河滩、沼泽)的适应性研究还相对缺乏,这些极端的生态环境对巨能草的生长有何影响?巨能草又是如何适应这些极端生态环境?为探究这两个问题,本试验采用盆栽控制实验的方式,通过控水、控肥的方法,测定不同水肥条件下巨能草的生长特征与光合色素含量,探讨不同水肥环境对巨能草生长的影响和巨能草在不同水肥环境下的适应机制,以及不同环境条件下的最佳水肥配比,这将有助于指导不同生境中巨能草的水肥管理,同时也有助于巨能草在生态环境治理、水土保持和种草养畜等方面发挥更大的作用。
1 材料与方法
1.1 试验材料
于2013年5月底选择健康的巨能草草种,将茎干截成长度一致的茎段(约20 cm),将其扦插在装有10 kg混合培养基质(冲积土∶细沙=3∶1,基质含水量为23%,最大含水量为52%)的聚乙烯花盆中,圆台型花盆口径为30 cm,深25 cm,每盆扦插一截,让其在半受控(只遮蔽雨水)的大棚中生长。待其发芽缓苗后,于6月底选择72株高度和长势大致一致、健壮无病虫害的幼苗进行控制实验。土壤背景值如表1所示。

表1 试验土壤背景值
1.2 试验处理
如表2所示,实验采用二因素三水平的完全随机设计: 3个水分梯度(水涝、对照、干旱)×3个养分梯度(2,4,8 g复合肥),使用的复合肥为美丰比利夫复合肥,按照一般作物施肥量200 kg/hm2,考虑到花盆的大小,以及每盆只有1株,所以按照2,4,8 g这样从低到高来设置养分梯度。处理过程为:根据水分梯度将植株分为3组F(水淹处理,淹没到土层表面5 cm)、C(田间持水量75%,作为对照,模拟正常水分条件)、D(田间持水量25%,模拟极端干旱条件),每组24株。再将每组分成L(低施肥量,施肥量为2 g)、M(中施肥量,施肥量为4 g)、H(高施肥量,施肥量为8 g)3个小组,即共有FL、FM、FH、CL、CM、CH、DL、DM、DH九个处理,每处理8个重复。施肥处理是在水分处理1个月后(8月初)进行,以便监测施肥对水分胁迫的效应。实验于2013年6-8月在四川农业大学成都校区生态林业研究所基地防雨棚内进行,在此期间,采用每天称重补水的方法控制盆中土壤水分含量。除水分、养分等实验因素外,其他条件尽可能保持一致。
1.3 测定指标
在实验处理前1 d测定每株植物的初始株高和分蘖数,在收获植株生物量时,测定处理后植株的株高和分蘖数,用处理后的值减去初始值表示株高和分蘖数。8月底收获植株的生物量,测定植株的地下、地上生物量、根冠比、生物总量和光合色素(叶绿素a、叶绿素b、类胡萝卜素)含量。各指标的测定均重复3次。
株高用皮尺量取,用称重法测定生物量。光合色素的测定方法如下:称取剪碎的去中脉的新鲜叶片0.1 g,共3份,分别放入15 mL小试管内。配制丙酮80%与无水乙醇1∶1,即8 mL丙酮与10 mL无水乙醇加2 mL蒸馏水至20 mL,移出3 mL加入放有叶片的小试管内,塑封。将小试管避光静置1 d后,叶片白色。拿出放入分光光度计在663,646和470 nm处进行比色。公式:Ca=12.21A663-2.81A646,Cb=20.13A646-5.03A663,Cx.c=(1000A470-3.27Ca-104Cb)/229,叶绿体色素的含量=色素的浓度×提取液体积×稀释倍数/样品鲜重(mg/g)。
1.4 统计分析
采用SPSS 18.0 for Windows统计软件进行数据统计分析,用单因素方差分析(one-way ANOVA)方法检验9种不同水肥组合对巨能草生长和光合色素的影响;用双因素方差分析(two-way ANOVA)检验水分和施肥及其交互作用对巨能草生长和光合色素的影响。数据整理、计算与作图均采用Microsoft Excel 2007软件进行。
2 结果与分析
2.1 株高和分蘖数
