
外来植物豚草入侵机制
2015-04-08 06:49:14邓贞贞白加德赵彩云李俊生
草业科学 2015年1期
邓贞贞,白加德,赵彩云,李俊生
(1.中国人民大学环境学院,北京100872;2.中国环境科学研究院环境基准与风险评估国家重点实验室,北京100012;3.北京麋鹿生态实验中心,北京100076)
豚草(Ambrosia artemisiifolia)是一种被世界公认的恶性入侵杂草[1],隶属于双子叶植物纲(Dicotyledoneae)菊科(Compositae)豚草属,又名艾叶破布草、豕草、美洲艾[2-3](图1);现已从原产地美国西南部及墨西哥北部的索诺兰沙漠地区,传播至欧、美、亚、澳等各大洲的30 多个国家和地区[4]。豚草最初传入中国的时间大约为20 世纪30 年代,随着贸易活动从朝鲜传入,目前广泛分布于东北、华北、华中和华东等地区近23 个省(市、自治区)的农田、草原、荒地等生态系统和自然保护区[5-6]。豚草对入侵地生物多样性和生态系统稳定性造成了严重的负面影响[7-8];给当地农牧业造成了巨大的经济损失[9];其释放的大量花粉,成为过敏性鼻炎、枯草热症和季节性哮喘等疾病的重要过敏原,严重威胁人类健康[10-11]。2003 年,豚草被列入国家环境保护总局公布的《中国第一批外来入侵物种名单》。

图1 豚草的形态学特征Fig.1 The morphology of Ambrosia artemisiifolia
外来物种是否能成功入侵,主要取决于自身的 入侵性(Invasiveness)和生境的可入侵性(Invasibility),并受到人类干扰的影响[12]。豚草的入侵也是如此。本文从豚草入侵性、生境可入侵性和人类活动干扰3 个方面综述了豚草的入侵机制。
1 豚草的入侵性
豚草是入侵性很强的外来物种,具有较高的繁殖体压力、竞争机制和遗传多样性机制。
1.1 繁殖体压力假说
研究表明,豚草具有较高的繁殖能力、较强的传播能力、较高的种子萌发率和休眠特性,从而使得其有较大的潜力向某一区域一次传入大量的繁殖体,或多次传入同一区域,且保持较高的最终成功存活率[13-14]。而繁殖体压力假说(Propagule Pressure Hypothesis,PPH)认为这些特征共同决定了外来入侵物种的繁殖体压力,繁殖体压力越大,建立和维持种群的可能性就越大[15-16]。因此,豚草具有高水平的繁殖体压力和成功入侵概率。
1.1.1 繁殖系统特性 豚草的繁殖系统特性主要表现为:单性同株,雌花和雄花分别着生于不同的头状花序(图1D、E);传粉类型高度特化,花和花序已由菊科植物典型的虫媒传粉演化适于风媒传粉[17];以异花授粉为主产生活性种子,子代容易产生新的遗传变异,避免了自交衰退,在长远效应上有利于豚草种群的发展壮大[18];单株产种300 ~62 000粒不等,最高可达10 万粒[19],即使仅有少数豚草植株存活,也能产生足够种群自我维持的种子量[20];强大的再生能力使得刈割、践踏和折断等均不能有效地抑制豚草种子的生成,反而可能促进植株产生更多新枝和种子[21],且新枝和种子的数量随刈割高度增加而增多[22]。豚草这些繁殖系统的优点有利于其产生大量繁殖体,克服新栖息地的定居阻碍,增加入侵成功的机率[20],同时为其高单次传入量奠定了基础。
1.1.2 传播扩散特性 豚草具有短距离和长距离扩散的适应机制。短距离扩散主要是瘦果成熟后利用重力自然掉落[8],或通过风力短距离传播[9],在母株周边建立种群;长距离扩散则主要利用瘦果短喙或锥状喙(图1F)勾住人类衣物、动物皮毛,或扎入包装箱、车辆的轮胎而进行远距离传播,也可利用其海绵质果皮在水中24 h 不下沉的功能随流水长距离传播[23],此外由于豚草瘦果大小、颜色近于农作物种子而不易被发现,可以随着粮食、饲料的运输而实现远距离乃至国际间传播扩散[22,24]。从豚草的扩散方式可知,其扩散很大程度上依赖于人类活动[8],特别是交通工具的携带[25],因此其首次入侵地主要分布在公路、铁路、河道两侧及机场,其次是畜场、农田、饲料场、码头等地[26]。豚草多途径的传播扩散特性成为其高频次传入入侵地的基础,是形成较高繁殖体压力的原因之一。
