
东海甲藻硅藻现场生长营养盐相关参数的计算
2015-04-11 03:26周超凡张传松韩秀荣石晓勇李雁宾
海洋科学 2015年10期
周超凡, 张传松, 韩秀荣, 石晓勇, , 刘 强, 李雁宾,
(1.中国海洋大学 化学化工学院, 山东 青岛 266100; 2.中国海洋大学 海洋化学理论与工程技术教育部重点实验室/海洋科学与技术青岛协同创新中心, 山东 青岛 266100; 3.国家海洋局 海洋减灾中心, 北京100194)
水体富营养化是目前全球面临的主要海洋环境问题之一。长江作为我国第一大河流, 每年输送到东海大量的营养物质, 导致东海近岸海域富营养化严重, 赤潮频发。近年来海洋局监测结果表明, 东海赤潮问题位居中国四大海区之首[1], 2005年东海赤潮发生频次占四大海域总赤潮发生次数的 63.3%。东海赤潮肇事种也发生了明显的演替现象, 由硅藻赤潮为主演变成近年的甲藻赤潮占优[2-3]。2002年以来几乎每年春季都会爆发大规模的甲藻赤潮, 如2005年5月到6月东海发生了面积超过7 000 km2的米氏凯伦藻赤潮[4-5]。营养盐是赤潮形成的必要条件, 其浓度和比例都会对浮游植物的生长产生显著影响。营养盐半饱和常数以及浮游植物细胞内的营养盐比例(氮磷比和硅氮比)是研究营养盐循环和营养盐对浮游植物生长影响的重要参数,能够指示出不同藻类对营养盐的亲和性, 揭示营养盐在浮游植物生长和演替中的作用[6-7]。此外, 这几种营养盐相关参数也广泛应用于生态动力学模型中[8-9], 是模拟浮游植物吸收营养盐过程和营养盐限制浮游植物生长过程的高灵敏度参数, 其取值是否合理往往会决定模型模拟浮游植物生长过程的准确性[10-11]。
营养盐培养实验是获取浮游植物半饱和常数的主要方法。营养盐培养实验可分为实验室培养实验和现场培养实验。实验室培养实验的优点在于实验条件可控, 易于设置较密营养盐梯度。但由于实验室条件(包括培养条件及藻种生理生化状态)与实际环境差距较大, 实验室培养实验得到的营养盐相关参数与实际海域出入较大。