
放牧强度对三江源典型高寒草甸生物量和土壤理化特征的影响
2015-04-08 06:49:12李凤霞李晓东周秉荣祁栋林王力傅华
草业科学 2015年1期
李凤霞,李晓东,周秉荣,祁栋林,王力,傅华
( 1. 青海省气象科学研究所,青海西宁810001; 2. 兰州大学草地农业科技学院,甘肃兰州730020)
全球草地面积约为3.42 ×109hm2,约占地球陆地面积的40%,是与陆地净初级生产力有关碳通量的重要组成部分[1]。另外,草地贮存着大于10%的陆地生物量碳,10% ~30%的全球土壤有机碳。草地生态系统是重要的碳汇,因此草地生态系统适度的碳库变化可以潜在地修复全球碳循环以及气候变暖的直接影响[2]。过度放牧和草地开垦被认为是人类对草地生态系统的两种主要干扰方式,其中过度放牧是人类对草地生态系统影响最为强烈的因素之一;全世界草地退化面积中,由于过度放牧而造成的草地退化面积约占35%,其影响规模远远超过草地开垦[3]。家畜放牧是世界草地普遍存在的最主要的草地利用方式,这种利用方式有可能从本质上改变草地生态系统的碳库:改变地上―地下生物量的分配从而改变碳库[4];改变微气候环境和光、水以及营养的有效性[5];通过改变物种组成和植物群落的多样性功能影响向土壤输入碳的数量和质量[6]。虽然放牧加速了放牧生态系统的碳循环过程,但是放牧导致的对生态系统碳库的影响常常是不一致和难以预测的。
过度放牧也是土壤荒漠化的原因之一,我国沙漠化土壤的28.3%是以过度放牧为驱动因素形成的。放牧干扰主要通过家畜采食和践踏影响土壤的物理结构,通过家畜采食活动及牲畜对营养物质的转化和排泄物归还等影响草地营养物质的循环,从而导致草地土壤化学成分发生变化,而草地土壤物理和化学变化之间也相互作用、相互影响[7]。随着放牧强度的提高,牲畜对土壤的压实作用增强,土壤容重也逐渐增加[4]。高寒草甸[8]、东北羊草(Leymus chinensis)草原[9]和内蒙古典型草原[10]土壤容重随着放牧强度增加逐渐增大,放牧对土壤容重的增加具有累积效应。过度放牧导致土壤容重和渗透阻力增加,土壤结构的空间分布发生变化,从而导致土壤通气性、渗透性和蓄水能力受到不良影响[11]。草地初级生产固定的碳素由于过度放牧向土壤的分配比例减少,家畜采食行为减少了由植物向土壤的碳素输入;同时过度放牧改变了土壤表层结构且增强了土壤呼吸作用,从而加快了土壤碳素向大气的释放[3]。Li 和Chen[12]研究,近40 年来,过度放牧导致内蒙古锡林河流域羊草草原表层土壤有机碳贮量减少了约12.4%。傅华等[13]研究表明,阿拉善左旗贺兰山前草原化荒漠区15 年持续过度放牧后,致使0 ― 20 cm 土层土壤有机碳含量下降了25.2%。有研究认为,随着放牧强度的逐渐增加,受放牧干扰后土壤中全氮含量会逐渐降低[14],王长庭等[15]研究提出,高寒草甸重度放牧的土壤全氮比轻度放牧降低了45%,也有研究认为,土壤全氮随放牧强度的增加而增加[16]。Trumbore 等[17]认为,土壤养分下降的主要原因之一是草地的过度放牧使草地生产力下降;而围封管理的草地,草地生产力始终保持在相对稳定的水平,土壤养分具有较高的增加潜力,因此较为稳定的草地生产力水平和草地根系向土壤的养分输入是保持和提高土壤肥力的关键因子。
