
磷水平对不同基因型甘蔗苗期根系性状的影响
2015-04-13 04:09郑超李奇伟黄振瑞敖俊华刘可星
热带农业科学 2015年2期
郑超+李奇伟+黄振瑞+敖俊华+刘可星
摘 要 采用盆栽砂培方法研究4个不同磷水平(0、0.01、0.1、1 mol/L)对粤糖93-159(YT93-159)、新台糖22号(ROC22)和粤糖00-236(YT00-236)甘蔗苗期根系性状的影响。结果表明:随着磷水平的提高,甘蔗生物量增加,地上部生长迅速,根冠比显著降低;低磷胁迫能促进根系生长,在一定磷浓度范围内,形态指标值随着磷水平的升高而提高;高磷(1 mmol/L)会抑制YT93-159和YT00-236根系的生长。根系活力和ATPase活性均随施磷量的增加有上升的趋势,根系活跃吸收面积始终保持在49%左右。YT93-159在低磷胁迫下,根系生长最发达,不同磷水平引起的根冠比变化值比ROC22、YT00-236小;YT93-159的根重、地上部重、根系活力、ATPase活性等均最大,ROC22次之,YT00-236最小。
关键词 磷素水平 ;甘蔗 ;基因型 ;根系性状
分类号 S566.1
磷是植物生长发育所必需的大量营养元素,它不仅是植物体的组成成分,也是植物体内能量载体的主要组分和提供者,在植物的光合作用、呼吸作用、糖分代谢、酶促反应和生理生化调节过程中起着至关重要的作用[1]。由于磷容易被土壤固定,且扩散系数很小,因此土壤磷的作物有效性很大程度上取决于作物根系的生物学特性,如根长、根半径、根系比表面积、根构型及根分泌物等[2-3]。根系是构成植物体的主要部分,也是植物与外界环境进行物质和能量交换的场所,其分布特征反映了植物对环境的利用程度。植物根系可通过一系列的形态、生理变化主动适应低磷胁迫并改善自身的磷素营养状况。在缺磷逆境中,植物通常为寻求磷源,根系发育增强、伸长,侧根的长度和密度都增加,以加强对磷的吸收[4-5]。在玉米、小麦、大豆等作物中已发现植物可在磷胁迫下通过对根毛的分化、侧根与不定根的发生、根冠比增加与根轴增长等方式改变自身根系形态和根系构型来提高对低磷环境的适应[6-7]。
以往对甘蔗的研究主要集中在磷对甘蔗叶的营养、生理、产量、品质和土壤养分等方面,而有关施用磷肥对甘蔗根系性状的影响鲜有报道。鉴于此,本试验研究不同磷水平对不同基因型甘蔗的根系形态特征、理化特性的影响,以期为甘蔗合理施磷提供理论依据,同时探讨磷对甘蔗根系生长发育的影响及其重要性,为磷高效基因型甘蔗的筛选和遗传改良提供科学依据。
1 材料与方法
1.1 材料
供试甘蔗品种组培苗:新台糖22号、粤糖00-236和粤糖93-159(以下简称ROC22、YT00-236、YT93-159)。
供试基质:粒径为0.6~2 mm的粗细适宜的河砂,用清水淘洗去除泥土和粉砂,然后用盐酸洗涤(使试验过程中不从砂中向培养液中溶解出盐类且不吸附盐类),用自来水清洗至中性,最后用去离子水清洗2次。
