
RHDV VLPs对口蹄疫病毒B细胞表位的展示效果
2015-03-26 10:57宋艳华范志宇魏后军薛家宾
江苏农业学报 2015年6期
胡 波, 盛 蓉,2, 宋艳华, 范志宇, 魏后军, 薛家宾, 王 芳
(1.江苏省农业科学院兽医研究所/农业部兽用生物制品工程技术重点实验室/国家兽用生物制品工程技术研究中心,江苏南京 210014;2.南京农业大学动物医学院,江苏 南京 210095)
兔出血症(Rabbit hemorrhagic disease,RHD)是由兔出血症病毒(Rabbit hemorrhagic disease virus,RHDV)引起的一种具有高度传染性、高发病率、高致死性的疾病,主要发生于2月龄以上成年兔[1-2]。兔出血症病毒属于杯状病毒属,无囊膜,直径为35~40 nm[3-6]。病毒衣壳由180个衣壳蛋白组成,表面有32个高5~6 nm的圆柱状壳粒,具有嵌杯样病毒典型的杯状结构形态[7]。RHDV衣壳蛋白VP60由579个氨基酸组成,是病毒衣壳组成的最基本单位。
衣壳蛋白VP60是病毒的免疫保护性抗原,与诱导机体抗感染免疫直接相关[8],单独表达VP60蛋白能够自聚成病毒样颗粒(VLPs)。目前研究学者已采用多种异源系统对VP60进行表达,如杆状病毒-昆虫细胞系统[9-10]、酵母表达系统[11]和植物[12]等。通过这些系统表达获得的重组VP60蛋白免疫动物后能够抵抗病毒的感染,提供完全的免疫保护。目前VP60 VLPs作为表位载体和基因转移载体的相关研究结果表明,可在VP60 N、C末端区域以及306~307 aa位插入外源片段,且不破坏病毒样颗粒的形成,为其发展成为携带多表位的载体疫苗提供了依据[6,13-14]。
口蹄疫病毒(Foot-and-mouth disease virus,FMDV)复合多表位可诱导强烈的免疫反应[15-17],O型FMDV至少有5个B细胞表位,其中包括VP1的141~160位(G-H环)和C端200~213位氨基酸残基线性表位,这2个表位是FMDV最重要的抗原位点[18]。本研究将FMDV VP1 2个B细胞表位串联后[GS-(200~213 aa)-GS-(141~160 aa)]插入VP60的C末端(位于VLPs的表面)、N末端(位于VLPs的内部)和306~307 aa之间,构建一系列嵌合VP60蛋白,并将嵌合病毒样颗粒免疫小鼠,检测机体诱导产生针对VP60和FMDV表位的特异性免疫应答,分析VLPs对外源表位的递呈能力,以及外源片段对VLPs的形成及其自身免疫特性的影响,为VP60 VLPs作为外源表位载体展示系统的研究奠定基础。
1 材料与方法
1.1 质粒、菌株、细胞和实验动物