青藏高原是受人为影响相对较少且面积广大的半自然区域,在全球气候变化的大背景下,高原春冬两季和地表温度都明显升高[17-20]。高寒草甸面积大约占青藏高原面积的35%,该区尤其在高原东部地区具有代表性的植被类型并且是最主要的放牧草地。高寒生态系统中的土壤含有较高的养分,因为高寒生态系统在生长季具有高产量且由于低温导致分解速率相对较低。放牧干扰是该地区草地生态系统影响土壤养分收支的一个关键因子,但是在高寒草甸生态系统中长期过度放牧将导致草地土壤养分退化甚至荒漠化,并且退化或者荒漠化的草地可能释放大量的碳进入大气,使高寒生态系统也有可能由于高强度的放牧成为重要的碳源生态系统。因此,本研究通过为期两年的不同放牧强度试验对高寒草甸植被和土壤碳、氮影响的研究,探讨放牧对高寒草甸植被、土壤质量以及碳氮特征的影响,以期为青藏高原草地生态系统和草地畜牧业的可持续发展提供理论依据。
1 研究区与研究方法
1.1 研究区概况
研究区地处青海省玉树州境内,位于玉树县隆宝滩,地理坐标为96°25' - 96°37' E,33°08' -33°14' N。海拔4 200 m,年降水量480 mm,年平均气温2.9 ℃,具有典型大陆性气候特征。该地区主要牧草种类以莎草科植物为主,有藏嵩草(Kobresia tibetica)、高山嵩草(K. pygmaea)、矮嵩草(K. humilis)、华扁穗草(Blysmus sinocompresus)等优势牧草,土壤类型为高寒沼泽土[21]。该区气候寒冷,生境潮湿,雨量充沛,沼泽遍地,属于典型的沼泽草甸和高山草甸区。区域主要由草地、沼泽、灌木林、裸岩、永久性雪地冰川、湖泊、河流、灌草过渡带、宜林和不宜林草地及其他水域组成,境内孕育有“三江源”地区典型湿地代表——玉树隆宝滩湿地,湿地属于“隆宝滩黑颈鹤自然保护区”。由于近年来过度放牧、气候变化导致干旱等原因,呈高寒沼泽湿地―高寒草甸―黑土滩的退化趋势[22]。
1.2 研究方法
1.2.1 样地设置 研究区设5 种不同处理:围栏封育(EN)、轻度放牧(LG)、中度放牧(MG)和重度放牧(HG)4 个放牧梯度以及对照区(CK),对照区是不受任何管理方式干扰的自由放牧草地。所有样地均从2006 年开始封育或放牧,土样及其他样品采集均从2008 年开始,2006 年之前各样地均为自由放牧地。由于玉树隆宝滩为高寒沼泽草地,可食鲜草产量约288.1 kg·亩-1(约合4 321.5 kg·hm-2),以每只羊每天4. 5 kg 采食量,采食70% 计[21],(4.5 ×180)/(288. 1 ×70%)≈4 亩·只-1(约合0.267 9 hm2·只-1),即在玉树隆宝滩夏季(180 d)放牧时,平均以每4 亩(0.267 9 hm2)草地放牧一只羊为宜。按此标准,上下浮动50%,制定重牧和轻牧标准。重牧区为2 亩· 只-1,中牧区为4亩·只-1,轻牧区为6 亩·只-1。对于自由放牧区,经实地调查,其放牧强度大于1 亩·只-1。