如表3所示,在水淹和干旱条件下,施肥量的多少对巨能草的株高无显著影响;但在正常水分条件下,中、高肥下的株高显著高于低肥处理。从另一角度来看,低肥处理下,不同水分处理间的株高差异不显著;但在中、高肥处理下,正常水分下的株高显著高于干旱处理,与水淹处理无显著差异。
水淹条件下,施肥量的多少对巨能草的分蘖数无显著影响。但在正常水分和干旱条件下,高肥下的分蘖数显著多于低肥处理。从另一角度来看,中、低肥处理下,正常水分下的分蘖数显著高于水淹;在高肥处理下,水淹和干旱下的分蘖数都显著低于正常水分。
2.2 生物量
2.2.1 地下生物量 由图1可见,在正常水分和干旱处理条件下,巨能草的地下生物量呈现有规律的变化,即随着施肥量的增加而增加,但中、高肥间差异并不显著;而在水淹条件下,巨能草在高施肥量下的地下生物量(25.17 g)显著低于中等施肥量(46.56 g)。从另一角度来看,低肥条件下,正常水分下的地下生物量显著高于干旱条件;中肥条件下,不同水分梯度间的地下生物量无显著差异;在高肥条件下,正常水分和干旱条件下的地下生物量显著高于水淹条件,分别是其2.06和1.75倍。总体来看,在正常水分条件下,高肥处理下的地下生物量是最大的,达到了51.96 g,是地下生物量积累最佳的水肥配比。而干旱条件下,低肥处理的生物量只有17.86 g,只有最佳水肥配比下地下生物量的1/3,是最不利于巨能草地下生物量积累的水肥环境。巨能草的地下生物量受到了水肥交互作用的显著影响(P<0.001)。

表3 不同水肥处理对巨能草株高和分蘖数的影响
注:同列不同小写字母表示差异显著(P<0.05)。
Note:The different small letters within the same column mean the significant differences atP<0.05.
2.2.2 地上生物量 如图2所示,正常水分条件下,地上生物量随施肥量的增加而显著增加。水淹和干旱条件下,地上生物量随施肥量的增加也有所增加,但中、高肥处理间的差异并不显著。从另一角度来看,在低肥和高肥条件下,正常水分下的地上生物量均显著高于水淹和干旱条件,但水淹和干旱处理间差异不显著。在中等施肥条件下,不同水分处理对巨能草地上生物量的积累有显著影响,干旱条件下的地上生物量只有59.24 g,而在正常水分条件下,其生物量达到了131.73 g,增加了1.22倍;水淹条件下的生物量(88.65 g)也在其基础上增加了0.5倍。总体上看,水肥协同效应体现在正常水分与中、高施肥量的交互处理下(CM和CH),地上生物量分别达到了131.73和172.34 g,显著高于其他水肥组合,地上生物量受水肥交互影响显著。
2.2.3 根冠比 由图3可知,在正常水分条件下,不同施肥量对巨能草根冠比的影响不显著。但在水淹条件下,施肥量的不同显著影响了巨能草的根冠比,中等施肥量的根冠比达到了0.53,是高肥处理下的2.02倍,低肥处理下的1.4倍。而在干旱条件下,中、高肥处理条件下的根冠比差异不显著,但都显著高于低肥条件。从另一角度来看,在低肥条件下,根冠比不受水分处理的影响。而在中肥条件下,水分处理显著影响着巨能草的根冠比,在水淹和干旱处理下,其根冠比显著高于正常水分条件,分别是其根冠比的1.62和1.94倍。在高肥条件下,水淹和正常水分处理下的根冠比并无差异,但干旱条件下的根冠比显著高于这两种处理,分别是它们的2.18和1.86倍。巨能草的根冠比受到了水肥交互作用的显著影响。