载体pFastBacTMHTA购自Invitrogen公司,质粒pFastBac1-VP60由本实验室构建并保存[8];E.coli DH5α感受态购买自北京全式金生物技术有限公司;E.coli DH10 Bac感受态由本实验室保存;Sf9昆虫细胞由本实验室培养。7~8周龄ICR雌性小鼠,购自扬州大学比较医学中心。
1.2 主要试剂与工具酶
DNA Marker DL2000及DL15000、限制性内切酶(EcoRⅠ、SalⅠ、XbaⅠ、HindⅢ)、T4 DNA连接酶、凝胶回收纯化试剂盒等购自大连宝生物工程有限公司,PfxDNA聚合酶、Taq高保真酶、脂质体LipofectamineTM2000和Grace’s不完全培养基购自invitrogen公司,柱式质粒DNAout试剂盒、柱式动物RNAout试剂盒购自北京天恩泽基因科技有限公司,胎牛血清购自GIBCO公司,酶标兔抗小鼠IgG(FITC-IgG)购自北京鼎国昌盛生物技术有限公司,DAB显色试剂盒购自武汉博士德生物工程有限公司,弗氏完全佐剂和不完全佐剂购自Sigma公司。其他试剂均为国产分析纯级。小鼠抗VP60单抗A3C为本实验室制备保存[19-20]。
1.3 引物设计
1.3.1 设计合成引物 根据GenBank数据库RHDV皖阜株VP60序列(FJ794180),利用Primer 5.0软件设计引物,由Invitrogen公司合成,引物序列见表1。
1.3.2 合成FMDV B细胞表位序列 FMDV VP1 B细胞表位的双串联序列[GS-(411~1 060 aa)-GS-(200~213 aa)-GS],简称FB,由Invitrogen公司合成。序列为:GTCGAC GGCAGCGTACCAAACCTGCGTGGTGACCTGCAGGTACTTGCTCAGAAAGTTGCTCGTACTC TGCCAGGCAGCCGTCACAAACAGGAAATCGTAGCTC CAGTAAAACAGAAGTTGGGCAGCTCTAGA(下划线部分是GS linker,斜体部分是酶切位点)。

表1 试验中所用引物序列Table 1 The sequences of primers used in the study
1.4 重组穿梭载体的构建
1.4.1 重组转移载体的构建 以VP60-F和VP60-1R为引物,质粒pFastBac1-VP60为模板,扩增目的片段,PCR产物经EcoR I和Sal I酶切后克隆至pFastBacTMHTA中,酶切鉴定后获得重组转移载体,命名为pFastBacTMHTA-1;以VP60-NF和VP60-3R为引物,以质粒pFastBac1-VP60为模板,扩增含有Xba I和Hind III酶切位点的PCR产物,并克隆至pFastBacTMHTA中,得到的重组转移载体命名为pFastBacTMHTA-2。