1.1.3 种子萌芽特性 豚草种子萌发率一般在21% ~46%[27],最高可达到90%[28]。高低交替的温度较有利于豚草种子萌发,如在10 ℃16 h、30 ℃8 h 的交替处理下豚草种子萌发率为75%,而在20 ℃16 h、30 ℃8 h 的交替处理下为50.1%[18],这可能是昼夜温差较大的东北、华北等地区适于豚草扩散的主要原因之一[7]。埋种深度对豚草种子萌芽率影响较小,土表下9 cm 以内都可以萌发,以1 ~5 cm最适,即使在土表面仍有3.3%的萌发率[29]。但土壤含水率对豚草种子萌芽影响较大,一般适宜的土壤含水率在15% ~60%,达到100%时不萌发[29]。此外,土壤肥力也是影响豚草种子萌发率的重要因素,荒地上的种子发芽率为3.6%,花圃内达38%,玉米(Zea mays)、茄子(Solanum melongena)田的则在30% ~74%[29],人为干扰的土壤环境更适合豚草种子的萌发[30]。
1.1.4 种子休眠特性 豚草种子具有二次休眠的适应进化机制[14]。高纬度生育地的豚草种子成熟后会进入休眠状态,且纬度越高进入休眠状态的种子比例越大,解除休眠所需要的低温层积时间越长[18,31],例如我国吉林、广东两地豚草的果熟期差异不大,将两地刚采集下来的种子放在25 ℃条件下培养,吉林的豚草种子13 d 时只有4.8%解除休眠萌发成功[32],而广东的豚草种子12 d 时萌发率已达到64.4%,可见吉林的豚草种子的休眠性明显高于广东的豚草种子,这是豚草适应环境、避免幼苗被冬季低温冻死的自我保护机制[33],也是豚草种子扩散的内在动力[31]。此外,能够正常发芽的豚草种子遇到不利环境条件时会进入二次休眠,此时即使给予适合的萌发条件也不能萌发,但可以在地下经过30 ~40 年仍保持萌发能力,经过低温层积可打破休眠[31,34]。这是豚草能够形成持久种子库,保证持续活力种子来源的原因[30],也是豚草种群爆发的基础[31]。
1.2 竞争机制
1.2.1 水肥吸收能力和生长抑制作用 研究表明,豚草生产1 t 干物质一般需要消耗近1 000 t水、15 kg 氮、1.5 kg 五氧化二磷[30]。然而无论是湿润肥沃的土地还是干旱贫瘠的荒坡,都有豚草存活。这是由于豚草根、茎、叶的内部结构有利于吸取、输导和贮藏足够的水分和营养物质,且具有反射强烈的太阳光照射和降低蒸腾作用的机能[35];同时也与豚草属于专性菌根营养型植物有关[36],豚草与丛枝菌根真菌共生形成丛枝菌根可以增强自身对硝态氮、铵态氮的吸收和适应土壤氮素变化的能力,进而适应野外干旱低氮的生境[37-38],提高豚草的入侵性[39]。
与此同时,豚草强大的水肥吸收能力还对土著植物的生长起到抑制作用,其一使得豚草幼苗生长速率快,幼龄期短,从而更早地长成成株,其繁茂的分枝可以达到排挤和遮蔽其他土著植物的作用[40];其二,豚草强烈的水肥消耗往往导致土壤的干旱和贫瘠,使得土著植物因缺少水肥而生长不良[30]。
1.2.2 化感抑制作用 豚草可以通过挥发、雨水淋溶和根系分泌等途径释放萜类、烯醇类和聚乙炔类化感物质,从而强烈地抑制其周边土著植物的生长发育[41-42],研究表明该抑制作用具有选择性和阶段性,不同植物或植物的不同部位在不同生长发育阶段对豚草化感物质的反应不同[43-44]。此外,豚草种群样地中地上[45]和土壤[46]昆虫的个体和类群数量都明显少于没有豚草的对照样地,说明豚草对某些昆虫类群具有驱避作用,从而减少了昆虫对豚草的采食压力[45-47]。这可能也与豚草的化感物质有关,但相关化感物质的化学结构、性质和抑制机制等有待进一步研究[48]。
1.3 遗传多样性机制
1.3.1 表型可塑性 豚草具有很高的表型可塑性,不但植株间的叶形(图1B、C)、毛、雄花总苞大小和茎色等性状存在个体差异[30],在不同生境下,其植株大小、种子产量和千粒重均存在较大差异,其中沟边和洼地的豚草植株较高大,籽粒多,千粒重低,而生长在丘陵旱地的植株较矮小,籽粒少,千粒重高[49]。此外,豚草表型对单一的非生物环境因子存在强烈的响应,如:豚草叶片的气孔分布位置受光强变化的影响,遮阴环境下的叶片气孔下生,而高光强时气孔双面分布[50];豚草根系的发达程度受土壤水分的影响,干旱环境下的豚草往往具有异常发达的根系[51]。表型可塑性使得豚草具有更宽的生态幅和更好的耐受性,是豚草适应异质生境的重要机制[52],也是豚草能够占据广阔地理范围和多样生境的主要原因之一[53]。