2.2.4 生物总量 如图4所示,在正常水分处理下,生物总量随着施肥量的增加而显著增加;在干旱条件下,生物总量也有所增加,但只有高肥和低肥之间差异显著;在水淹条件下,中、高肥处理下的生物总量显著高于低肥处理。从另一角度来看,在低肥条件下,正常水分下的生物总量(123.02 g)显著高于水淹(81.06 g)和干旱处理(67.65 g),分别是其1.52和1.82倍。在中肥条件下,不同水分处理间的生物总量差异显著:正常水分下的生物总量(174.62 g)分别是水淹(135.24 g)和干旱(96.72 g)处理下的1.3和1.8倍。在高肥下,正常水分处理下的生物总量显著高于水淹和干旱处理,分别是其1.9和1.8倍,但水淹和干旱处理间的差异并不显著。总体来看,水肥协同效应体现在正常水分与中、高施肥量的交互处理下(CM和CH),生物总量分别达到了174.62和224.30 g,显著高于其他水肥组合。生物总量受水肥交互作用的影响显著,正常水分下的高肥环境是巨能草生物总量积累的最佳水肥配比。

图1 不同水肥处理对巨能草地下生物量的影响Fig.1 Effects of different combinations of irrigation and fertilization on below-ground biomass of P. sinese

图2 不同水肥处理对巨能草地上生物量的影响Fig.2 Effects of different combinations of irrigation and fertilization on above-ground biomass of P. sinese
不同小写字母表示差异显著(P<0.05),下同。The different small letters mean the significant differences atP<0.05,the same below.

图3 不同水肥处理对巨能草根冠比的影响Fig.3 Effects of different combinations of irrigation and fertilization on the root-shoot ratio of P. sinese
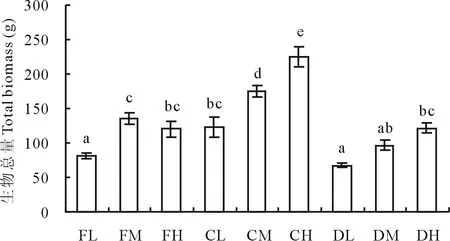
图4 不同水肥处理对巨能草生物总量的影响Fig.4 Effects of different combinations of irrigation and fertilization on total biomass of P. sinese
2.3 光合色素

图5 不同水肥处理对光合色素含量的影响Fig.5 Effects of different combinations of irrigation and fertilization on photosynthetic pigments contents

图6 不同水肥处理对叶绿素a/b的影响Fig.6 Effects of different combinations of irrigation and fertilization on chlorophyll a/b content
2.3.1 光合色素含量 如图5所示,水淹条件下,中、高肥处理间的叶绿素a、叶绿素b和类胡萝卜素含量的差异并不显著,但都显著高于低肥处理。正常水分条件下,高肥处理下的叶绿素a、叶绿素b和类胡萝卜素都显著高于低肥处理。干旱条件下,叶绿素a、叶绿素b和类胡萝卜素的含量都随着施肥量的增加而显著增加,高肥下叶绿素a含量达到了1.62 mg/g FW,是低肥处理(0.77 mg/g FW)的2倍,中肥处理(1.15 mg/g FW)的1.4倍;高肥下叶绿素b的含量高达0.31 mg/g FW,而低肥处理下的只有0.14 mg/g FW,只有高肥处理的1/2。类胡萝卜素的含量在低肥处理下只有0.19 mg/g FW,高肥下其含量增加了89.72%,达到了0.37 mg/g FW。