以VP60-F和VP60-2R为引物,质粒pFastBac1-VP60为模板,扩增VP60 1~918 bp片段,经EcoR I和Sal I酶切后克隆至pFastBacTMHTA中,得到重组转移载体pFastBacTMHTA-3。此外,以VP60-2F和VP60-3R为引物,扩增获得VP60 918~1 740 bp片段,经Xba I和Hind III酶切后克隆至pFastBacTMHTA-3中,得到重组转移载体,命名为pFastBacTMHTA-4。FMDV(O/China/5/99 strain) B细胞表位[FB(GS-(141-160 aa)-GS-(200-213 aa)-GS)]由人工合成,FMDV-F/FMDV-R经退火处理后形成双链(Sal I/Xba I),用T4 DNA Ligase将Sal I/Xba I酶切消化后的载体(pFastBacTMHTA-1、pFastBacTMHTA-2和pFastBacTMHTA-4)与退火磷酸化处理的FMDV VP1 B细胞表位双链DNA连接,形成3种重组转移载体 pFastBacTMHTA-VP60-2FB、pFastBacTMHTA-VP60-578FB 和 pFastBacTMHTAVP60-306FB。嵌合体结构示意图见图1。
1.4.2 重组穿梭载体的构建 制备E.coli DH10 Bac感受态细胞,分别转化3种重组转移载体,转化产物涂于含有3种抗生素(50 μg/ml卡那霉素、7 μg/ml庆大霉素、10 μg/ml四环素)、100 μg/ml X-gal和20 μg/ml IPTG的LB平板,37℃培养48 h后,挑选白色菌落培养后提取质粒。用M13/PUC通用上下游引物对提取的重组穿梭载体进行PCR鉴定,并进行测序,重组穿梭质粒分别命名为Bacmid-VP60-578FB、Bacmid-VP60-2FB和Bacmid-VP60-306FB。
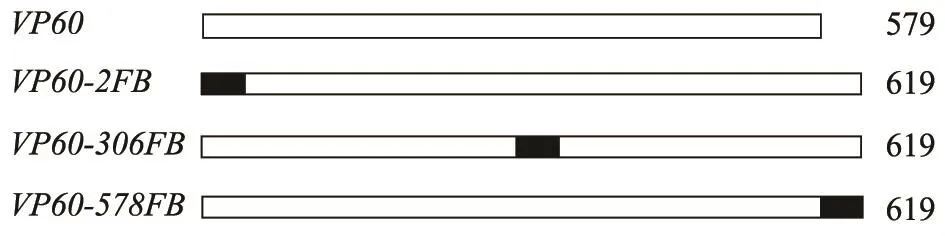
图1 嵌合体VP60-2FB、VP60-306FB和VP60-578FB示意图Fig.1 The schematic diagram of the chimeric constructs (VP60-2FB,VP60-306FB and VP60-578FB)
1.5 VP60-VLPs的表达和鉴定
1.5.1 细胞转染 将鉴定为阳性的重组穿梭载体(Bacmid-VP60-578FB、Bacmid-VP60-2FB和 Bacmid-VP60-306FB)用脂质体LipofectaminTM2000转染至对数生长期的Sf9昆虫细胞。转染后每12 h观察1次,细胞病变明显时收集细胞及上清液,作为重组杆状病毒原液,4℃保存。将获得的重组杆状病毒分别命名为 rAcV-Bac-578FB、rAcV-Bac-306FB和 rAcV-Bac-2FB。将第1代病毒液反复冻融3次后以1%体积比接种Sf9细胞,进行传代,得到第2代重组病毒。