1.3.2 遗传多样性 豚草具有较高的遗传多样性,这可能与其在入侵地具有多个起源,且繁殖方式以异交有性繁殖为主有关[17,54]。对东北[55]、湖南[56]和广东[57]等地豚草居群的遗传变异水平进行研究发现,豚草的遗传变异系数明显高于植物种群遗传变异系数的平均值和一年生草本植物的平均值,甚至高于植物种群中遗传变异水平最高的裸子植物[58]。然而不同地理分布区间豚草的遗传变异水平存在差异[55],以上三地豚草相比,无论是多态性位点比率,还是居群遗传分化系数,广东豚草居群的多样性指数均为最高[57]。较高的遗传多样性可以使豚草通过有性生殖产生较多遗传性不同的后代,从而更能适应不同的环境和同一地区环境的变化,入侵能力更强[57,59]。
1.3.3 生态型 生态型(Ecotype)是遗传变异和自然选择的结果[60]。关广清[61]对沈阳、铁岭、本溪、抚顺、丹东和辽阳等地区分布的豚草进行研究,认为光周期和温度等因素的差异使得豚草至少有早熟型、迟熟型和雌株型3 种生态型,其中早熟型豚草植株矮小,分枝少,始花期为7 月中、下旬,雄花序轴较少,籽实量少;迟熟型植株高大,分枝多且呈密丛状,始花期为9 月下旬,雄花序较多,单株产籽万粒以上;雌株型植株中等,分枝多,不产生雄花序,籽实数量多但体积较小。周伟等[62]对广东省广州市花都区豚草居群进行研究,发现由于珠三角地区全年气候温和,雨量充沛,适于豚草常年生长,已有少数豚草表现出两年生习性,冬季不枯死,早春现花蕾,使得花都区豚草居群与我国其他地区相比具有开花早、花量大、花期长、繁殖速度快、适应性和扩散能力更强的优势。生态型的出现和种群的分化使得豚草更能适应不同生境,入侵性更强。
2 生境可入侵性
生态系统中非生物环境和生物往往对入侵物种具有一定的抵抗力[63],豚草的成功入侵意味着传入地群落对豚草生态抵抗的失败[64],容易遭受豚草入侵的生态系统往往具有适宜豚草的气候条件和可利用资源、较低的生物多样性且缺乏高竞争力的土著植物和豚草天敌。
2.1 非生物环境对豚草的抵抗力
2.1.1 气候条件 气候条件在很大程度上影响着豚草的分布。中国许多地区的气候条件符合豚草的生活习性,对豚草入侵的抵抗力较弱,所以豚草在中国有着较广泛的分布范围[7]。郭琼霞等[4]比对了豚草的生活习性和中国气候区划特征,认为江西、陕西、山西、河南、北京、天津、湖南、江苏、安徽、浙江、上海、山东、湖北、重庆等地区为豚草适宜生长区,与马骏等[65]采用CLIMEX 生态系统软件生态气候指数模拟的结果相似,东北、华北、华中、华东和华南地区较适宜豚草生长。通过对比豚草在中国的适宜生长区和实际分布区,发现豚草在我国的江西、河南、北京、天津、湖南、江苏、安微、浙江、上海、山东和湖北等地区均已有发生记录[2,4],且其目前在我国的实际分布远未达到其最大的潜在分布范围,仍有可能继续扩散,特别是山西、陕西和重庆等省(市),虽尚未发现豚草,但由于地处华南及西南,气候条件对豚草入侵的抵抗力小,容易遭到豚草入侵[66-67];预测的适宜生长区中虽没有西藏、台湾、福建等区域,但发现这些地区已有豚草发生[2,4],可见豚草具有一定的耐受力,可以通过自身抗氧化酶系统的消除自由基作用和表型可塑性来耐受高温、降水稀少或过多等极端气候条件[51,66]。
2.1.2 资源可利用性 豚草受资源可利用性限制的可能性较小,因豚草自身具有较强的水肥利用能力和形成菌根的能力,在干旱贫瘠的环境下也能较容易地获得所需的水分和养分等生态资源[30,37];而豚草幼苗生长迅速,成株高大、枝叶繁茂,使其更容易抢夺光照资源[40]。也就是说,豚草可以通过优先抢占资源而成为先驱植物,荒坡、隙地、墙头、岩坎和石缝等资源可利用性较弱的生境对于豚草来说仍然存在一定的可入侵性[66,68]。
2.2 生物环境对豚草的抵抗力