相对于实验室培养实验, 现场培养实验更接近于海域自然条件, 但受现场实验条件限制, 现场培养实验时往往难以设置较密营养盐梯度, 难以通过实验数据直接计算得到半饱和常数等营养盐相关参数。本研究基于生态动力学模型简化法, 建立了现场船基培养实验营养盐半饱和常数和浮游植物胞内营养盐比例计算方法, 并获得了东海硅藻和甲藻的营养盐吸收半饱和常数, 为东海赤潮生态动力学模型的构建和营养盐迁移—转化动力学机制研究提供了基础参数。
1 数据与方法
1.1 数据来源
本文采用数据为 2003~2005年东海多批次船基培养实验结果, 包括: 2003年 5月份船基围隔实验,(简称数据1)[12]、2003年5月份现场船基培养实验(简称数据2)[13]、2005年3月底到4月初现场船基培养实验[14](简称数据3)、2005年5月底到6月初现场船基培养实验(简称数据4)[15]。培养站位如图1所示。
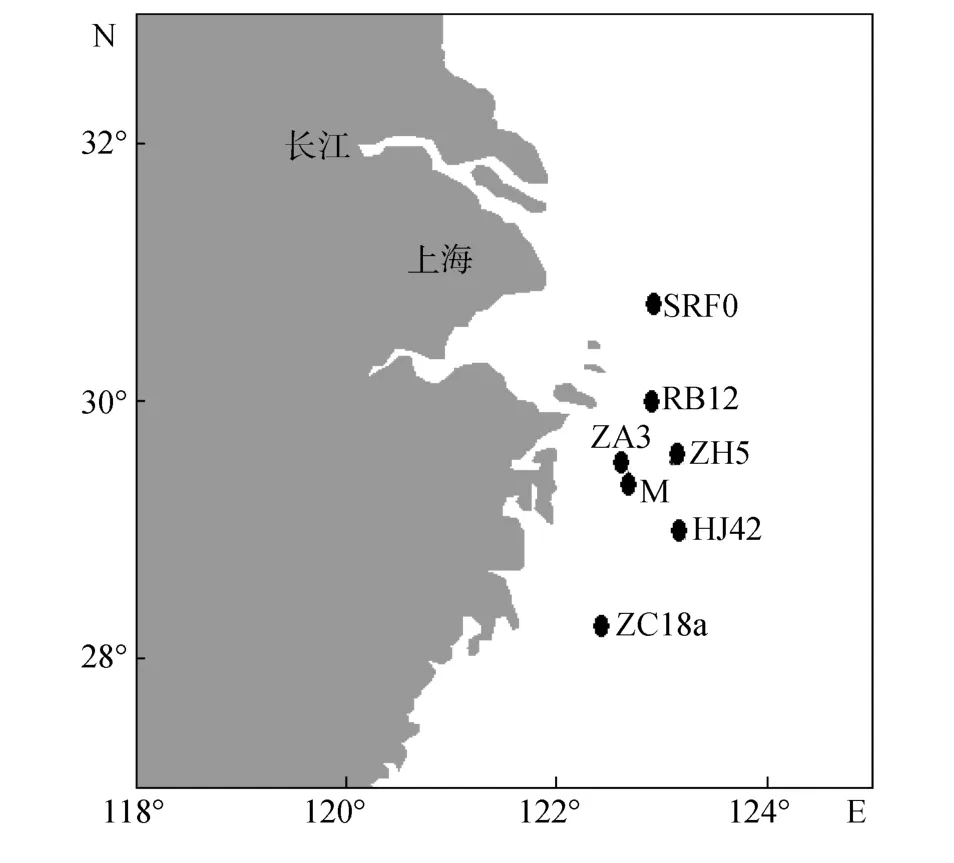
图1 现场培养站位示意图Fig.1 A map showing the sites of field incubation experiments
现场船基培养实验周期为7~8 d, 详细方法见文献[12-15]。表层海水经200 μm的筛网过滤去除浮游动物后, 添加一定量硝酸盐、磷酸盐、硅酸盐, 混匀后装入聚碳酸酯培养瓶中, 在海水流水浴中培养, 每天摇瓶 3~4次保持溶解气体量并防止生物聚集。现场培养实验测定六项参数: SiO3-Si、PO4-P、NO3-N、NO2-N、NH4-N、颗粒氮(PN)。硅酸盐、磷酸盐、硝酸盐分别采用硅钼蓝分光光度法、磷钼蓝分光光度法、铜镉还原法测定。PN采用碱性硫酸钾氧化法测定。
1.2 数据处理方法
营养盐等变量随时间变化图采用Origin 8.0软件绘制。简化生态动力学模型在Modelmaker 4.0软件环境下开发。采用Matlab软件编程计算模拟结果和实测结果的相关系数(R2)、置信度区间(P)和相对标准偏差(RSD)。
2 结果与讨论
2.1 生态动力学模型简化法计算现场培养实验营养盐相关参数方法建立