营养液配制:依据甘蔗对N、P、K的吸收特性对Hoagland 营养液配方进行适当改良。KNO3 252 mg/L,Ca(NO3)2·4H2O 506 mg/L,MgSO4 246 mg/L,ZnSO4·7H2O 0.21 mg/L,CuSO4·5H2O 0.1 mg/L,H3BO3 2.86 mg/L,(NH4)6Mo7O24·4H2O 0.01 mg/L,MnCl2·4H2O 1.81 mg/L。铁盐溶液(需单独配制):FeSO4·7H2O 7.95 g,EDTA·Na 9.3 g,蒸馏水500 mL,稀释1 000倍。KH2PO4 139 mg/L(单独配制),分别稀释1、10、100倍。
1.2 方法
1.2.1 试验设计
试验于2009年8月26日在广东海洋大学兴农楼5楼玻璃温室进行。试验设置3个甘蔗品种(ROC22、YT00-236、YT93-159),4个磷水平(0、0.01、0.1、1 mol/L,以下简称P0、P1、P2、P3),共12个处理,每个处理重复4次,总共96桶。每桶装砂1.5 kg,砂桶底部留有小孔,浸入盛装营养液的桶5 cm深。选用20 cm左右、长势均匀的甘蔗组培苗移植(移苗应在10:00 a.m.前或17:00 p.m.后进行),每桶移栽2株。试验均按照温室试验管理,每天按时补充去离子水至营养液为种植时的水平,防止由于水分减少而导致营养液的养分浓度改变,调节营养液的pH值至6.0,每周更换1~2次营养液。在移栽30~40 d后,收获并测定各个根系的性状。
1.2.2 测定项目及方法
(1)植株生物量的测定:采用烘干测重法,首先将地上部与根部分别称取鲜重,然后在105℃下杀青,而后在60℃下烘干至恒重,再分别称重。
(2)根系形态的测定方法:将植株根系鲜样充分清洗整理后,运用数字化扫描仪将根系图像扫描存入电脑,再利用根系图像分析软件对根图像进行分析,最后获取根长、根尖数、根体积等根系形态参数。
(3)根系活力的测定:分别将侧根根尖的根毛区切断,称取不同处理相同部位侧根根毛区0.5 g,具体测定方法采用氯化三苯基四氮唑(TTC)法。以μg TTF/(g FW·h)为单位表示活力大小。
(4)根系总吸收面积和活跃吸收面积的测定:用量筒排水法分别量取不同处理相同部位侧根根毛区鲜样1.0 mL,具体测定方法采用甲烯蓝法。
(5)根系ATPase活性的测定:参照Chen等[8]的方法。
1.3 数据处理
采用Microsoft Excel 2003和SPSS(V17.0)软件对数据进行统计分析,利用Duncan法进行多重比较,显著性水平α=0.05。
2 结果与分析
2.1 磷水平对甘蔗生物量及根冠比的影响
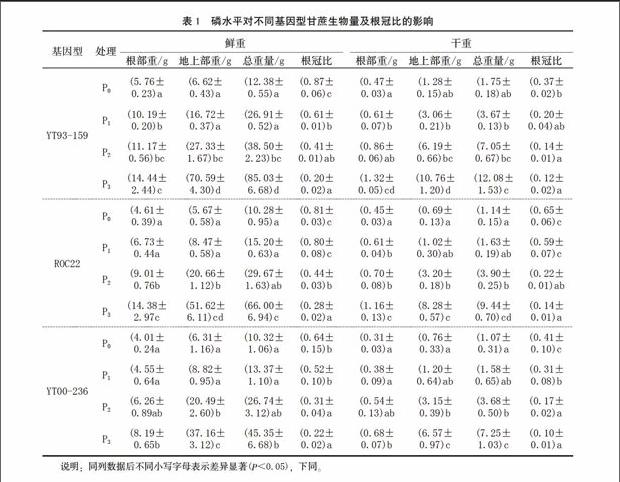
由表1可见,施磷使甘蔗生物量明显增加,根部重和地上部重均与磷浓度的变化呈正相关。随着磷浓度的增加,生物量呈上升的平滑曲线变化,YT93-159的增加量明显最高,ROC22次之,YT00-236最低。说明施磷对甘蔗生物量的积累有显著的促进作用。YT93-159的生物量既与品种遗传特性有关,也与其对磷的利用效率有关。