1.5.2 间接免疫荧光检测 3种重组病毒分别感染24孔细胞培养板上对数生长期的Sf9细胞,同时设置空白对照(正常细胞)和阴性对照(感染野生型杆状病毒),本实验室构建的rAcV-Bac-VP60作为阳性对照。在感染Sf9细胞24 h后,加入预冷的乙醇固定液,4℃作用1 h;PBS洗涤3次,分别以1∶200稀释的RHDV单抗A3C和1∶100稀释的牛“O”型FMDV多抗血清为一抗,37℃孵育细胞1 h;随后用PBS反复洗涤3次,分别对应加入FITC标记的羊抗鼠IgG和FITC标记的羊抗牛IgG为二抗,37℃作用1 h。多次充分洗涤,防止留有非特异性荧光,在荧光显微镜下观察。
1.5.3 SDS-PAGE和Western blot分析 收集感染重组病毒的细胞培养物,冻融处理后加入上样缓冲液进行蛋白质电泳(SDS-PAGE),电泳结束后,将凝胶转印NC膜,封闭后,分别以1∶200稀释的RHDV单抗A3C和1:100稀释的牛源“O”型FMDV多抗血清为一抗,分别对应以HRP标记的兔抗鼠IgG和HPR标记的羊抗牛IgG为二抗,最后DAB显色。
1.6 电镜观察
分别将重组病毒接种Sf9细胞,细胞明显病变后收集细胞培养物,反复冻融后离心取上清,经超滤管浓缩,6 000 r/min离心7 min,浓缩10倍后备用。将待检样品滴于载样铜网上,吸附作用2 min;将2%的磷钨酸染液滴于铜网上,固定2 min;最后除去多余的磷钨酸染液,室温干燥5 min,于H-7650型透射电镜上进行观察。
1.7 嵌合蛋白免疫特性研究
1.7.1 抗原的制备 3种重组杆状病毒分别接种对数生长期的Sf9细胞,出现明显病变后,用灭菌PBS(pH 7.4)重悬细胞。反复冻融3次,离心去除细胞碎片,取上清经超滤管纯化浓缩,用无菌PBS将嵌合蛋白稀释为400 μg/ml。按抗原、弗氏佐剂1∶1体积比混合,乳化后备用。1.7.2 动物免疫 7-8周龄雌性ICR小鼠共36只,随机分成6组,分别经腹腔注射乳化后的嵌合蛋白VP60-2FB、VP60-306FB、VP60-578FB、VP60、FMDV灭活疫苗和PBS,每只200 μl(40 μg)。共免疫3次,每次免疫间隔2周,首次免疫采用弗氏完全佐剂,加强免疫采用弗氏不完全佐剂。其中PBS组为阴性对照,VP60组为检测VP60特异性抗体的阳性对照,FMDV组为检测外源B细胞表位应答的阳性对照。于首免后第0、1、2、3、4、5和6周对小鼠进行断尾采血,分离血清,采用本实验室建立的检测VP60特异性抗体水平的间接ELISA方法[21],检测各组免疫小鼠诱导产生VP60特异性抗体的水平;利用O型FMDV ELISA检测试剂盒(购自武汉科前生物制品有限责任公司)检测FMDV VP1 B细胞表位特异性抗体水平。