如方程 1 所示, 生态动力学模型中浮游植物动力学过程主要包括浮游植物光合作用(Photo)、呼吸作用(Resp)、体外分泌(Exud)、自然死亡(Mort)和被浮游动物捕食(Graz)等过程。
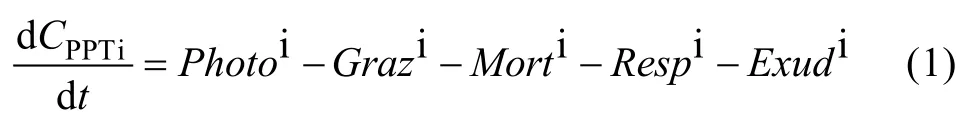
其中, 光合作用过程(Photo)需要考虑光照、温度、营养盐的限制作用, 可用方程 2表示。其中, 采用Michaelis-Menten方程表示营养盐对浮游植物生长的限制作用[16], 采用乘法法则[17]处理多营养盐和多环境因子的综合作用。由于甲藻的生长不需SiO3-Si,对于甲藻只考虑 DIN和 PO4-P的限制作用, 硅藻则考虑DIN、PO4-P、SiO3-Si的限制作用。
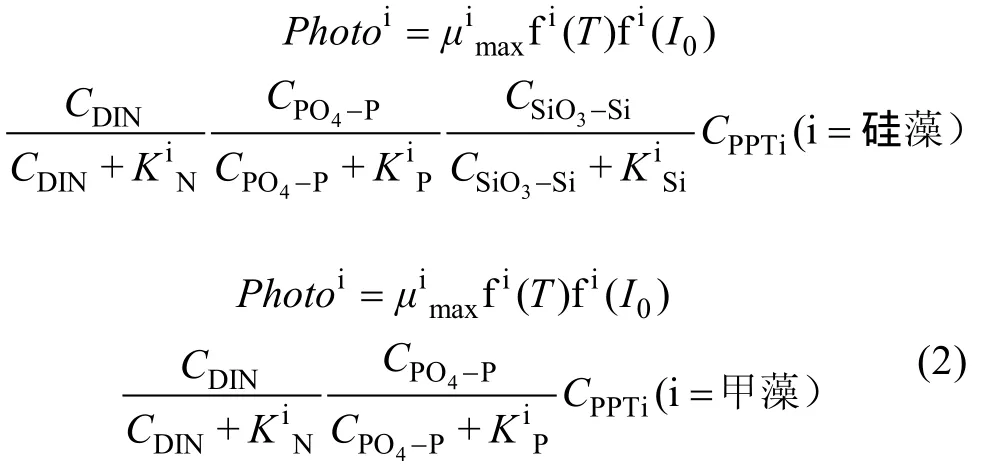
其中,CPPTi为浮游植物生物量, 以氮(N)表示;μimax为最大生长速率常数; fi(T)为浮游植物生长温度限制因子; fi(I0)为浮游植物生长光照限制因子;CDIN为 DIN浓度;CPO4-P为 PO4-P浓度;CSiO3-Si为SiO3-Si浓度; i=1时代表甲藻, i=2时代表硅藻。
由方程 1和方程 2可见, 浮游植物生长动力学方程中包括过多参数, 很难通过参数优化方法求出所有参数, 需对模型进行进一步简化。方程简化基于如下几个假设: 假设 1, 呼吸作用可以作为光合作用的“可逆”过程, 其速率与浮游植物生长速率呈较好的正相关性[18], 分泌过程在模型中也处理为与光合作用成正比, 因此这两个过程可以隐含到光合作用过程中; 假设 2, 本文所选用培养实验已去除了大型浮游生物, 捕食作用较弱, 假设浮游动物摄食过程可忽略; 假设 3, 现场培养时间较短, 环境条件变化小, 因此假设死亡速率常数为一恒定值(α)。基于这三个假设, 方程1可以简化为方程3。
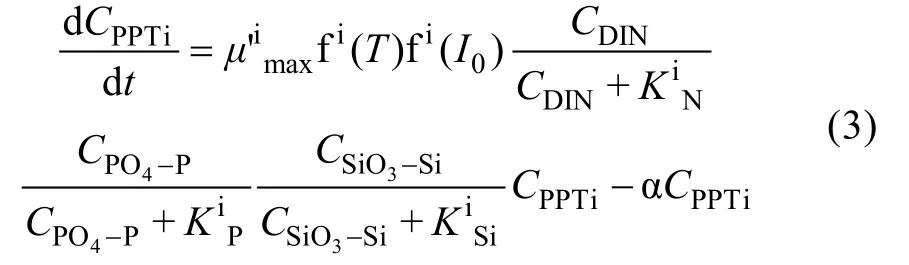