根冠比的变化被认为是植物对低磷的耐受机制之一,是植物对低磷胁迫适应性反应和耐低磷能力的标志[6]。中国对小麦、油菜、玉米、水稻等的研究结果也表明,缺磷时,不同作物品种间的地上部分干重变化很大,磷高效品种的根冠比远大于磷低效品种[6,9-10]。从P0处理到P1处理,YT93-159、ROC22、 YT00-236的鲜重根冠比依次下降26%、1%、12%,干重根冠比依次下降17%、6%、10%。初步说明,YT93-159在低磷环境下仍能保持正常的生长速度,属于耐低磷品种。从P0处理到P3处理,YT93-159、ROC22、YT00-236的鲜重根冠比依次下降67%、53%、42%,干重根冠比依次下降25%、51%、31%。随着施磷量的增加,各处理品种的根冠比呈下降趋势。这说明光合产物在分配方向和强度上随磷素水平的改变而变化,高磷供应使光合产物分配到地上部的比例增大,以促进茎叶的生长发育,提高作物产量;缺磷或低磷胁迫使光合产物分配到地下部的比例增大,以促使根系发达,扩大与土壤的接触面积,从而提高根际养分有效性。而YT93-159的生长速度比较快。在低磷环境下,YT93-159的生物量最大,干重根冠比最小。由此可见,YT93-159对磷素反应敏感,能有效利用土壤中的磷素增加根重。
2.2 磷水平对甘蔗根系形态的影响
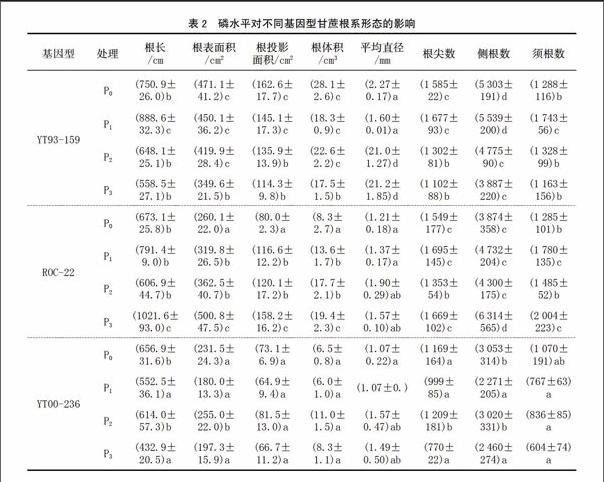
由表2可知,YT93-159在低磷条件下,根系生长得更好;ROC22和 YT00-236则是随着磷水平的提高,根系生长得更好。但超过一定的磷浓度,根系的生长则受到抑制,尤其在YT93-159上表现的更加明显。总之,施磷可促进根系的生长。
植物一般仅能吸收距根表面1~4 mm根际土壤中的磷[11]。植物受低磷胁迫时,根系发育加快,以缩短磷离子扩散到植物根的距离和扩大根系的吸收面积。植物适应低磷胁迫的一个重要的反应就是改变根系的生长及构型[12]。本试验中,在P0和P1处理下,各基因型品种的根系各项形态指标值明显增加,尤其YT93-159在各处理中表现得更加明显。在P1处理下,YT93-159除根尖数和须根数仅次于ROC22外,其余数值均比相同磷水平的ROC22和 YT00-236高,并且在YT93-159的所有磷水平处理中排在第1位,可见,低磷胁迫下,YT93-159的根系生长反而更加好,说明YT93-159可能是耐低磷品种,低磷胁迫使其根系发达,以吸收更多的养分来维持正常的生长。因此,YT93-159耐低磷能力最强,ROC22次之,YT00-236最低。因为根直径与磷的吸收效率呈反比,在根重相同情况下,根越细,磷的吸收效率越高[13];而其他根系性状如根长、侧根数、须根数等均与磷吸收效率呈正比,表明低磷促进YT93-159根系的生长,高磷则抑制根系的生长。
2.3 磷水平对甘蔗根系活力的影响
2.3.1 根系TTC还原量的差异
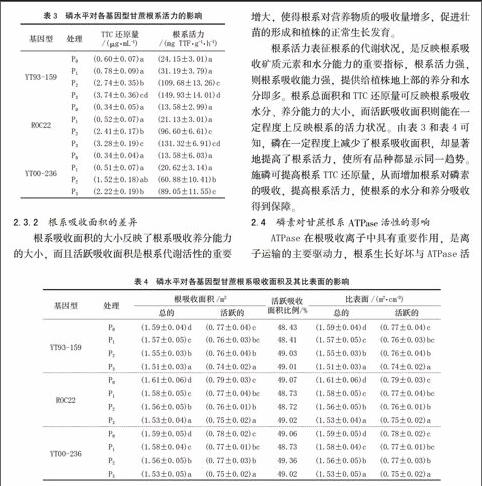
植物根系是活跃的吸收器官和合成器官,根的生长状况和活力水平直接影响地上部生长、营养状况及产量水平。根系活力大小一定程度上反映了作物吸收养分能力的强弱,一般情况下,根系活力越高,吸收养分的能力越强。根系TTC还原量通常作为衡量根系活力大小的有效指标。由表3可知,各基因型的TTC还原量均随着磷水平的提高而增加,根系活力也呈上升趋势,并且在P3达到最大值,说明磷可提高根系活力。此外,YT93-159在各个磷水平中根系活力最大,尤其在低磷条件下表现得更加明显。ROC22和 YT00-236在P0与P1处理时,差异并不明显,而在P2与P3处理时,ROC22的TTC还原量与根系活力明显高于YT00-236。