从另一角度来看,低肥条件下,水分处理对叶绿素a含量的影响不显著;但水淹处理下的叶绿素b和类胡萝卜素含量都显著高于正常水分。中肥条件下,水淹和干旱处理下的叶绿素a、叶绿素b和类胡萝卜素含量都显著高于正常水分处理。高肥条件下,水分处理显著影响了巨能草叶绿素a、叶绿素b和类胡萝卜素的含量,其中正常水分条件下的含量最低,分别只有0.89,0.17和0.23 mg/g FW;水淹处理下的其次,分别为1.17,0.23和0.28 mg/g FW,分别是正常水分处理下的1.32,1.32和1.25倍;干旱处理下的含量最高,分别达到了1.62,0.31和0.37 mg/g FW,分别是正常水分处理下的1.82,1.81和1.63倍。
总体来看,叶绿素a、叶绿素b和类胡萝卜素在不同水肥处理间的变化趋势极其相似,只是在量上有所差异。干旱条件与高施肥量的交互处理下(DH),水肥的协同效应表现极为明显,该处理下的光合色素含量显著高于其他水肥处理。
2.3.2 叶绿素a/b 如图6所示,水淹条件下,不同施肥处理间叶绿素a/b无显著差异。正常水分和干旱条件下,叶绿素a/b受施肥量的影响显著(P=0.038,P=0.015)。正常水分下,高肥下的比值显著低于中肥,但低肥下的比值与高肥和中肥之间无显著差异。干旱条件下,低肥下的比值显著高于中、高肥,但中、高肥之间差异不显著。从另一角度来看,低肥处理下,叶绿素a/b受不同水分处理的影响显著,随着水分含量的减少,其比值显著增大。干旱下的比值(5.57)与水淹条件下的比值(5.03)相比,增加了10.69%。中肥条件下,正常水分和干旱处理下的比值显著高于水涝。但高肥条件下,不同水分处理间叶绿素a/b无显著差异。
3 讨论
分蘖数和株高是两个重要的农艺性状。分蘖数是禾本科等植物在地面以下或近地面处所发生的分枝,产生于比较膨大而贮有丰富养料的分蘖节上。株高则是植物营养生长的重要性状之一。作为重要的新型能源牧草,人们主要是利用巨能草的地上部分,利用其较强的分蘖能力和生长能力,通过多次刈割收获其生物量,增加其经济价值。但分蘖数和株高受水、肥等多种因素的综合影响,条件适当,分蘖数才多,植株才能正常生长。本研究表明,巨能草的株高和分蘖数受水分和施肥单因素的影响显著,但受水肥交互作用不显著(P>0.05)(表4)。水淹不利于植株的分蘖和高生长,即使增加施肥量,也不能显著增加分蘖数和株高;干旱条件同样也会抑制植株的高生长,但可通过施肥提高植株的分蘖能力。在正常水分下,增加施肥量有利于植株的分蘖和高生长。邵玺文等[7]的研究也得出了类似的结论:分蘖期水分胁迫严重制约着水稻分蘖的正常进行,分蘖少,生长量不足,从而影响水稻产量性状而降低产量。杨贵羽等[8]在研究不同土壤水分处理对冬小麦根冠生长的影响时,发现当冬小麦遭受干旱胁迫时,作为冠层结构体现者之一的茎秆也受到抑制。造成此现象的可能原因是[9]:在微观上,由于水分的亏缺,茎秆内细胞膨压下降,细胞壁的伸张受到限制;在宏观上,由于水分胁迫,冠层光合同化物减少,同化物积累降低。因此,水分胁迫通常会造成植株生长变慢或几乎停止,使整体的株高降低。

表4 水分、施肥及水肥交互作用对巨能草各参数效应的显著性检验
P>Fw,不同水分处理间差异的显著性概率;P>Ff, 不同施肥量处理间差异的显著性概率;P>Fw×f,水分和施肥交互作用对差异的显著性概率。*,P≤0.05; **,P≤0.01; ***,P≤0.001; ns, 无显著差异。
P>Fw, probability of the significant difference between different water treatment;P>Ff, probability of the significant difference between different water treatment;P>Fw×f, probability of the significant difference between interaction of water and fertilizer.*,P≤0.05; **,P≤0.01; ***,P≤0.001; ns, no significant difference.