1.2.2 样品采集 试验对草地生物量和土壤样品的采集分别于2008 年和2009 年的5 月上旬到9 月上旬每月测定一次。土壤样品采用直径为50 mm的土钻5 点“S”形取样,每个处理内设3 个样区,每个样区内随机取3 个重复。每个处理两年共采集90 个样本。试验中测定的土壤水分参数为土壤重量含水量,测定方法用土钻法,测定深度为50 cm,10 cm 深度为一层,共5 层。由于高原地区长年枯落物以及根系不能完全分解导致土壤上层0 ―20 cm 内积累大量的植物残余物,因此土壤容重均主要测定20 cm 以下土层,而土壤有机碳、土壤全氮测定20 ―30 cm 土层。其中土壤湿度和土壤容重均为2009 年测定值。
草地地上生物量测定在各样区内随机设置1 m×1 m 样方,用剪刀齐地面采集植被的地上部分,然后烘干至恒重称取生物量作为每次测定生物量。草地地下生物量测定通过根钻钻取测定地上生物量的样方内的土壤,测定深度为50 cm,10 cm 深度为一层,共5 层。通过去土洗根的方法获取植物根系,然后烘干至恒重称取生物量作为每次测定草地的地下生物量。
1.2.3 样品分析 土壤样品室内风干,去除残留根系用玻璃管压碎,分出部分样品过0.5 mm 筛,测定有机碳、全氮。有机碳采用重铬酸钾容量法,全氮用10 mL 硫酸,加3.3 g 催化剂(K2SO4/CuSO4·5H2O =10/1)消化,采用连续流动分析仪测定(AA3)。
1.2.4 数据分析 采用Excel 2003 作图,用SPSS 18.0软件对试验数据进行统计分析。采用LSD 法对不同处理和不同土层的试验数据进行单因素方差分析。
2 结果与分析
2.1 短期放牧对草地生物量的影响
放牧对高寒草甸生态系统的草地植物、土壤和微环境等具有非常重要的影响,地上生物量的多少直接反映了生态系统的水平及所贮藏物质的多少。同样,放牧干扰下,草地地上生物量变化也可以直接表征草地的变化趋势和健康水平。本试验发现,随着放牧强度的增加,草地生态系统地上生物量呈现减少的趋势。2009 年地上生物量各处理都明显高于2008 年,其中封育区、轻牧区和中牧区2008 年和2009 年地上生物量差异显著(P <0.05),其他样地两年之间没有明显差异(图1)。地上生物量在2008 和2009 年封育区均显著高于其他处理,其次是轻牧区,重牧区和对照区均较低。
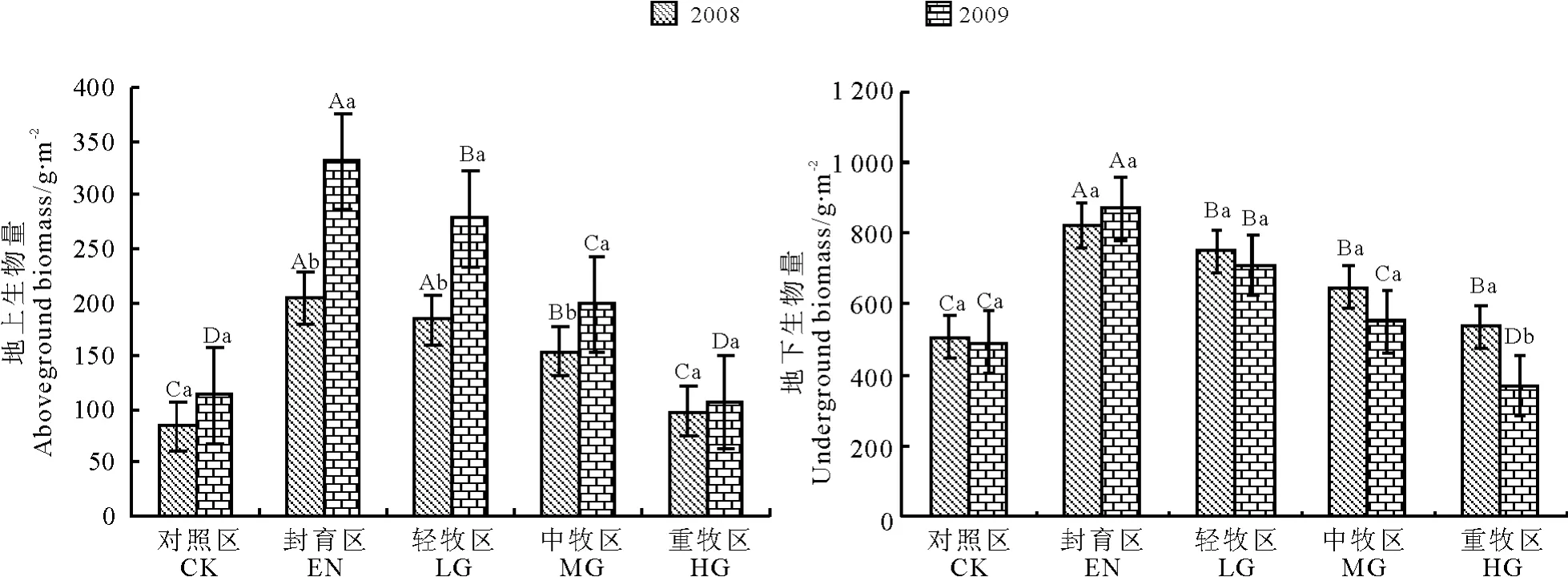
图1 放牧处理对高寒草甸地上和地下生物量的影响Fig.1 Effects of grazing on aboveground and underground biomass
植被地下部分主要指位于土壤层内的根系部分,根系起着支持与固定植物地上部分的作用,同时是植物吸收土壤中水分和养分的重要器官,是营养物质的贮存器,也是多种激素与生长调节剂的合成与制造器官。在草地生态系统中,根系又是向土壤中归还输入碳素及氮素的重要形式;因此,植被地下部分根系研究是探讨草地生态系统结构与功能不可或缺的部分之一。分析发现,草地在放牧的干扰下,两年的地下生物量均以封育区最高,对照区和重牧区较低。封育区2008 年和2009 年积累的地下生物量最高,分别达823.6 和868.0 g·m-2,重牧区2009 年最低,仅为369.6 g·m-2,相比2008 年减少了168.27 g·m-2,说明在重度放牧条件下地下生物量损失较大(图1)。2008 年和2009 年除了重牧区两年地下生物量有显著差异外,其余样地两年处理均没有明显差异(P <0.05)。地下生物量在2008 年和2009 年封育区均显著高于其他处理(P <0.05)。