2.3.2 根系吸收面积的差异
根系吸收面积的大小反映了根系吸收养分能力的大小,而且活跃吸收面积是根系代谢活性的重要参数。由表4可知,各基因型甘蔗根系吸收面积随磷浓度的升高有一定的下降趋势,且呈显著性差异。不同基因型甘蔗在相同磷水平处理下根系吸收面积无显著性差异。以所有处理的根体积为1 cm3计算,从P0处理到P3处理,YT93-159、ROC22、YT00-236的根系总吸收面积依次降低为5.04%、4.79%、3.33%,根系活跃吸收面积依次降低为3.84%、4.73%、3.25%。这表明无磷或低磷供应能在一定程度上提高根系的吸收面积,此结果与前述低磷对根系性状的影响一致。由于根系吸收面积的增大,使得根系对营养物质的吸收量增多,促进壮苗的形成和植株的正常生长发育。
根系活力表征根系的代谢状况,是反映根系吸收矿质元素和水分能力的重要指标,根系活力强,则根系吸收能力强,提供给植株地上部的养分和水分即多。根系总面积和TTC还原量可反映根系吸收水分、养分能力的大小,而活跃吸收面积则能在一定程度上反映根系的活力状况。由表3和表4可知,磷在一定程度上减少了根系吸收面积,却显著地提高了根系活力,使所有品种都显示同一趋势。施磷可提高根系TTC还原量,从而增加根系对磷素的吸收,提高根系活力,使根系的水分和养分吸收得到保障。
2.4 磷素对甘蔗根系ATPase活性的影响
ATPase在根吸收离子中具有重要作用,是离子运输的主要驱动力,根系生长好坏与ATPase活性高低密切相关[14],ATPase活性的增强是根系代谢旺盛的重要标志。本试验结果表明,磷对根系ATPase活性产生显著的影响。由表5可见,从P0到P3,YT93-159、ROC22、 YT00-236的ATPase活性依次提高了4.11、3.12、3.47倍。P0与P1之间ATPase活性差异并不显著,到P2时出现显著差异,P3时出现极显著差异。所有的根系ATPase活性均表现出随供磷水平的提高而增强的趋势,说明供磷能提高根系的吸收能力和能量代谢的水平,磷对ATPase活性具有重要的促进作用。此外,在不同磷水平处理下,YT93-159的ATPase活性最高,ROC22次之,YT00-236最低。因此,YT93-159根系对养分的吸收较强,具有较强的生理活动能力,根系新陈代谢也较为旺盛。
3 讨论与结论
根系是作物吸收水分和养分的重要器官,也是较先感受并传导养分胁迫信号的器官,尤其对磷的亏缺反应敏感。磷胁迫条件下,作物地上部的光合产物向根部转移量相对增加,使根冠比增大[15]。杨瑞吉等[16]的研究结果发现,不同基因型小麦开花期的根干重均随施磷量的增加而增加,磷胁迫下根数量增加,根系变长,根半径减小,以适应低磷逆境。在本研究中,甘蔗也会通过改变其根构型与各种根系性状等来适应低磷胁迫,各基因型甘蔗在P0和P1(0.01 mol/L)处理下,根长、根表面积、根投影、根体积、根尖数、侧根数、根尖数均明显增加,根平均直径减小,地上部生长受抑制,根冠比显著提高。随着供磷水平的提高,只有ROC22的各根系指标值相应地增大,YT00-236和YT93-159的根系生长却受到不同程度的抑制。这与王建霄等[17]的研究结果一致,即随着磷浓度的增加,橡胶树幼苗1~4级根的平均长度、数量和总长度随着供磷浓度的增加,呈先上升后下降的趋势。