2.2.1 物种多样性阻抗假说 物种多样性对豚草入侵的阻抗作用仍处于研究当中。但研究表明,存在强竞争力或化感作用植物的植被系统对豚草的抵抗能力较强[69]。豚草喜光,郁闭度大的生境对豚草的抵抗力较大[35],小灌木紫穗槐(Amorpha fruticosa)、沙棘(Hippophae rhamnoides)、小冠花(Coroilla varia)、草地早熟禾(Poa pratensis)、菊芋(Helianthus tuberosus)、胡枝子(Lespedeza bicolor)、紫丁香(Syringa oblata)、百脉根(Lotus tenuis)、鹰嘴紫云英(Astragalus eiar)、紫苜蓿(Medicago sativa)10 种植物可以抢在豚草旺盛生长期到来之前形成郁闭冠层,破坏豚草生长期的能量积累,使其不能正常生长发育[69];多年生禾草类植物根系庞大、盘根错节,豚草无法与其竞争,因此未退化的草原或草甸对豚草的抵抗力较强[35],研究表明,杂交狼尾草(Pennisetum americanum×P. purpureum)在高密度时可与豚草形成强烈的资源竞争[70-72];此外,生境中若存在能对豚草产生化感作用的植物[73],如白花夹竹桃(Nerium indicum)、海桐(Pittosporum tobira)、鬼针草(Bidens pilosa)、杨梅(Myrica rubra)和大叶桉(Eucalyptus robusta)等,也可能对豚草入侵产生较大的抵抗作用[43]。但自然状态下小灌木和多年生草本往往出现在演替的中后期[69],而处于演替初期的群落是豚草的主要入侵对象。
2.2.2 天敌逃逸假说 长时间缺乏天敌压力是生境对豚草抵抗力较低的原因之一。苏联、美国、加拿大三国学者曾在北美和南美的17 种豚草属植物上发现450 种天敌[74],但由于天敌并未随着豚草的入侵而自发并迅速地迁徙[4],中国2001 年才首次发现豚草的天敌昆虫——广聚萤叶甲(Ophraella communa),在此之前没有相关记录[75]。也有极少的土著广食性天敌可以取食豚草,如湖南的苍耳螟(Ostrinia orientalis)[76]和江西的二黑条豚叶甲(Zygogramma sp.)[77],但自然条件下一般发生的虫量水平难以致死植株,且优先选择土著植物为取食寄主[76,78]。此外,豚草也存在一些病原微生物天敌,如婆罗门白锈菌(Albugo tragopogortis)、万寿菊叶斑病菌(Pseudomonas syringae)等,可以在一定程度上降低豚草植株生物量、种子产量或质量[28,79]。但在自然条件下能够控制豚草的病原微生物案例尚未发现。
3 人类活动对豚草入侵的影响
国际贸易、耕作等人类活动及全球变化对豚草的入侵性和生境的可入侵性具有促进作用。
3.1 国际贸易
国际贸易活动使得豚草种子有更多机会在全球范围内被携带传播。据资料记载,豚草在欧、美、亚洲大部分国家的传播都是随着商业、农业贸易的进行而发生的,首次进入中国便是通过朝鲜农作物种子的进口[28]。从中国的进境植物检疫记录中也发现,小麦(Triticum aestivum)、大豆(Glycine max)、波斯菊(Cosmos bipinnata)、玉米(Zea mays)、羊毛等都可能携带豚草种子入境[25]。仅2000 年,从俄罗斯进口的大豆中就检出豚草种子高达45 次[80]。因此,内检及外检是豚草防控工作的重要手段[26],中国农业部于2006 年发布的《全国农业植物检疫性有害生物名单》,以及2007 年发布的《中华人民共和国进境植物检疫性有害生物名录》都将豚草属植物列为检疫对象。
3.2 频繁干扰
人类的频繁干扰可能降低生境对豚草入侵的抵抗力,促进豚草种子萌发,并提高豚草的生长、繁殖和传播扩散能力。刈割、践踏等人类的频繁干扰不利于许多土著杂草的生长,从而降低了生物多样性和生态系统稳定性,使该生态系统抵抗豚草入侵的能力变弱[14];而这些行为却很难使豚草致死,反而促使豚草生成更多的新枝和瘦果,从而克服定居障碍,产生较多的繁殖体[20,30]。土地翻耕等活动有可能将土壤深处二次休眠的种子带到土壤表面,这些种子在经历低温解除休眠之后得以萌发,形成新的 入 侵[31];Macdonald 和Kotanen[81]研 究 也 表 明,土壤的扰动强烈促进了豚草成株的生长和存活。此外,人类的频繁活动也使得豚草瘦果有更多的机会勾住人类衣物或扎入包装箱和车辆轮胎,从而被携带传播到新的区域[23]。