2.2 短期放牧对草地土壤湿度的影响
放牧干扰改变了地表覆盖状况,进一步对高寒草甸生态系统的土壤湿度产生相应的影响,而土壤湿度的减少也是高寒草甸生态系统退化的强信号。不同放牧处理下0 ―50 cm 各土层土壤湿度由高到低依次为封育区>轻牧区>中牧区>对照区>重牧区(图2),封育区和轻牧区显著高于其他处理(P <0.05)。其中封育区10 cm 的土壤湿度最高,达143.9%,分别比对照区、轻牧区、中牧区和重牧区高出45.4、6.7、27.7 和56.6 百分点。在土壤剖面上,各处理表层土壤湿度均高于底层,0 ―50 cm 土层土壤湿度土壤表层最高且随土壤深度的增加呈明显降低趋势。
2.3 短期放牧对草地土壤容重的影响
家畜通过放牧践踏作用可以影响土壤结构,从而影响土壤容重。随着家畜放牧强度的增加,增强的践踏作用可能将进一步增加土壤容重。通过2009 年的试验数据分析得出(图3),20 ―30、30 ―40 和40 ― 50 cm 各土层土壤容重重牧区均为最高,而封育区最低。重牧区土壤容重在20 ― 30、30 ―40 和40 ―50 cm 各土层分别为1.23、1.18 和1.34 g·cm-3,土壤容重最低的封育区各土层土壤容重仅为0.48、0.49 和0.63 g·cm-3。
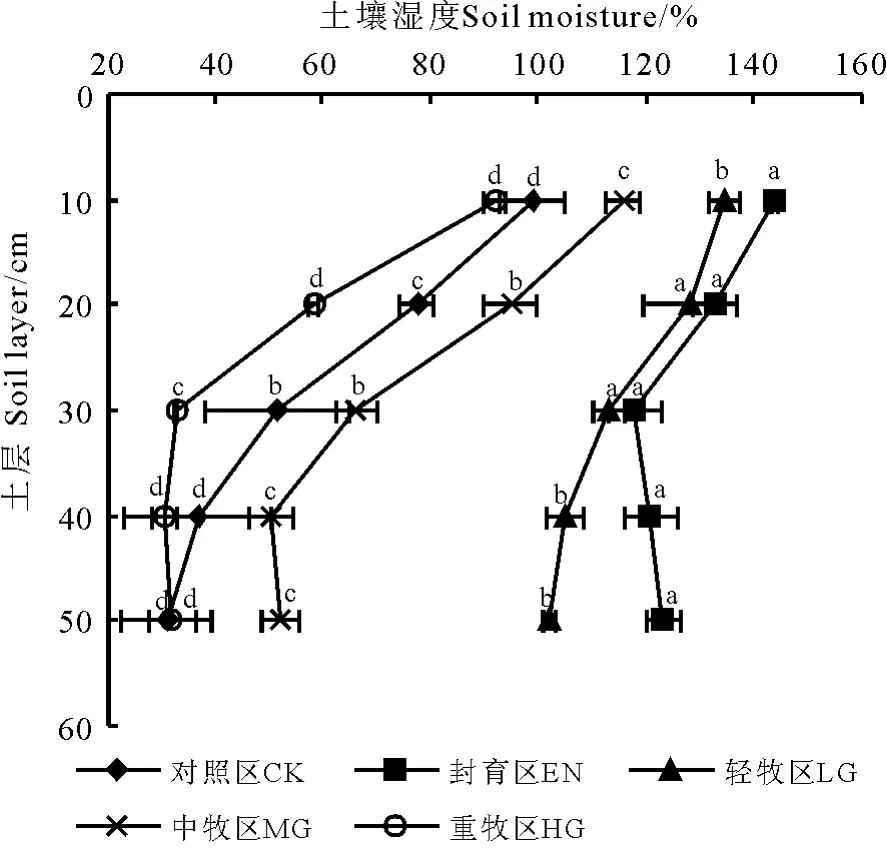
图2 放牧处理对高寒草甸土壤湿度的影响Fig.2 Effects of different grazing intensity on soil moisture

图3 放牧处理对高寒草甸土壤容重的影响Fig.3 Effects of grazing on soil bulk density
2.4 短期放牧对土壤全氮的影响
土壤氮含量直接或间接地受家畜放牧过程中采食、践踏和排泄等行为的影响。本研究中,2008 年和2009 年各处理土壤全氮含量均以轻牧区最高,其次是封育区、中牧区,对照区和重牧区土壤全氮含量较低(图4)。轻牧区2008 年和2009 年土壤全氮含量分别达11.39 和11.57 g·kg-1,而重牧区土壤全氮含量仅为7.76 和8.08 g·kg-1。2008 年和2009年轻牧区分别比对照区高出50.3%和39.9%,中牧区分别比对照区高出29.3%和10.9%,可见适度的放牧能提高土壤中氮素含量,增加土壤肥力,将有助于草地生态系统的良性发展。

图4 放牧处理对高寒草甸土壤全氮的影响Fig.4 Effects of grazing on soil total nitrogen
2.5 短期放牧处理对土壤有机碳的影响