高磷供应下(1 mmol/L),各基因型甘蔗的干物质积累都达到最大值,积累量大小为YT93-159>ROC22>YT00-236。这与曹爱琴等[18]研究结果不同基因型菜豆在高磷处理条件下其生物量和吸磷量均高于低磷处理的观点相类似。此外,高磷在一定程度上抑制根系的生长,从而使光合产物向地上部运输,降低了根冠比。同时,TTC还原量明显增大,根系活力和ATPase活性明显增强。
刘灵等[19]发现,在低磷处理下供试大豆基因型间生物量和产量具有极显著的基因型差异,且根形态、构型与磷效率密切相关。另有研究为表明,植株根冠比、根长的增加被认为是植物适应低磷胁迫的特征,在低磷条件下,根冠比、根长增加显著的物种或基因型具有较好的低磷适应性[20]。与正常供磷相比,低磷处理条件下山核桃根长、根表面积和根体积显著降低[21]。本试验结果表明,YT93-159在低磷胁迫下,根平均直径最小,而根长、根尖数、侧根数、根表面积等都达到最大值,并且根系生长明显发达,由此推断,YT93-159与ROC22、YT00-236相比,苗期相对耐低磷。由于在各个磷水平处理下,YT93-159的生物量、根长、根体积、平均直径、根系活力和ATPase活性等均高于ROC22与 YT00-236,YT93-159的磷利用效率最高,ROC22次之,YT00-236最低。笔者还发现,在各个磷水平下,植物根系的活跃吸收面积无显著差异,始终保持在49%左右,这说明一定范围的根系活跃吸收面积是甘蔗生长的必须保障。
综上所述,低磷胁迫在一定程度上促进了甘蔗根系的生长,适当供磷,促进地上部生长,降低根冠比,有利于生物量的积累,增强根系活力,使得根系代谢增强,因此磷对甘蔗的生长至关重要。目前,磷在土壤中的难溶性和难移动性决定了围绕提高根际磷的溶解性、扩展根系的吸收范围的研究始终是工作的核心。磷效率的广泛变异性给人们以启示,培育磷高效的作物品种具有巨大的潜力[22]。
参考文献
[1] Abelson P H. A potential phosphate crisis[J]. Science, 1999, 283(11): 2 015-2 021.
[2] Gardner W K, Barber D A. The acquisition of phosphorus by Lupinus albus L. The probable mechanism by which phosphorus movement in the soil root interface is enhanced[J]. Plant and Soil, 1983, 70(1): 107-124.
[3] Ae N, Arihara J, Okada K, et al. Phosphorus uptake by pigeon pea and its role in cropping systems of the Indian Subcontinent[J]. Science(Washington D C), 1990, 248(3): 477-480.