水分作为影响植物生长发育的主要因素之一,影响着植物的各个生理生化过程,最终在宏观上以植物各部分生物量的累积、产量的高低直接体现。作为水分捕获器官的根系及物质能量的供应器官的冠层,均协同影响着植物产量的形成[8],根冠比的大小则反映了植物地下部分与地上部分的相关性。有研究表明[10]:根冠是一个相互作用的共同体,在二者的消长变化及干物质的分配关系中水分扮演着重要角色。植物根系不仅是植物吸收水分和盐类的主要器官,而且是多种物质同化、转化和合成的重要器官[11],并且根系能最早最直接地感受到土壤水分含量的变化,从而对干旱胁迫作出迅速反应,这是植物抗旱性的基础。在水分胁迫条件下,同化物优先供根系生长以吸收更多的水分,来满足作物的生长。徐炳成和山仑[12]通过对苜蓿(Medicagosativa)和沙打旺 (Astragalusadsurgens)在苗期需水及其根冠比的研究就发现根系占总生物量的比重随着土壤水分的降低而增大。在本研究中也发现:巨能草的地上生物量、地下生物量、根冠比和生物总量都受到了水肥交互作用的显著影响(表4)。正常水分下,增加施肥量有利于巨能草地下和地上生物量的增加,但对其根冠比无影响。水淹条件下,高施肥与中等施肥相比,其地上生物量的增产效果并不显著,且中等施肥量更有利于巨能草地下生物量的积累,其根冠比显著增大,有利于植物根系适应水淹条件下的缺氧环境。因此,在水淹环境下,中等施肥量即可。与水淹条件相比,干旱条件更不利于巨能草地上生物量的积累,但中、高肥有利于巨能草根系的生长,进而促进地下生物量的积累,其根冠比显著增大。由此可见,巨能草在干旱环境条件下表现出一定的适应性,通过增加生物量向根系的分配(即增大根冠比),扩大根系表面积,便于吸取土壤深层的水分,以维持体内水分平衡[13-14]。同时也说明了在干旱条件下,施肥能够在一定程度上降低水分胁迫对植物生长的影响[15]。
植物产量的高低首先取决于光合作用系统的面积和效率,植物的光合作用离不开光合色素的参与。光合色素能够对光能进行吸收、传递和转化,其含量直接影响着植物的光合能力,其中叶绿素(Chl)和类胡萝卜素(Car)与光合作用的关系较密切。绝大多数叶绿素a,全部叶绿素b,类胡萝卜素组成辅光系统,负责收集光能并将其传递给中心色素,极少数处于特殊状态的叶绿素a与蛋白质结合为复合体,形成反应中心,引起原初光化学反应。类胡萝卜素作为光吸收复合体的重要组成部分,除了吸收传递光能的作用,还可保护光合器官,防止光的氧化损伤,是植物体内抗氧化系统的重要组成物质[16]。有研究表明:随着干旱胁迫时间的延长,叶片叶绿素降解加剧,活性氧和膜脂过氧化产物含量明显增加[17-18];淹水处理也会导致植株叶片的叶绿素含量呈逐渐下降趋势[19-20]。但也有研究表明,淹水或干旱处理并不会减少植物叶片的光合色素含量[21-22],干旱胁迫下抗旱性强的植株其叶绿素含量和类胡萝卜素含量显著增加[23]。从本研究来看,叶绿素含量并未因水分胁迫而下降,相反地,水分胁迫下的叶绿素a(除低肥条件下)、叶绿素b和类胡萝卜素含量都显著高于正常水分。无论是正常水分还是水分胁迫下,随着施肥量的增加,光合色素的含量都有所增加,干旱处理下的增加尤为显著。这可能与巨能草的耐涝和抗旱机制以及施肥效应有关:在水分胁迫下,植物通过增加类胡萝卜素含量,保护光合器官,防止光的氧化损伤;同时,因为氮是叶绿素等化合物的主要组成元素之一,在一定浓度范围内,氮含量的增加能提高叶绿素等化合物的含量。在本研究中,叶绿素a、叶绿素b和类胡萝卜素的含量都随着施肥量的增加显著增加,这说明施肥能够在一定程度上降低水分胁迫对植物光合作用的影响。低肥条件下,干旱处理下的叶绿素a/b显著高于正常水分,说明巨能草具有较强的耐旱性,通过保持较高的叶绿素a/b维持一定的光合作用[24]。
本研究探讨了不同水肥耦合方式对巨能草生长和光合色素的影响。总体来看,巨能草的地上生物量、地下生物量、总生物量和根冠比都受到了水肥交互作用的显著影响(表4)。在正常水分条件下,高肥处理下的分蘖数、株高、地下生物量、地上生物量和生物总量都是最大的,是生物量积累最佳的水肥配比。从生物量积累和光合色素含量的变化来看,干旱条件下,巨能草会通过增大根冠比和光合色素含量来适应干旱逆境。同时,施肥能够在一定程度上降低干旱胁迫对植物生长的影响,缓减干旱所带来的负面影响。水淹环境下,中等施肥即可,该环境条件有利于巨能草地下生物量的积累,其根冠比也显著增大,有利于植物根系适应水淹条件下的缺氧环境。与干旱条件相比,水淹条件造成的生物量积累损失较小。可见,巨能草会通过其各种形态和生理适应机制来适应干旱和水淹环境,表现出一定的耐涝性和抗旱性,即可应用于干旱的荒山、荒坡治理,也可以应用于河滩地等可能遭到积水的区域。
[1] Xiao Z T, Jiang W J, Yu H J. Research progress in crop water and fertilizer coupling effect. Crops, 2007, (6): 18-22.