其中, α为浮游植物死亡速率常数;μ′imax为将分泌作用和呼吸作用扣除后的最大生长速率。
此外, 由于现场培养时间一般较短, 温度和光照波动较小, 温度和光照因子可以做常数处理, 这样假设培养期间温度限制因子和光照限制因子为常数, 将最大生长速率常数(μ′imax)和温度(fi(T))光照(fi(I0))限制因子合为一个参数(μ′i)后, 方程3可以简化为方程4。

最终的浮游植物生长方程简化为方程4, 相对应的 PN、DIN、PO4-P、SiO3-Si的微分方程则可由方程 5~8表示。简化后模型参数仅包括浮游植物生长速率常数、营养盐半饱和常数(N, P, Si)、氮磷比和硅氮比, 相关参数可以通过参数优化方法得到。模型在Modelmaker 4.0软件环境下运行, 微分方程数值积分采用 Runge-Kutta法, 模拟 DIN、PN、PO4-P、SiO3-Si随时间变化。采用马夸特(Marquardt)法对参数优化得到最优参数取值。为了减少计算过程中参数的数量, 保证优化结果的准确性, 选取培养实验数据时, 仅选取培养期间单一藻种(甲藻或硅藻)为优势种(优势度>80%)的培养数据, 参数优化时假设培养体系中仅含有一种浮游植物(优势种), 这样可以减少将近一半的参数。进一步采用现场培养实验数据应用该方法计算了东海甲藻和硅藻的现场半饱和常数及浮游植物营养盐比例。

其中,μ′i为生长速率常数;PPTNP、PPTSiN为浮游植物营养盐氮磷比、硅氮比; i=1时代表甲藻, i=2时代表硅藻。
2.2 营养盐等状态变量模拟结果
浮游植物生长过程吸收氮、磷、硅营养盐, 培养期间, DIN、PO4-P、SiO3-Si营养盐的浓度会随着时间而降低, PN浓度则随着浮游植物数量的增多而增加。以2005年6月SRF0站、RB12站为例, 将DIN、PN、PO4-P、SiO3-Si几个变量的实测值和模拟值比较, 见图2。由图可知, DIN、PN、PO4-P、SiO3-Si浓度的模拟结果和实验结果基本吻合, 较好地模拟了PN的增长趋势及DIN、PO4-P、SiO3-Si下降趋势。