2 结果
2.1 目的基因的获得及其鉴定
根据GenBank数据库RHDV皖阜株VP60序列(FJ794180),设计引物,以pFastBac1-VP60为模板,进行PCR扩增,PCR产物经1%琼脂糖凝胶电泳,获得符合目的片段大小的条带(图2)。

图2 重组VP60基因PCR扩增结果Fig.2 PCR amplification of recombinant VP60 gene
2.2 重组转移载体的鉴定
将酶切处理的目的基因与pFastBacTMHTA进行连接,转化DH5α感受态细胞,提取重组质粒。分别利用限制性内切酶对3种重组阳性质粒进行酶切鉴定,经琼脂糖凝胶电泳可见2条大小分别为4 775 bp和1 800 bp左右的条带(图3),证明目的片段已成功克隆至载体pFastBacTMHTA中。
2.3 重组穿梭载体的鉴定
利用pUC/M13上、下游引物对重组Bacmid质粒进行PCR扩增,产物大小约为4 000 bp,而以同样引物对空Bacmid质粒进行扩增,产物大小约为300 bp;以目的基因上、下游引物对重组Bacmid质粒进行PCR扩增,获得大小为1 800 bp左右的条带,证明转座成功。
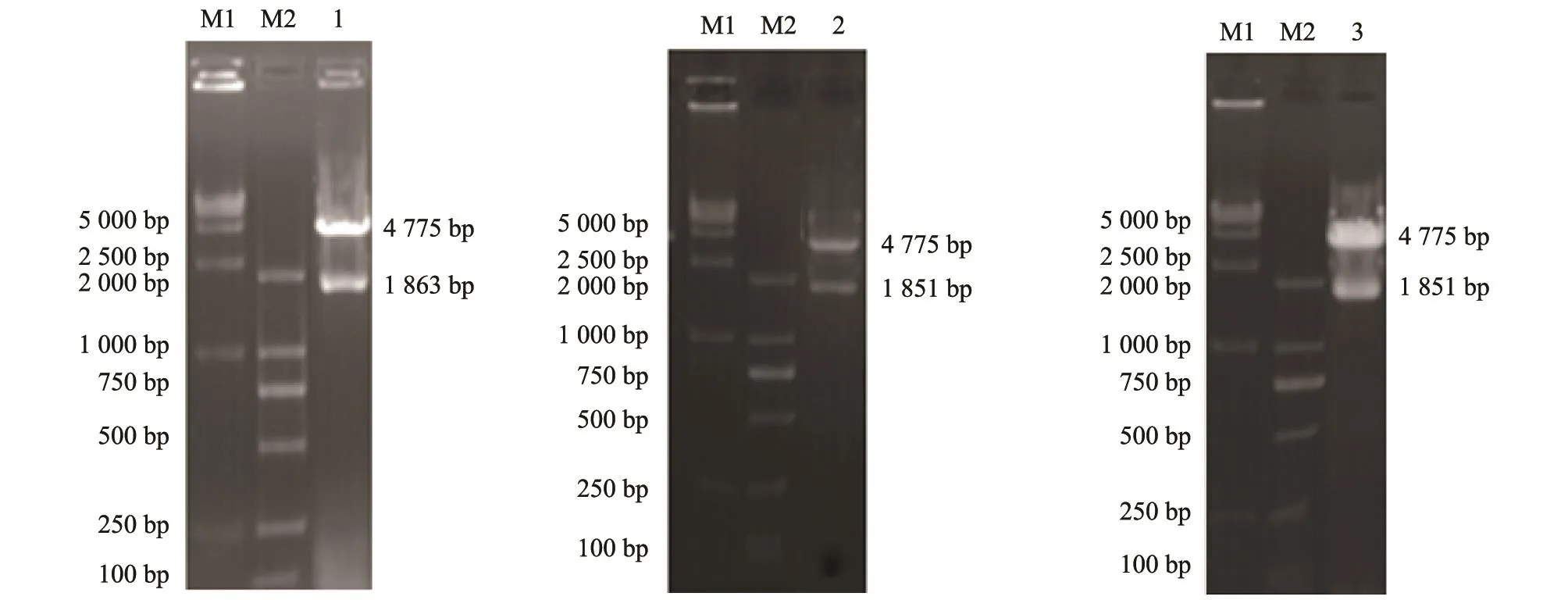
图3 重组质粒的酶切鉴定Fig.3 Identification of the recombinant plasmid by restriction endonuclease digestion
2.4 嵌合蛋白的表达和鉴定
将重 组 病 毒 rAcV-Bac-578FB、rAcV-Bac-306FB、rAcV-Bac-2FB和rAcV-Bac-VP60分别感染Sf9细胞,其中rAcV-Bac-VP60为阳性对照。感染24 h后,以RHDV单抗A3C为一抗,FITC标记兔抗鼠IgG为二抗,进行荧光染色。结果(图4)表明,感染重组杆状病毒的Sf9细胞具有很强的特异性荧光,而感染野生型杆状病毒的Sf9细胞和空白对照细胞无荧光,说明嵌合蛋白得到有效表达。

图4 间接免疫荧光试验(IFA)检测3种嵌合VP60蛋白的表达Fig.4 Indirect immunofluorescence assay(IFA)analysis of the expression of chimeric VP60 proteins
将重组病毒 rAcV-Bac-578FB、rAcV-Bac-306FB、rAcV-Bac-2FB和rAcV-Bac-VP60分别感染Sf9细胞,其中rAcV-Bac-VP60为阳性对照。感染24 h后,以牛FMDV多抗血清为一抗,FITC标记羊抗牛IgG为二抗,进行荧光染色。结果(图5)表明,分别感染 rAcV-Bac-578FB、rAcV-Bac-306FB和rAcV-Bac-2FB的Sf9细胞具有很强的特异性荧光,而感染AcV-Bac-VP60、野生型杆状病毒的Sf9细胞和空白对照细胞无荧光,说明嵌合蛋白携带的VP1 B细胞表位获得有效的表达。