通过两年试验研究得出(图5),玉树隆宝滩地区高寒草甸放牧处理条件下2008 年和2009 年各处理土壤有机碳含量从高到底依次为轻牧区>封育区>中牧区>重牧区>对照区,其中轻牧区在2008年和2009 年最高,分别达156.5 和176.2 g·kg-1,而对照区在2008 年和2009 年均最低,仅为90.8 和102.3 g·kg-1;2008 年土壤有机碳含量轻牧区比对照区和重牧区分别高出72. 4%和48.5%,而2009年土壤有机碳含量轻牧区比对照区和重牧区分别高出72.2%和62.9%。

图5 放牧处理条件下高寒草甸土壤有机碳变化Fig.5 Soil organic carbon under different grazing intensities
3 讨论
放牧是草地利用的主要方式之一,而不合理的放牧活动是草地退化的主要原因[23]。汪诗平等[24]研究发现,适度放牧条件下,草地生态系统的年净初级生产力能通过植物的补偿性生长增加,本研究显示在轻度放牧和中度放牧条件下,草地地上地下生物量均显著高于对照区(1 亩·只-1)。封育区由于长期没有受到放牧干扰,其积累的地上地下生物量最高。周华坤等[25]的研究也表明,随着放牧强度的增加,高寒植被群落地上现存生物量降低明显,也有研究指出,在不同放牧处理下,地上生物量与放牧强度显著负相关[26],均与本研究结果一致。
本研究发现,玉树隆宝滩地区高寒草甸表层土壤湿度呈过饱和状态,但随着放牧强度的增加而减少,土壤湿度在各土层从大到小依次为封育区>轻牧区>中牧区>重牧区。这与张伟华等[27]对内蒙古草原不同放牧强度下土壤湿度的变化趋势研究是一致的。由于放牧强度的增加,牲畜啃食与践踏强度加大,草地表层土壤覆盖度减小,土壤蒸发也可能相应增大,因而导致土壤持水能力下降最终使土壤湿度相应减少[21]。在土壤剖面上,由于牲畜践踏作用随着放牧强度增加,地上生物量也因此减少,增加了表层土壤水分蒸发量,土壤表层缺水,形成向地面的水分梯度、根系层水分通过毛管作用向土壤表层输送水分含量大于牧草因为蒸腾作用减弱而节约的水分量,因此随着放牧强度的增加根系分布层水分含量呈现降低的趋势[21]。
一般认为,随着放牧强度的增加,牲畜对土壤的压实作用愈来愈强烈,土壤容重也逐渐增加。Holt等[28]研究报道,土壤容重在强的放牧压力干扰下显著高于低的放牧压力。玉树隆宝滩地区的放牧强度下土壤容重的研究发现,土壤容重在封育区最低,重牧区最高,表明土壤容重随放牧强度的增加而增加。内蒙古典型草原[10]和三江源区[29]、小嵩草为主的高寒草甸[8]的研究均表明,放牧干扰下,土壤容重的变化具有累积效应;土壤容重随着放牧压力的增加而逐渐增大,且随着土壤深度的增加而逐渐增加,本研究的结果与上述研究结果一致,也与Greenwood 等[30]的结果一致,随放牧强度的增大,牲畜践踏作用的增强,土壤孔隙分布的空间格局发生变化,土壤的总孔隙随之减少,土壤容重也因此增加,但是由于该研究区草地表层草毡层较厚,牲畜的践踏作用可能对草毡层下的土壤有一定的影响,但是对根系层土壤容重的影响可能较小。