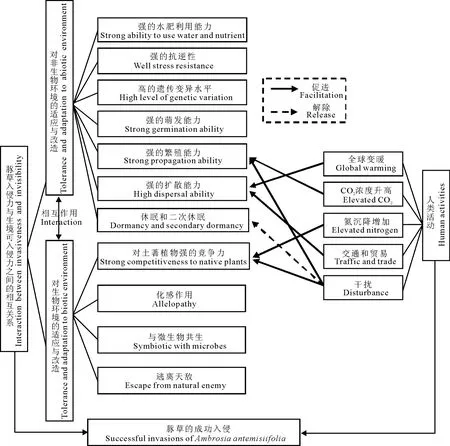
图2 豚草在生态系统中成功入侵的机制Fig.2 Mechanisms of the successful invasion of Ambrosia artemisiifolia in ecosystems
3.3 全球变化
人类活动对气候变暖、CO2浓度和氮沉降明显增加等全球变化的驱动是至关重要的[82],而这些变化将可能提高豚草的入侵力,促进这一气候适应范围广的入侵植物的传播扩散[83-85]。研究表明,全球变暖将大大提高豚草在欧洲大部分地区的入侵潜力,使其种群扩大并向北扩散[83];CO2浓度升高会使豚草植株生长速度加快,植株大小、生物量和花粉产量都有所增加,补偿了后来生长季节中任何有关的不利条件,最终有利豚草的生长和繁殖[86];此外,氮沉降的增加有利于豚草这种能将氮迅速转化为新生物量的速生种,但却不利于生长缓慢的土著物种[87],王晋萍等[84]的取代系列试验也表明,氮素的添加显著提高了豚草的种间竞争力,使豚草在与土著种的竞争中占据优势。
综上所述,豚草的入侵可以概括为全球变化背景下,豚草自身的入侵性和生境的可入侵性的协同作用(图2)。
4 展望
豚草入侵机制的研究是充分理解豚草成功入侵的核心基础,有助于预测豚草未来的入侵趋势,并为豚草的防控提供较好的理论支持。然而豚草的入侵机制是复杂多变的,在未来的研究中应注意以下方面:1)豚草的生物学特性对其成功入侵的作用还有待于进一步研究,如从繁殖体压力假说着手,研究单次传入的个体数量和传入次数,验证豚草生物学特征对豚草成功建立种群的作用,探讨豚草的生物学特性与入侵力之间的关系。2)豚草的表型变化和遗传多样性是解释其成功入侵的原因之一,我国虽然开展了部分研究,发现豚草在不同地区出现不同的生态型,表明入侵我国的豚草具有丰富的遗传多样性,但是还需要进一步开展同质园试验,分析豚草表型可塑性和遗传多样性与入侵力之间的关系,明确不同地理种群应对环境胁迫的机制。3)在豚草入侵机制的研究中,豚草与土壤生物、微生物的相互作用尚未揭示,哪些微生物的存在促进了豚草的入侵?豚草如何通过化感作用趋避限制其生长的微生物?均需要进一步通过研究证实,建议豚草未来的研究工作可以尝试由地上过程转向地下过程。4)生境可入侵性是外来物种成功入侵的主要因素之一,物种丰富度假说认为物种丰富度越高的群落外来物种入侵越困难,而不同尺度上的研究结论差异较大。目前,有关功能群的丰富度在解释外来物种入侵与群落稳定性之间关系的研究较多,未来可以从功能群组成、不同空间尺度、不同试验方法等角度探讨豚草的入侵机制。
[1] 齐国君,高燕,钟锋,邵晓迎,吕利华.广聚萤叶甲对广东佛冈豚草的控制作用[J].环境昆虫学报,2012,34(3):315-321.
[2] 吴海荣,强胜,段惠,林金成.豚草(Ambrosia artemisiifolia L.)[J].杂草科学,2004(2):50-52.
[3] 康芬芬,魏亚东,杨菲,张瑞峰,程瑜,刘勇,印丽萍.不同处理对豚草种子休眠与萌发的影响[J].植物检疫,2010,24(6):14-16.
[4] 郭琼霞,黄可辉,刘景苗.美洲豚草(Ambrosia artemisiifolia)风险分析[J].武夷科学,2004,20(1):115-121.
[5] 王建军,赵宝玉,李明涛,李蓉.生态入侵植物豚草及其综合防治[J].草业科学,2006,23(4):71-75.
[6] 李会娜,刘万学,万方浩,曹远银.入侵植物豚草与本地植物马唐对土壤肥力与酶活性影响的比较[J].中国生态农业学报,2009,17(5):847-850.
[7] 周忠实,郭建英,李保平,孟玲,傅建炜,陈红松,马明勇,史梦竹,李敏,郭薇,罗敏,郑兴汶,郑海燕,罗源华,万方浩.豚草和空心莲子草分布与区域减灾策略[J].生物安全学报,2011(4):263-266.
[8] Sǎrǎteanu V,Moisuc A,Cotuna O.Ambrosia artemisiifolia L.An invasive weed from ruderal areas to disturbed grasslands[J].Seria Agronomie,2010,53:235-238.