具体而言, 对于RB12站(图2的A-11-1~A-11-4), 颗粒态营养盐(PN)浓度随着时间呈 S型曲线升高; 溶解无机态营养盐(DIN、 PO4-P、SiO3-Si)随着浮游植物的吸收利用而被消耗, 随时间呈反S型曲线减少。对于SRF0站(图2的A-10-1~A-10-4), 溶解无机态营养盐随着浮游植物的吸收利用而被消耗, 基本在2~3 d降到最低值, 颗粒态营养盐的变化也在 2~3 d达到高值, 然后基本维持不变。进一步用Matlab软件计算了所有站位模拟值与实测值的相关系数(R2)、置信度(P)以及相对标准偏差(RSD), 统计结果见表1。模拟结果与实测值之间的R2较高, 80%以上R2都大于0.60, 且R2大于0.80的超过了70%。只有 A-10-1~A-10-4颗粒氮、A-11-4颗粒氮、A-5-1颗粒氮的P值大于 0.01,P值小于 0.01的超过了90%,P值小于0.05的超过了95%, 由相对标准偏差RSD计算也可以看出溶解态营养盐和PN模拟效果较好, 大部分RSD小于 20%。该模型能较好模拟研究海域浮游生态系统各状态量的变化, 表明所确定的模型参数基本能反映研究海域的地域化特征。
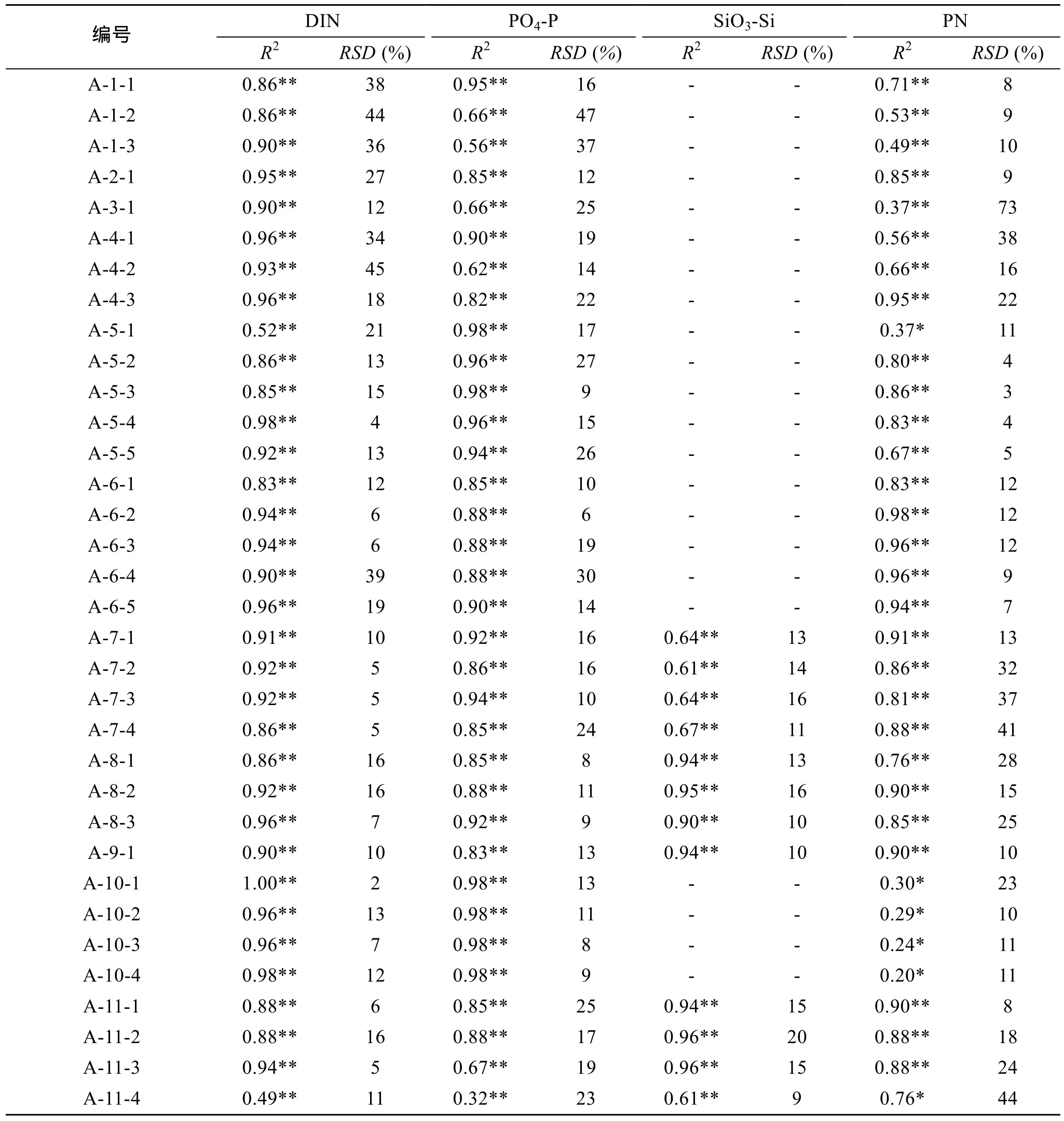
表1 所有站位模拟效果统计Tab.1 Statistical work of simulation results
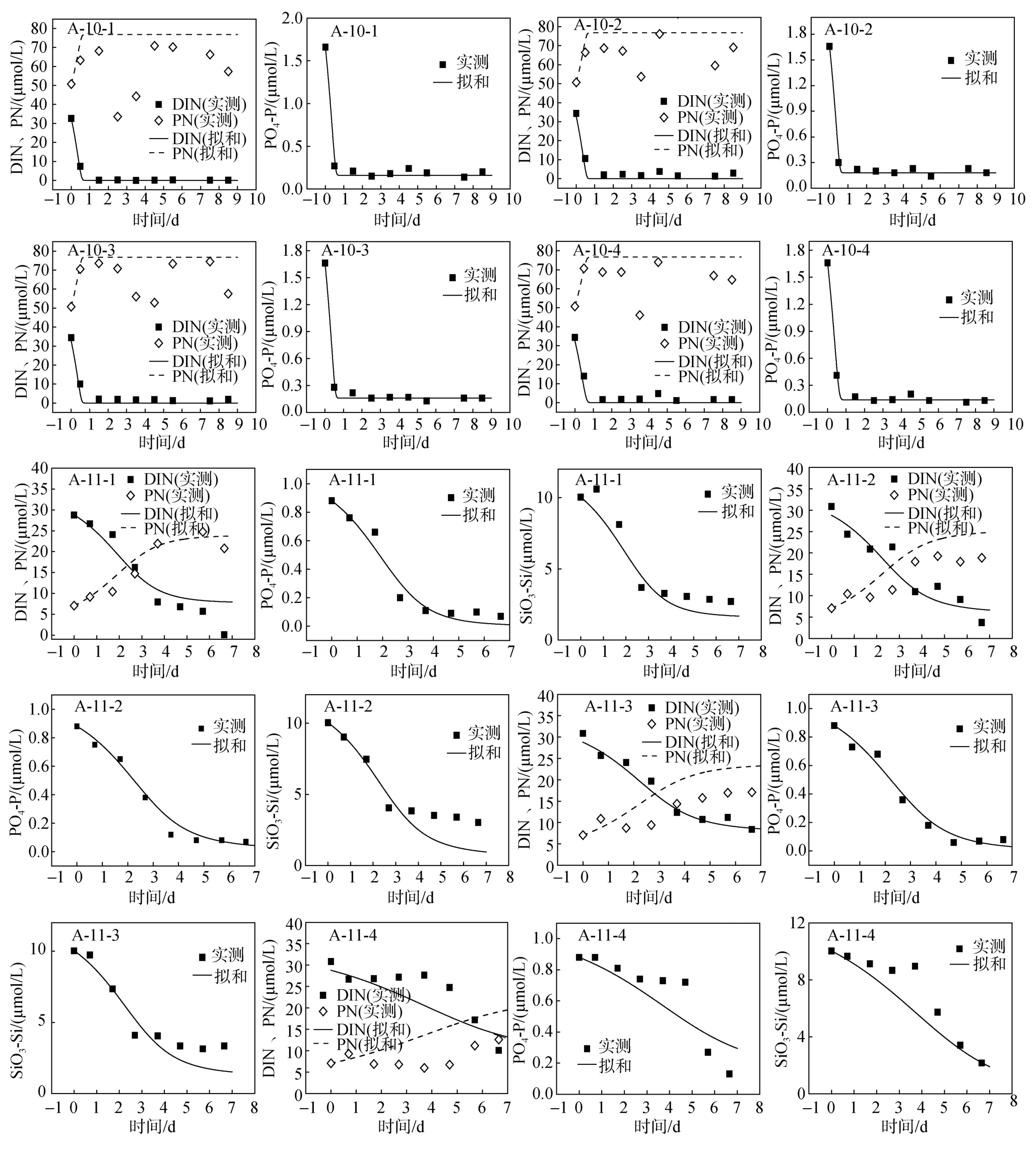
图2 2005年6月SRF0、RB12站培养实验营养盐浓度变化模拟值与实测值的比较Fig.2 The comparison of simulated and observed nutrition concentration at sites SRF0 and RB12 in June 2005
2.3 东海硅藻、甲藻营养盐相关参数计算结果
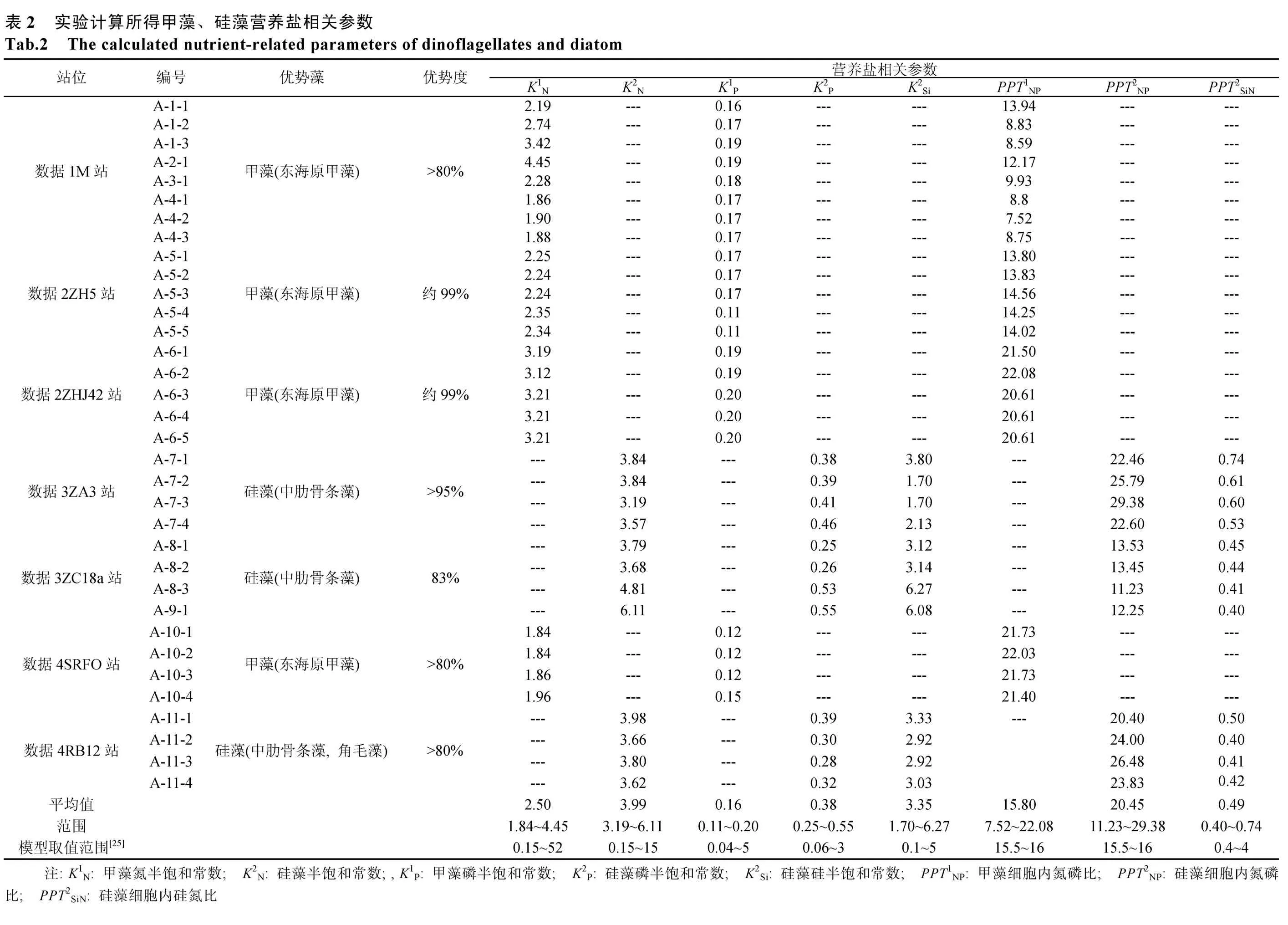