土壤中的氮是植物生长发育不可或缺的重要养分之一,土壤中可利用氮主要受土壤矿化作用、植物吸收量以及家畜的排泄物量等的影响。Frank等[31]、Han 等[32]和Pei 等[14]的研究认为,随着放牧强度的增加,土壤中全氮含量会逐渐降低,张伟华[27]的研究也得出相同的结论。Abril 和Bucher[33]对阿根廷草原的放牧研究发现,随着放牧强度的增加,土壤全氮含量从20 年未放牧草地的0.28%降低到极端过度放牧草地的0.14%。本研究对玉树隆宝滩地区高寒草甸的放牧试验也得出与上述学者一致的结论,而轻牧区土壤全氮含量最高可能说明适度放牧刺激了植被地上部分的生长,进而促进了地下根系的发育,提高了根向地下分泌的微生物碳即可利用碳,可利用碳相对氮充足时,氮的固定潜能就相对较高。
土壤有机质(主要指土壤碳素)是陆地生物圈生物地球化学循环的主要成分之一,是指示土壤健康的关键指标[34]。土壤有机质是最大的有机碳库,占整个系统有机碳的90%左右,且土壤有机质是植物养分元素循环中心,在土壤中是一个关键因子。一些研究认为,放牧增加了土壤有机质含量[35],对玉树隆宝滩高寒草甸的研究表明,适度放牧的轻牧区土壤有机碳含量显著高于放牧强度较强的中牧区和重牧区。植物凋落物由于适度的牲畜践踏作用而破碎,并与土壤充分接触从而被土壤吸收;另外,放牧导致凋落物的积累量相对减少,有助于凋落物的暴露和充分分解,也有助于碳和养分元素更多地转移到土壤中[36]。而本研究表明,随着放牧强度的逐渐增加,土壤有机碳含量有所减少,这与裴海昆[37]对高寒草甸草原不同放牧强度对土壤有机碳变化的研究结果一致,而对我国东北羊草草原[9]、内蒙古草甸草原[32]的研究也认为土壤有机碳含量随放牧强度的增加而逐渐降低。这是由于放牧干扰改变了地表覆盖状况,影响了土壤温湿度和理化性质,也就对土壤呼吸产生了影响,这可能改变整个高寒草甸的碳循环,进一步对高寒草甸生态系统碳的源汇效应产生影响。过度的放牧使高寒草甸土壤有效养分通过牧草-家畜而流出土壤系统,减少了养分的归还量,加速了整个草地养分的失调过程。草地也因土壤有效养分不能及时补充严重退化,最终导致植被和土壤的双重退化[38]。
4 结论
在适度放牧条件的轻度放牧和中度放牧制度下,草地地上地下生物量均显著高于对照区。封育区由于从封育管理起没有受到放牧干扰,其积累的地上地下生物量最高。
玉树隆宝滩地区高寒草甸表层土壤湿度呈过饱和状态,但随着放牧强度的增加而减少,土壤湿度在各土层上从大到小依次为封育区>轻牧区>中牧区>重牧区。
土壤容重在封育区最低,重牧区最高,表明土壤容重随放牧强度的提高而增加。放牧干扰下,土壤容重的变化具有累积效应;土壤容重随着放牧压力的增加而逐渐增大,且随着土壤深度的增加而逐渐增加。
随着放牧强度的增加,土壤中全氮含量会逐渐降低,且轻牧区土壤全氮含量最高可能说明适度放牧刺激了植被地上部分的生长,进而增加了地下根系的发育,提高氮的固定潜能。对玉树隆宝滩高寒草甸的研究表明,适度放牧的轻牧区土壤有机碳含量显著高于放牧强度较强的中牧区和重牧区,随着放牧强度的逐渐增加,土壤有机碳含量有所减少。
致谢:感谢青海省科技项目(2012-N-529)的资助。感谢青海省玉树州气象局相关人员在野外采样中所给予的大力支持。
[1] Scurlock J M,Hall D O.The global carbon sink:A grassland perspective[J].Global Change Biology,1998,4:229-233.