[2] Yu F Y, Xu X Z. A review on plant stress physiology. World Forestry Research, 2003, 16(5): 6-11.
[3] Tan S D, Zhu M Y, Zhang K R,etal. Effect of submergence on the antioxidative enzymes and carbohydrate contents ofPaspalumdistichum. Acta Prataculturae Sinica, 2013, 22(1): 217-224.
[4] Magcale-Macandog D B, Predo C D, Menz K M,etal. Napier grass strips and livestock:a bioeconomic analysis. Agroforestry Systems, 1998, 40(1): 41-58.
[5] Xie Y L, He G W, Xiong J P. The production performance and utilization ofPennisetumhydridum. Sichuan Animal and Veterin Sciences, 1999, (4): 23.
[6] Ma C J, Liu F G. The research progress ofPennisetumhydridumin the application of ecological environmental governance. Soil and Water Conservation in China, 2012, (1): 41-44.
[7] Shao X W, Ruan C C, Zhao L P,etal. Effects of water stress on growth and yield of rice in tillering stage. Journal of Jilin Agricultural University, 2005, 27(1): 6-10.
[8] Yang G Y, Luo Y P, Li B G,etal. Effect of different soil water conditions on growth of root and shoot of winter wheat. Agricultural Research in the Arid Areas, 2003, 21(3): 104-109.
[9] Arulselvi S, Selvi B. Genetic diversity of seedling traits conferring drought tolerance in pearl millet. Madras Agriculture Journal, 2009, 96(1-6): 40-46.
[10] Kramer P J.Water Deficit and Plant Growth. Water Relations of Plants[M]. New York: Academic Press, 1983: 342.
[11] Li C S, Liu P, Cai M Z,etal. Physiological response of buckwheat to acid-aluminum stress in growth. Journal of Soil Water Conservation, 2005, 19(3): 105-109.
[12] Xu B C, Shan L. A study comparing water use efficiency and root/shoot ratio of Alfalfa andAstragalusadsurgensat seedling stage. Acta Agrestia Sinica, 2003, 11(1): 78-82.
[13] Lai Y P, Li J, Zhang Z Q,etal. Grey correlation analysis of morphological traits related to drought tolerance of wheat at seedling stage. Journal of Triticeae Crops, 2009, 29(6): 1055-1059.
[14] Muhammad A A, Amjad A, Shahid N,etal. Morpho-physiological criteria for drought tolerance inSorghum(Sorghumbicolor) at seedling and post-anthesis stages. International Journal of Agriculture and Biology, 2009, 11(6): 674-680.
[15] Jiang H J, Li J L, Li L X. Effect of water and fertilizer coupling on ecological characters ofFestucaarundinaceaunder heat stress. Pratacultural Science, 2009, 26(7): 152-157.
[16] Pan R Z, Wang X J, Li N H. Plant Physiology[M]. Beijing: Higher Education Press, 2008.
[17] Cao H, Wang X W, Han Z H,etal. Relationship between changes of endopeptidases activity and active oxygen inMalushupehensisleaves during senescence induced by water stress. Scientia Agricultura Sinica, 2004, 37(2): 274-279.