[9] 张葵.恶性杂草——豚草[J].生物学通报,2006,41(2):25-26.
[10] Mandrioli P,Di Cecco M,Andina G.Ragweed pollen:The aeroallergen is spreading in Italy[J].Aerobiologia,1998,14(1):13-20.
[11] Bass D J,Delpech V,Beard J,Bass P,Walls R S.Ragweed in Australia[J].Aerobiologia,2000,16:107-111.
[12] Lodge D M.Biological invasions:Lessons for ecology[J].Trends in Ecology and Evolution,1993,8(4):133-137.
[13] Essl F,Dullinger S,Kleinbauer I.Changes in the spatio-temporal patterns and habitat preferences of Ambrosia artemisiifolia during its invasion of Austria[J].Preslia,2009,81(2):119-133.
[14] 李建东,殷萍萍,孙备,孙佳楠,王国骄,燕雪飞.外来种豚草入侵的过程与机制[J].生态环境学报,2009,18(4):1560-1564.
[15] Memmott J,Graze P G,Harman H M,Syrett P,Fowler S V.The effect of propagule size on the invasion of an alien insect[J].Journal of Animal Ecology,2005,74:50-62.
[16] Maron J L.The relative importance of latitude matching and propagule pressure in the colonization success of an invasive forb[J].Ecography,2006,29(6):819-826.
[17] 关广清.豚草花器结构及排粉过程[J].植物检疫,1993(3):167-170.
[18] Basset I J,Crompton C W.The biology of Canadian weeds:11.Ambrosia artemisiifolia L.and A.psilostachya DC[J].Canadian Journal of Plant Science,1975,55(2):463-476.
[19] 王洪林,王芝恩,凌帅.浅析辽宁中东部地区豚草分布及防控策略[J].辽宁林业科技,2010(4):39-40,54.
[20] Warren R J,Bahn V,Bradford M A.The interaction between propagule pressure,habitat suitability and density-dependent reproduction in species invasion[J].Oikos,2012,121(6):874-881.
[21] 温有学,于立山,郑兰芬.大田作物的杀手——豚草[J].吉林农业,2003(10):16.
[22] 杜淑梅,姚兴举.黑龙江省豚草发生种类、分布及综合防除措施[J].中国植保导刊,2007,27(4):39-40.
[23] 曾珂,朱玉琼,刘家熙.豚草属植物研究进展[J].草业学报,2010,19(4):212-219.
[24] Bohren C.Ambrosia artemisiifolia L. -in Switzerland:Concerted action to prevent further spreading[J]. Nachrichtenblatt des Deutschen Pflanzenschutzdienstes,2006,58(11):304-308.
[25] 吴海荣,胡学难,吕利华,王卫芳,冯黎霞,崔汝强,何自福,周国辉.豚草在广东韶关的分布调查及防控建议[J].植物检疫,2009,23(2):63-64.
[26] 曲波,吕国忠,杨红,刘绍芹.豚草属植物的研究进展[J].安徽农业科学,2006,34(7):1387-1388.
[27] 罗艳,刘爱华.青岛外来入侵植物的初步研究[J].山东科学,2008,21(4):19-23.
[28] 王明勇.安徽省豚草发生现状与控制对策[J].安徽农业科学,2005,33(9):1771-1786.
[29] 段惠萍,陈碧莲.豚草生物学特性、为害习性及防除策略[J].上海农业学报,2000,16(3):73-77.
[30] 闫恩维,罗春玲,胡瑛.牡丹江地区豚草特性及危害[J].中国林副特产,2009(2):108-109.
[31] 王志西,刘祥君,高亦珂,祖元刚.豚草和三裂叶豚草种子休眠规律研究[J].植物研究,1999,19(2):158-164.
[32] 杨逢建,张衷华,王文杰,祖元刚,陈华峰,贾晶,关宇,张乃静.八种菊科外来植物种子形态与生理生化特征的差异[J].生态学报,2007,27(2):442-449.
[33] 冯莉,岳茂峰,田兴山,齐国君,吕利华.豚草在广东的分布及其生长发育特性[J].生物安全学报,2012(3):210-215.
[34] Squiers E R.The effects of seasonal timing of disturbance on species composition in a first-year old field[J]. Bulletin of the Torrey Botanical Club,1989,116(4):356-363.
[35] 邢艳芳.豚草和三裂叶豚草在吉林省内的分布及解剖结构研究[D].长春:东北师范大学,2012.
[36] Crowell H F,Boerner R E J.Influences of mycorrhizae and phosphorus on belowground competition between two old-field annuals[J].Environmental and Experimental Botany,1988,28:381-392.
[37] 黄栋,桑卫国,朱丽.氮碳添加和丛枝菌根对外来入侵植物豚草的影响[J].应用生态学报,2010,21(12):3056-3062.