SDS-PAGE和Western blot结果(图6)显示:以VP60单抗A3C为一抗的NC膜在约6.0×104处出现目的条带,与预期大小一致,说明嵌合蛋白得到有效表达;以FMDV牛多抗血清为一抗的NC膜同样在6.0×104处出现目的条带,与预期大小一致,说明FMDV B细胞表位得到有效表达,且免疫印迹与电泳的目的条带位置相符。

图5 IFA检测3种嵌合蛋白中B细胞表位的表达Fig.5 IFA analysis of the expression of B cell epitope in chimeric proteins
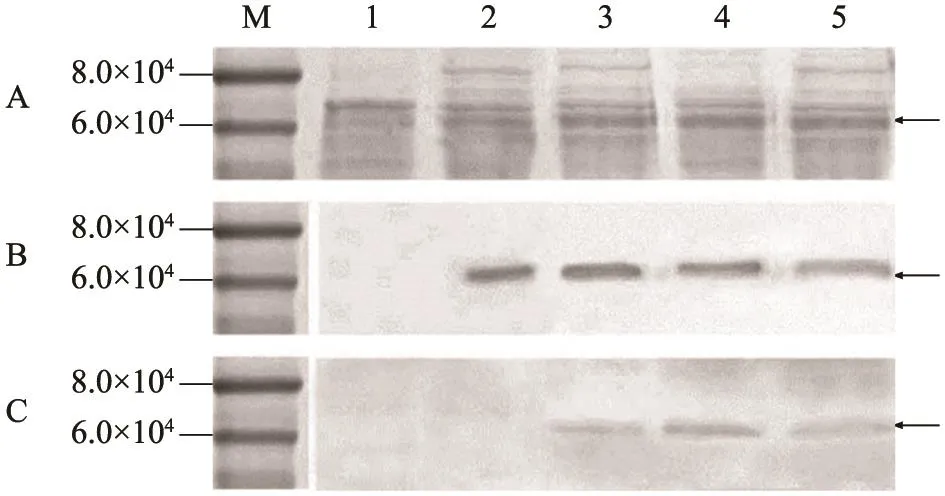
图6 3种嵌合蛋白的SDS-PAGE和Western blot分析Fig.6 SDS-PAGE and Western blot analysis of the chimeric proteins
2.5 电镜观察结果
电镜观察结果(图7)显示,本试验构建并表达的3种嵌合蛋白均可形成大小约为40 nm的VLPs,形态和大小均与天然RHDV VLPs的粒子(阳性对照)相似,说明外源表位的插入未影响VP60自身VLPs的组装。
2.6 嵌合蛋白诱导小鼠的体液免疫应答
为检测嵌合蛋白VP60刺激免疫系统产生的体液免疫反应,本研究用间接ELISA检测方法测定小鼠血清中抗载体VP60的特异性抗体水平和抗外源FMDV B细胞表位的特异性抗体水平。实验小鼠分为VP60-2FB组、VP60-306FB组和VP60-578FB组,同时设立VP60组和FMDV灭活疫苗组作为阳性对照,PBS组作为阴性对照。分别于首免后0、1、2、3、4、5、6周对各组小鼠进行尾静脉采血,分离血清进行检测。结果显示:首免后4-6周,3种嵌合蛋白均可产生较强的VP60特异性抗体,与PBS组比较差异显著(P<0.05)(图8);3种嵌合蛋白刺激机体产生的表位特异性IgG抗体与VP60组相比差异显著(P<0.05),VP60组未产生表位特异性抗体应答,3种嵌合蛋白均可诱导产生较强的表位特异性抗体应答水平(图9)。此外,与嵌合VP60组相比,作为阳性对照的FMDV商品化灭活苗组诱导产生的表位特异性IgG抗体水平显著高于嵌合VP60组(P<0.05)(图9)。
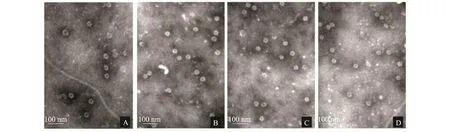
图7 RHDV VP60蛋白和3种嵌合蛋白病毒样粒子的电镜观察结果Fig.7 RHDV VP60 protein and chimeric VP60 VLPs under electron microscopy