东海甲藻硅藻营养盐相关参数计算结果如表2所示。东海甲藻: 氮半饱和常数2.50 μmol /L、磷半饱和常数 0.16 μmol/L、营养盐氮磷比建议取值为15.80。东海硅藻: 氮半饱和常数3.99 μmol/ L、磷半饱和常数0.38 μmol/ L、硅半饱和常数3.35 μmol /L、营养盐氮磷比20.45、营养盐硅氮比建议取值为0.49。结果表明, 甲藻的无机氮半饱和常数以及磷酸盐半饱和常数要低于硅藻, 相对于甲藻而言, 硅藻更容易受到营养盐限制。总体而言, 本研究所得到的营养盐相关参数取值在文献报道的取值范围内。对于甲藻, 李英[19]等通过实验室培养方法, 研究了东海原甲藻与磷源营养盐的关系, 计算得到的甲藻磷酸盐半饱和常数K1P为 0.13 μmol/L, 赵晓玮[20]得出东海米氏凯伦藻对磷的半饱和常数为0.20 μmol/L, 李天深[21]等获得的亚历山大藻的磷半饱和常数为 0.75 μmol/L,与本研究所计算的甲藻磷酸盐半饱和常数 0.16 μmol/L
相近; 对于硅藻, 郑凌凌[22]测得的硅藻氮半饱和常数为 3.25 μmol/L, 与本研究计算的硅藻氮半饱和常数3.99 μmol/L基本一致, 张诚[23]等获得的尖刺拟菱形藻氮半饱和常数为2.18 μmol/L, 李铁[24]等通过实验室培养实验计算得出了中肋骨条藻和新月菱形藻的氮半饱和常数分别为7.8 μmol/L和6.0 μmol/L, 也与本文结果相近; 李铁[24]等测得的中肋骨条藻和新月菱形藻的磷半饱和常数分别为0.61 μmol/L和0.48 μmol/L, 与本研究计算得到的硅藻磷酸盐半饱和常数0.38 μmol/L基本一致。
3 结论
建立了生态动力学模型简化法和现场培养实验结合计算现场营养盐相关参数的方法, 并通过处理东海现场培养实验数据, 计算了东海甲藻、硅藻营养盐半饱和参数以及营养盐氮磷比、硅氮比。将优化得到的参数应用于生态动力学模型中, 能够较好模拟颗粒氮、溶解无机氮、磷酸盐、硅酸盐的时间变化规律, 80%以上R2都大于0.6, 且R2大于0.8的超过了70%,P值小于0.01的超过了90%,P值小于0.05的超过 95%, 表明模拟结果与实验结果吻合较好,也说明优化得到的模型参数能基本反映所研究东海海域的地域化特征, 优化得到的东海甲藻: 氮半饱和常数 1.84~4.45 μmol/L、磷半饱和常数 0.11~0.20 μmol/L、