[18] Qian Y Q, Sun Z Y, Han L,etal. Response of reactive oxygen and its scavenging system in leaves ofBuchloedactyloides(Nutt.) engelm to water stress. Acta Ecologica Sinica, 2010, 30(7): 1920-1926.
[19] Xia Y. Effects of water stress on changes of proline and chlorophyll in fruit crops. Journal of Gansu Agricultural University, 1993, 28(1): 26-31.
[20] Wang R, Liang K L, Zhou Z Y,etal. Effect of different waterlogging stress conditions on growth and some physiological characteristics. Acta Prataculturae Sinica, 2012, 21(1): 149-155.
[21] Pezeshki S R, Pardue J H, Delaune R D. Leaf gas exchange and growth of flood-tolerant and food-sensitive tree species under low soil redox condition. Tree Physiology, 1996, 16: 453-458.
[22] Chen L H, Zhang S, Zhao H X,etal. Sex-related adaptive responses to interaction of drought and salinity inPopulusyunnanensis. Plant, Cell and Environment , 2010, 33: 1767-1778.
[23] Xu X, Peng G Q, Wu C C,etal. Drought inhibits photosynthetic capacity more in females than in males ofPopuluscathayana. Tree Physiology, 2008, 28: 1751-1759.
[24] Zhang M S, Tan F. Relationship between ratio of chlorophyll a and b under water stress and drought resistance of different sweet potato varieties. Seed, 2001, (4): 23-25.
参考文献:
[1] 肖自添, 蒋卫杰, 余宏军. 作物水肥耦合效应研究进展. 作物杂志, 2007, (6):18-22.
[2] 喻方圆, 徐锡增. 植物逆境生理研究进展. 世界林业研究, 2003, 16(5): 6-11.
[3] 谭淑端, 朱明勇, 张克荣, 等. 水淹对双穗雀稗抗氧化酶活性及碳水化合物含量的影响. 草业学报, 2013, 22(1): 217-224.
[5] 谢永良, 何光武, 熊建平. 皇竹草生产性能与开发利用. 四川畜牧兽医, 1999, (4): 23.
[6] 马崇坚, 刘发光. 皇竹草在生态环境治理中的应用研究进展. 中国水土保持, 2012, (1): 41-44.
[7] 邵玺文, 阮长春, 赵兰坡, 等. 分蘖期水分胁迫对水稻生长发育及产量的影响. 吉林农业大学学报, 2005, 27(1): 6-10.
[8] 杨贵羽, 罗远培, 李保国, 等. 不同土壤水分处理对冬小麦根冠生长的影响. 干旱地区农业研究, 2003, 21(3): 104-109.
[11] 李朝苏, 刘鹏, 蔡妙珍, 等. 荞麦对酸铝胁迫生理响应的研究. 水土保持学报, 2005, 19(3): 105-109.
[12] 徐炳成, 山仑. 苜蓿和沙打旺苗期需水及其根冠比. 草地学报, 2003, 11(1): 78-82.
[13] 赖运平, 李俊, 张泽全, 等. 小麦苗期抗旱相关形态指标的灰色关联度分析. 麦类作物学报, 2009, 29(6): 1055-1059.
[15] 江宏娟, 李建龙, 李良霞. 高温胁迫下水肥耦合对高羊茅生态性状的影响. 草业科学, 2009, 26(7): 152-157.
[16] 潘瑞炽, 王小菁, 李娘辉. 植物生理学[M]. 北京: 高等教育出版社, 2008.
[17] 曹慧, 王孝威, 韩振海, 等. 水分胁迫诱导平邑甜茶叶片衰老期间内肽酶与活性氧累积的关系. 中国农业科学, 2004, 37(2): 274-279.
[18] 钱永强, 孙振元, 韩蕾, 等. 野牛草叶片活性氧及其清除系统对水分胁迫的响应. 生态学报, 2010, 30(7): 1920-1926.