[4] Burton H R, Kasperbauer M J. Changes in chemical composition of tobacco laminar during senescence and curing 1, plastid pigments[J]. Agriculture and Food Chemistry, 1985, 33(5): 879-883.
[5] 秦 松,刘大翠,刘 静,等. 土壤肥力对烟叶化学成分及品质的影响[J]. 土壤通报,2007,38(5):901-905.
[6] 马样庆,梁 霞. 植物高效利用磷机制的研究进展[J]. 应用生态学报,2004,15(4):712-716.
[7] 何 蓉,程雪斌,胡秋芬,等. 云南12种豆科灌木饲料中主要磷形态分析[J]. 云南大学学报:自然科学版,2003,25(6):528-530.
[8] Chen J C, Fu W H. Determine for activity of membrane cooped ATPase in leaf cell[J]. Chinese Journal of Cell Biology, 1983, 5(3): 21-24.
[9] 孙海国,张福锁. 缺磷胁迫下的小麦根系形态特征研究[J]. 应用生态学报,2002,13(3):295-299.
[10] 郭玉春,林文雄,石秋梅,等. 低磷胁迫下不同磷效率水稻苗期根系的生理适应性研究[J]. 应用生态学报,2003,14(1):61-65.
[11] 李庆逵. 现代磷肥的研究进展[J]. 土壤学进展,1986,2(1):1-7.
[12] Lynch J P. Root architecture and plant productivity[J]. Plant Physiology, 1985, 109(1): 7-13.
[13] 王 毅. 植物耐低磷胁迫遗传学研究策略[J]. 热带农业科学,2004,24(4):34-40.
[14] 朱 林,张春兰,沈其荣. 施用稻草等有机物料对连作黄瓜根系活力、硝酸还原酶、ATP酶活力的影响[J]. 中国农学报,2002,18(1):17-19.
[15] 刘国顺,肖庆礼,王艳丽. 不同供磷能力的土壤施磷对烤烟根体积和根冠比以及根系伤流组分的影响[J]. 中国烟草学报,2009,15(2):28-32.
[16] 杨瑞吉,张小红,王鹤龄,等. 不同基因型春小麦对磷胁迫适应性研究[J]. 西北植物学报,2005,25(11):2 314-2 318.
[17] 王建霄,罗 微,茶正早,等. 水培条件下不同磷水平对橡胶树幼苗根系生长发育的影响[J]. 海南大学学报,2009,27(3):265-269
[18] 曹爱琴,廖 红,严小龙. 低磷土壤条件下菜豆根构型的适应性变化与磷效率[J]. 土壤学报,2002,39(2):276-281.
[19] 刘 灵,廖 红,王秀荣,等. 不同根构型大豆对低磷的适应性变化及其与磷效率的关系[J]. 中国农业科学,2008,41(4):1 089-1 099.
[20] 曹一平. 高等植物的矿质营养[M]. 北京:北京农业大学出版社,1988:137-144.
[21] 李永夫,金松恒,叶正钱,等. 低磷胁迫对山核桃幼苗根系形态和生理特征的影响[J]. 浙江林学院学报,2010,27(2):239-245.
[22] 樊明寿,张福锁. 植物磷吸收效率的生理基础[J]. 生命科学,2001,13(3):129-131.
猜你喜欢
安徽农学通报(2022年6期)2022-04-07
小猕猴学习画刊(2019年8期)2019-09-16
广东第二课堂·小学(2019年1期)2019-03-06
新课程·下旬(2018年8期)2018-11-10
作文·初中版(2018年4期)2018-05-22
广东第二课堂·初中(2018年4期)2018-05-08
试题与研究·教学论坛(2017年28期)2017-11-29
吉林农业·下半月(2016年11期)2017-01-09
中学生理科应试(2016年4期)2016-11-19
作文大王·低年级(2016年9期)2016-09-21