营养盐氮磷比建议取值范围为 7.52~22.08; 东海硅藻: 氮半饱和常数3.19~6.11 μmol/L、磷半饱和常数0.25~0.55 μmol/L、硅半饱和常数, 1.70~6.27 μmol/L,营养盐氮磷比 11.23~29.38、营养盐硅氮比为 0.40~0.74, 。东海甲藻氮半饱和常数、磷半饱和常数、营养盐氮磷比建议取值分别为2.50 μmol/L, 0.16 μmol/L,15.80; 东海硅藻氮半饱和常数、磷半饱和常数、硅半饱和常数、营养盐氮磷比、营养盐硅氮比建议取值分别为 3.99 μmol/L, 0.38 μmol/L, 3.35 μmol/L,20.45, 0.49。
?
[1] 国家海洋局.中国海洋环境质量公报[R].北京: 国家海洋局, 2001-2013.
[2] Zhou M J, Shen Z L, Yu R C.Responses of a coastal phytoplankton community to increased nutrient input from the Changjiang (Yangtze) River[J].Continental Shelf Research, 2008, 28(12): 1483-1489.
[3] Li J, Glibert P M, Zhou M J, et al.Relationships between nitrogen and phosphorus forms and ratios and the development of dinoflagellate blooms in the East China Sea[J].Marine ecology-progress series, 2009,383: 11-26.