[19] 夏阳. 水分逆境对果树脯氨酸和叶绿素含量变化的影响. 甘肃农业大学学报, 1993, 28(1): 26-31.
[20] 王瑞, 梁坤伦, 周志宇, 等. 不同淹水梯度对紫穗槐的营养生长和生理响应. 草业学报, 2012, 21(1): 149-155.
[24] 张明生, 谈锋. 水分胁迫下甘薯叶绿素a/b比值的变化及其与抗旱性的关系. 种子, 2001, (4): 23-25.
Effects of water and fertilizer on growth and photosynthetic pigments ofPueliasinense
ZHANG Ming-Jin1, CHEN Liang-Hua1, Hu Xiang-Wei1, Xu Rui1, ZHANG Jian1,2*
1.KeyLaboratoryofEcologicalForestryEngineering,InstituteofEcology&Forestry,SichuanAgriculturalUniversity,Chengdu611130,China; 2.CollegeofForestry,SichuanAgriculturalUniversity,Ya’an625014,China
A pot experiment was conducted to detect the interaction effect of water and fertilizer on growth and photosynthetic pigments ofPueliasinense. The experiment was a completely randomized design with 9 factorial combinations of 3 levels of water [drought, 25% field capacity (FC); control, 75% FC; flooding, 100% FC)] and fertilizer (low, moderate, high levels of fertilizer). Above-ground biomass, below-ground biomass, total biomass, and root: shoot ratio were significantly affected by the interaction of water and fertilizer. For the control water treatment, a high level of fertilization led to increased tiller number and plant height, and greater below-ground, above-ground, and total plant biomass. This combination was the best among the 9 combinations. These results indicate that normal supply of water and high nutrient supply are the optimal conditions to achieve a high yield for this species. Both flooding and drought were detrimental to tillering and growth. However, under water deficit where a tillering reduction was observed, tillering capacity could be improved through fertilization. Under flooding, high fertilization was not beneficial and a moderate amount of fertilizer was sufficient for optimal accumulation of below-ground biomass. Plants in these conditions had a higher root to shoot ratio, possibly a plant response to hypoxic conditions. Compared to flooding, drought was more detrimental to the accumulation of above-ground biomass. In order to adapt to drought,P.sinenseallocated more assimilate to the belowground organs. The increased root: shoot ratio was presumably advantageous for plant water capture and so to adaptation to drought. Interestingly, under water deficit stress, concentrations of photosynthetic pigments were significantly higher than those under control conditions. With increase in fertilization, contents of photosynthetic pigments also increased, especially when exposed to drought at the same time. These results elucidated some morphological and physiological adaptive mechanisms of this species when responding to water-deficit. Increased fertilization could mitigate the detrimental effects of water deficit stress on plant growth to some degree.
Pueliasinese; water and fertilizer coupling; growth; photosynthetic pigments
10.11686/cyxb20150509
http://cyxb.lzu.edu.cn
2014-04-21;改回日期:2014-06-10
国家科技支撑计划课题(2011BAC09B05)资助。
张明锦(1990-),女,四川芦山人,在读硕士。E-mail:958192283@qq.com *通讯作者Corresponding author. E-mail:sicauzhangjian@163.com
张明锦,陈良华,胡相伟,徐睿,张健. 水肥耦合对巨能草生长和光合色素的影响. 草业学报, 2015, 24(5): 75-83.
Zhang M J, Chen L H, Hu X W, Xu R, Zhang J. Effects of water and fertilizer on growth and photosynthetic pigments ofPueliasinense. Acta Prataculturae Sinica, 2015, 24(5): 75-83.
猜你喜欢
汽车实用技术(2022年16期)2022-09-03
现代农村科技(2022年9期)2022-08-16
林业科技(2020年3期)2021-01-21
农民致富之友(2019年4期)2019-03-13
中国科技纵横(2018年14期)2018-08-30
浙江农林大学学报(2018年4期)2018-08-22
小哥白尼(军事科学)(2018年3期)2018-06-15
现代农业科技(2017年16期)2017-09-22
江苏农业科学(2014年1期)2014-07-18
作物研究(2014年6期)2014-03-01