[38] Medve R J.The mycorrhizae of pioneer species in disturbed ecosystems in western Pennsylvania[J].American Journal of Botany,1984,71:787-794.
[39] 于文清,张利莉,刘万学,万方浩.土壤真菌差异影响入侵豚草与本地植物生长及互作[J].生态学杂志,2010,29(3):523-528.
[40] Rejmáek M,Richardson D M.What attributes make some plant species more invasive[J].Ecology,1996,77(6):1655-1661.
[41] 王大力,祝心如.豚草的化感作用研究[J].生态学报,1996,16(1):11-19.
[42] 杨逢建,王纪坤,张莹,张衷华,陈华峰,祖元刚. 林业有害植物豚草挥发油GC-MS 成分分析[J]. 植物研究,2005,25(4):457-459.
[43] 陈贤兴,单和好.豚草对几种经济作物的生化他感作用[J].海南大学学报(自然科学版),2003,21(1):69-73.
[44] 张风娟,郭建英,龙茹,万方浩.不同处理的豚草残留物对小麦的化感作用[J].生态学杂志,2010,29(4):669-673.
[45] 史彩华,王福莲,刘万学.保护地豚草昆虫群落结构及动态研究[J].长江大学学报(自然科学版),2009(4):13-15.
[46] 孙刚,房岩,殷秀琴.豚草发生地土壤昆虫群落结构及动态[J].昆虫学报,2006,49(2):271-276.
[47] 黄红英,徐剑,邹佩贞,石海英,张明.豚草不同生长时期土壤动物的群落结构特点[J].广东农业科学,2010,37(10):165-168.
[48] Smith M,Cecchi L,Skjøth C A,Karrer G,Škoparija B.Common ragweed:A threat to environmental health in Europe[J].Environment International,2013,61:115-126.
[49] 刘香梅,张茂伟.美洲豚草的发生调查与防治研究[J].中国农学通报,1995,11(3):40-41.
[50] Mott K A,Gibson A C,O’Leary J W.The adaptive significance of amphistomatic leaves[J].Plant,Cell & Environment,1982,5(6):455-460.
[51] 陈永亭,吴降星,陆军良.豚草的生物学特性及防除[J].安徽农学通报,2006,12(4):136.
[52] Sultan S E.Phenotypic plasticity and plant adaptation[J].Acta Botanica Neerlandica,1995,44:363-383.
[53] 耿宇鹏,张文驹,李博,陈家宽.表型可塑性与外来植物的入侵能力[J].生物多样性,2004,12(4):447-455.
[54] Jahodová Š,Trybush S,Pyšek P,Wade M,Karp A.Invasive species of Heracleum in Europe:An insight into genetic relationships and invasion history[J].Diversity and Distributions,2007,13(1):99-114.
[55] 沙伟,周福军,祖元刚.东北地区豚草种群的遗传变异与遗传分化[J].植物研究,1999,19(4):452-456.
[56] 邓旭,王娟,谭济才.湖南省不同居群豚草种群遗传多样性ISSR 分析[J].广东农业科学,2011,38(14):124-127.
[57] 黄久香,刘宪宽,庄雪影,翟翠花,黄川腾,徐瑞晶.广东豚草居群的遗传分化[J].广东农业科学,2012,39(3):135-138.
[58] Hamrick J L,Godt M J W.Allozyme diversity in plant species[A].Brown A H D,Clegg M T,Kahler A L,Weir B S(eds).Plant Population Genetic,Breeding,and Genetic Resources[C].Sunderland:Sinauer Associates,1990:23-63.
[59] 祖元刚,王文杰,杨逢建,于景华,曹建国,赵则海.植物生活史型的多样性及动态分析[J].生态学报,2002,22(11):1811-1818.
[60] 郭兰萍,黄璐琦,华永丽,陈美兰. 从表型可塑性,生态型到药用植物化学型的研究[J]. 资源科学,2008,30(5):744-753.
[61] 关广清.豚草和三裂叶豚草的形态特征和变异类型[J].沈阳农学院学报,1985(4):9-17.
[62] 周伟,徐瑞晶,赵倩,谢亚利,庄雪影.广州市花都区豚草种群监测调查[J].杂草科学,2010(3):9-13.
[63] 王卿,安树青,马志军,赵斌,陈家宽,李博. 入侵植物互花米草——生物学、生态学及管理[J]. 植物分类学报,2006,44(5):559-588.
[64] Eschtruth A K,Battles J J.Assessing the relative importance of disturbance,herbivory,diversity,and propagule pressure in exotic plant invasion[J].Ecological Monographs,2009,79:265-280.
[65] 马骏,万方浩,郭建英,游兰韶,卢德勇. 豚草卷蛾在我国的生物气候相似性分析[J]. 中国农业科学,2003,36(10):1156-1162.