[2] Conant R T,Paustian K,Elliott E T.Grassland management and conversion into grassland:Effects on soil carbon[J].Ecological Applications,2001,11:343-355.
[3] 李凌浩.土地利用变化对草地生态系统土壤碳贮量的影响[J].植物生态学报,1998,22(4):300-302.
[4] Briske D D,Boutton T W,Wang Z.Contribution of flexible allocation priorities to herbivory tolerance in C4perennial grasses:An evaluation with C labeling[J].Oecologia,1996,105(2):151-159.
[5] Kielland K,Bryant J P.Moose herbivory in taiga:Effects on biogeochemistry and vegetation dynamics in primary succession[J].Oikos,1998,82:377-383.
[6] Ritchie M E,ilman D,Knops J M.Herbivore effects on plant and nitrogen dynamics in oak savanna[J].Ecology,1998,79:165-177.
[7] Krzic M,Broersma K,Thompson D J,Bomke A A.Soil properties and species diversity of grazed crested wheatgrass and native rangelands[J].Journal of Range Management,2000,53(3):353-358.
[8] 王长庭,王启兰,景增春,冯秉福,杜岩功,龙瑞军,曹广民.不同放牧梯度下高寒小嵩草草甸植被根系和土壤理化特征的变化[J].草业学报,2008,17(5):9-15.
[9] 王玉辉,何兴元,周广胜.放牧强度对羊草草原的影响[J].草地学报,2002,10(1):45-49.
[10] 关世英,文沛钦,康师安.不同牧压强度对草地土壤养分含量的影响[A].草原生态系统研究(第五集)[M].北京:科学出版社,1997:17-22.
[11] 安慧,徐坤.放牧干扰对荒漠草原土壤性状的影响[J].草业学报,2013,22(4):35-42.
[12] Li L,Chen Z.Changs in soil carbon storage due to over-grazing in Leymus chinensis steppe in the Xilin river basin of Inner Mongolia[J].Journal of Environmental Science,1997,9(4):486-490.
[13] 傅华,陈亚明,周志宇.阿拉善荒漠草地恢复初期植被与土壤环境的变化[J].中国沙漠,2003,23(6):661-664.
[14] Pei S F,Fu H,Wan C G. Changes in soil properties and vegetation following exclosure and grazing in degraded Alxa desert steppe of Inner Mongolia,China[J].Agriculture,Ecosystems & Environment,2008,124(1):33-39.
[15] 王长庭,龙瑞军,王启兰.放牧扰动下高寒草甸植物多样性,生产力对土壤养分条件变化的响应[J].生态学报,2008,28(9):4144-4152.
[16] 戎郁萍,韩建国,王培,毛培胜.放牧强度对草地土壤理化性质的影响[J].中国草地,2001,23(4):41-47.
[17] Trumbore S E,Davidson E A,Barbosa D P,Daniel C,Nepstad L A. Belowground cycling of carbon in forests and pastures of eastern Amazonia[J].Global Biogeochemical Cycles,1995,9:512-528.
[18] 韦志刚,黄荣辉,董文杰.青藏高原气温和降水的年际和年代际变化[J].大气科学,2003,27(12):157-170.