[4] 黄备, 唐静亮, 王一鸣, 等.东海赤潮发生时的营养盐分布研究[J].中国环境监测, 2013, 29(5): 108-110.
[5] 昝爱宗.浙江海域暴发大面积赤潮[N ].中国海洋报,2005-06-07.
[6] 石瑞雪, 何连生, 孟睿, 等.月牙藻、四尾栅藻生长的氮营养动力学特征[J].环境科学与技术, 2011, 34(6G):23- 26.
[7] 王小冬, 王艳.赤潮异弯藻和海洋卡盾藻抗氧化酶活性对氮磷比失衡的响应[J].海洋环境科学, 2012, 31(3):338-340.
[8] 乔方利, 袁业立, 朱明远, 等.长江口海域赤潮生态动力学模型及赤潮控制因子研究[J].海洋与湖沼, 2000,31(1): 94-99.
[9] Spitz Y, Moisan J R , Abbott M R.Configuring an ecosystem model using data from the Bermuda Atlantic Time Serie(BATS) [J].Deep-Sea ResearchⅡ , 2001, 48: 1733-1768.
[10] 高会旺.水层生态系统动力学模式参数的敏感性分析[J].青岛海洋大学学报, 1999, 29: 398-404.
[11] Estrada V, ElisaR, Parodi M, et al.Determination of biogeochemical parameters in eutrophication models with simultaneous dynamic optimization approaches [J].Computers&Chemica Engineering, 2009, 33(10): 1760-1769.
[12] 张春雷, 石晓勇, 韩秀荣, 等.营养盐对东海赤潮优势藻种生长影响的船基围隔试验[J].海洋水产研究,2006, 27: 30-35.
[13] 孙霞.光照对东海赤潮高发区赤潮藻类生长的影响[D].青岛: 中国海洋大学, 2005.
[14] 王爱军, 王修林, 王江涛, 等.光照对东海赤潮高发区春季硅藻生长的影响[J].中国海洋大学学报, 2006,36: 173-178.
[15] 孙百晔, 王修林, 李雁宾, 等.光照在东海近海原甲藻赤潮发生中的作用[J].环境科学, 2008, 29(2):363-367.
[16] Dugdale R C.Nutrientlimitation in the sea: dynamics,identification, and significance [J].Limnology and Oceanography, 1967, 12: 685-695.
[17] SchnoorJ L.Environmental Modeling[M].New York:John Wiley & Sons Inc, 1996.185-229.
[18] Laws E A, Caperon J.Carbon and nitrogen metabolism of Monochrysislutheri: measurement of growth dependent respiration rates.Marine Biology, 1976, 36:85-97.
[19] 李英, 吕颂辉, 徐宁, 等.东海原甲藻对不同磷源的利用特征[J].生态科学, 2005, 24(4): 314-317.
[20] 赵晓玮.环境中不同氮磷营养盐浓度及氮源形态对米氏凯伦藻(Karenia mikimotoi)生长的影响研究[D].青岛: 中国海洋大学, 2010.
[21] 李天深, 于仁成, 周名江.链状亚历山大藻(东海株)对磷营养物质的需求与吸收策略[J].海洋环境科学,2009, 28(4): 356-359.
[22] 郑凌凌, 宋立荣, 吴兴华, 等.汉江硅藻水华优势种的形态及18Sr rDNA序列分析[J].水生生物学报,2009, 33(3): 563-565.
[23] 张诚, 邹景忠.尖刺拟菱形藻氮磷吸收动力学以及氮磷限制下的增值特性[J].海洋与湖沼, 1997, 28(6): 600-603.
[24] 李铁, 史致丽, 仇赤斌, 等.中肋骨条藻和新月菱形藻对营养盐的吸收速率及环境因素影响的研究[J].海洋与湖沼, 1990, 30: 641-645.
[25] 李雁宾.长江口及邻近海域季节性赤潮生消过程控制机理研究[D].青岛: 中国海洋大学, 2008.
猜你喜欢
文萃报·周五版(2021年10期)2021-09-13
海洋学报(2021年2期)2021-03-15
学与玩(2019年8期)2019-10-29
资源节约与环保(2019年10期)2019-01-21
青年文学家(2015年13期)2016-05-09
青年文学家(2015年16期)2016-05-09
思维与智慧·下半月(2014年7期)2014-11-27
中国酿造(2014年9期)2014-03-11
棋艺(2001年9期)2001-07-17
棋艺(2001年11期)2001-05-21