[66] 邓旭,王娟,谭济才.外来入侵种豚草对不同环境胁迫的生理响应[J].植物生理学通讯,2010,46(10):1013-1019.
[67] 陈浩,陈利军,Albright T P.以豚草为例利用GIS 和信息理论的方法预测外来入侵物种在中国的潜在分布[J].科学通报,2007,52(5):555-561.
[68] Lake J C,Leishman M R.Invasion success of exotic plants in natural ecosystems:The role of disturbance,plant attributes and freedom from herbivores[J].Biological Conservation,2004,117(2):215-226.
[69] 关广清.经济植物替代控制豚草的研究[J].沈阳农业大学学报,1995,26(3):277-283.
[70] Austin M P,Fresco L F M,Nicholls A O,Groves R H,Kaye P E.Competition and relative yield:Estimation and interpretation at different densities and under various nutrient concentrations using Silybum marianum and Cirsium vulgare[J].The Journal of Ecology,1988,76:157-171.
[71] Wilson J B.Shoot competition and root competition[J].Journal of Applied Ecology,1988,25:279-296.
[72] 岳茂峰,冯莉,田兴山,杨彩宏.杂交狼尾草与豚草的竞争效应[J].广东农业科学,2013,40(8):83-86.
[73] Csiszár Á,Korda M,Schmidt D,Šporci'C D,Suele P,Teleki B,Tiborcz V,Zagyvai G,Bartha D.Allelopathic potential of some invasive plant species occurring in Hungary[J].Allelopathy Journal,2013,31(2):309-318.
[74] Harris P,Piper G L.Ragweed (Ambrosia spp.:Compositae):Its North American insects and the possibilities for its biological control[J].Technical Bulletin.Commonwealth Institute of Biological Control,1970,13:117-140.
[75] 孟玲,李保平.新近传入我国大陆取食豚草的广聚萤叶甲[J].中国生物防治,2005,21(2):65-69.
[76] 万方浩,马骏,郭建英,游兰韶.豚草卷蛾和苍耳螟对豚草的联合控制作用[J].昆虫学报,2003,46(4):473-478.
[77] 吴德龙,魏洪义,桂爱礼.二黑条豚叶甲对豚草控制初步调查[J].江西植保,2004,27(4):155-156.
[78] 李宏科,李彦宁.湖南豚草病原菌调查简报[J].生物防治通报,1993(1):45-46.
[79] 李宏科,李萌,李丹.豚草及其防治概况[J].世界农业,1999(8):40-41.
[80] 郑超,张箭,吴永生,何浩.对俄贸易中豚草的检疫地位[J].植物保护,2001,27(5):47-48.
[81] MacDonald A A M,Kotanen P M.The effects of disturbance and enemy exclusion on performance of an invasive species,common ragweed,in its native range[J].Oecologia,2010,162(4):977-986.
[82] Holland E A,Dentener F J,Braswell B H,Sulzman J M.Contemporary and pre-industrial global reactive nitrogen budgets[A].New Perspectives on Nitrogen Cycling in the Temperate and Tropical Americas[M].Netherlands:Springer,1999:37-43.
[83] Cunze S,Leiblein M C,Tackenberg O.Range expansion of Ambrosia artemisiifolia in Europe is promoted by climate change[J].ISRN Ecology,2013,doi:10.1155/2013/610126.
[84] 王晋萍,董丽佳,桑卫国.不同氮素水平下入侵种豚草与本地种黄花蒿、蒙古蒿的竞争关系[J].生物多样性,2012,20(1):3-11.
[85] Hellmann J J,Byers J E,Bierwagen B G,Dukes J S.Five potential consequences of climate change for invasive species[J].Conservation Biology,2008,22(3):534-543.
[86] Rogers C A,Wayne P,Macklin E,Muilenberg M L,Wagner C W,Epstein P R,Bazzaz F A.Interaction of the onset of spring and elevated atmospheric CO2on ragweed (Ambrosia artemisiifolia L.)pollen production[J].Environmental Health Perspectives,2006,114:865-869.
[87] Nordin A,Strengbom J,Witzell J,Näsholm T,Ericson L.Nitrogen deposition and the biodiversity of boreal forests:Implications for the nitrogen critical load[J].AMBIO:A Journal of the Human Environment,2005,34(1):20-24.
猜你喜欢
草食家畜(2022年5期)2023-01-02 02:53:26
儿童时代·幸福宝宝(2019年9期)2019-10-28 18:04:52
幼儿园(2018年15期)2018-10-15 19:40:36
莫愁·家教与成才(2017年7期)2017-07-11 21:31:47
红领巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
新农业(2016年19期)2016-08-16 12:00:18
少儿科学周刊·儿童版(2015年10期)2015-11-07 03:42:03
少儿科学周刊·儿童版(2015年1期)2015-07-07 04:12:52
湖南农业(2014年7期)2014-04-30 08:25:08