[19] 周宁芳,秦宁生,屠其璞,李栋梁.近50 年青藏高原地面气温变化的区域特征分析[J].高原气象,2005,24(3):344-349.
[20] 李栋梁,钟海玲,吴青柏,张拥军,候依玲,汤懋苍.青藏高原地表温度的变化分析[J].高原气象,2005,24(3):291-298.
[21] 周秉荣,李凤霞,颜亮东,才仁扎西,耿晓平.高寒沼泽湿地土壤湿度对放牧强度的响应[J].草业科学,2008,25(11):75-78.
[22] 马宗泰,李凤霞,李甫,肖建设.青海玉树隆宝地区生态环境动态变化研究[J].草业科学,2009,26(7):6-11.
[23] 陈佐忠,汪诗平.中国典型草原生态系统[M].北京:科学出版社,2000:125-156.
[24] 汪诗平,王艳芬,李永宏,陈佐忠.不同放牧率对草原牧草再生性能和地上净初级生产力的影响[J].草地学报,1998,6(4):275-281.
[25] 周华坤,赵新全,汪诗平,赵亮,徐世晓.青藏高原高寒灌丛植被对长期放牧强度试验的响应特征[J].西北植物学报,2008,28(10):2080-2093.
[26] 朱绍宏.牦牛放牧强度对高寒草原植被群落及土壤养分影响的研究[D].兰州:甘肃农业大学,2006.
[27] 张伟华,关世英,李跃进.不同牧压强度对草原土壤水分、养分及其地上生物量的影响[J].干旱区资源与环境,2000,14(4):61-64.
[28] Holt J A,Bristow K L,McIvor J G.The effects of grazing pressure in soil animals and hydraulic properties of two soils in semiarid tropical Queensland[J].Australia Journal of Soil Research,1996,34:69-79.
[29] 孙飞达,龙瑞军,蒋文兰,郭正刚,聂学敏.三江源不同鼠洞密度下高寒草甸植物群落生物量和土壤容重特性研究[J].草业学报,2008,17(5):111-116.
[30] Greenwood K L,Macleod D A,Hutchinson K J.Long-term stocking rate effects on soil physical properties[J].Australia Journal of Experimental Agriculture,1997,37:413-419.
[31] Frank A B,Tanaka D L,Hofmann L,Follett R F.Soil carbon and nitrogen of Northern Great Plains grasslands as influenced by long-term grazing[J].Journal of Range Management,1995,48(5):470-474.
[32] Han G D,Hao X Y,Zhao M L,Wang M J,Ellert B H,Willms W,Wang M J.Effect of grazing intensity on carbon and nitrogen in soil and vegetation in a meadow steppe in Inner Mongolia[J].Agriculture,Ecosystems & Environment,2008,125:21-32.
[33] Abril A,Bucher E H.Overgrazing and soil carbon dynamics in the western Chaco of Argentina[J].Applied Soil Ecology,2001,16:243-249.
[34] Percival H J,Parfitt R L,Scott N A.Factors controlling soil carbon levels in New Zealand grassland:Is clay content important[J].Soil Science Society of America Journal,2000,64:1623-1630.
[35] Reeder J D,Schuman G E.Influence of livestock grazing on C sequestration in semi-arid mixed-grass and shot-grass rangelands[J].Enviromental Pollution,2002,116:457-463.
[36] Naeth M A,Bailey A W,Pluth D J,Chanasyk D S,Hardin R T.Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta[J].Journal of Range Management,1991,44(1):7-12.
[37] 裴海昆.不同放牧强度对土壤养分及质地的影响[J].青海大学学报(自然科学版),2004,22(4):29-31.
[38] 高英志,韩兴国,汪诗平.放牧对草原土壤的影响[J].生态学报,2004,24(4):790-797.
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
天津农林科技(2022年2期)2022-04-19 10:48:14
河南畜牧兽医(2022年3期)2022-04-13 09:12:58
绿色科技(2021年10期)2021-06-23 03:13:06
天津农林科技(2020年3期)2020-08-13 03:17:10
电子制作(2019年15期)2019-08-27 01:12:12
活力(2019年21期)2019-04-01 12:17:10
草业学报(2019年2期)2019-02-25 01:57:16