图8 间接ELISA检测小鼠血清中VP60特异性抗体水平Fig.8 The detection of anti-VP60 antibody level in the serum of mice by indirect ELISA
3 讨论
杯状病毒成球形或近球形,无囊膜,核衣壳呈二十面体对称,由180个单一结构蛋白组成,病毒核酸为单股、正链线性RNA(ssRNA)[22-26]。杯状病毒的VLPs结构简单,由单一衣壳蛋白自行装配而成,是一种研究VLPs的理想模型。很多病毒的宿主范围很广,不仅可以感染人类,而且可以感染多种动物,这就为VLPs作为疫苗和药物的应用带来了机遇。开发一种多功能的VLPs展示平台,从而可以避免体内预先的中和抗体反应对VLPs所介导的免疫应答的限制,具有重要的意义。RHDV宿主专一性较强,只对兔易感,对人和其他动物不存在安全威胁; RHDV作为杯状病毒的成员之一,其VP60可自行装配形成VLPs,使RHDV VP60作为研究其他动物疾病甚至是人类疾病VLPs疫苗的研究模型成为可能,但其作为展示系统的可行性、效果及其适用范围还需要进一步研究。
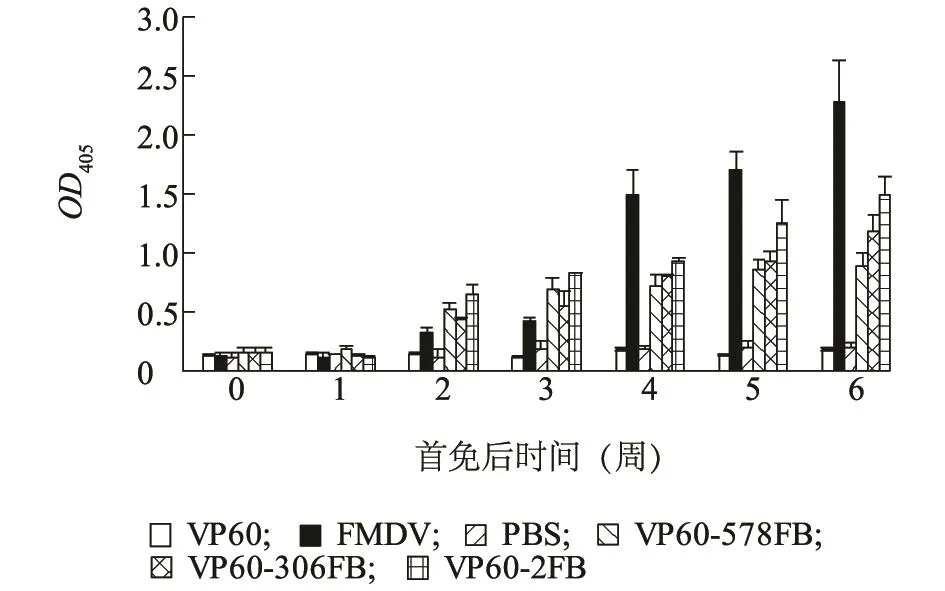
图9 间接ELISA检测FMDV B细胞表位抗体水平Fig.9 The detection of anti-FMDV B cell epitope antibody level in the serum of mice by indirect ELISA
本研究通过插入外源表位(FMDV VP1 B细胞表位)构建VP60嵌合蛋白,研究VP60作为载体递呈外源表位的能力,分析外源表位的插入是否影响其自身病毒样颗粒的形成,评价VP60作为外源表位展示系统的可行性。研究结果表明,3种嵌合蛋白均能够有效表达并形成VLPs,说明在VP60蛋白的第306~307 aa、C端和N端插入42 aa外源片段均不影响VLPs的组装,具备自我装配能力是VLPs疫苗候选物的基本特性。
选择口蹄疫病毒两个重要的B细胞表位GS-(200~213 aa)-GS-(141-160 aa)作为外源插入表位,构建VP60嵌合VLPs。通过IFA和Western blot试验发现VP60嵌合VLPs获得有效表达,并具有VP60特异性和FMDV表位特异性抗原反应。动物试验结果显示,3种嵌合蛋白免疫的小鼠均能够诱导产生强烈的VP60特异性和表位特异性体液免疫应答。综上所述,VP60 VLPs能够有效递呈外源表位,刺激机体对外源B细胞表位产生体液免疫应答,可作为异源抗原的展示载体。此外,我们在前期研究的基础上,将单串联的B细胞表位增加成双串联的B细胞表位,外源片段大小增加至126 bp,序列的增加不影响VLPs的形成和免疫原性,可见VP60-VLPs作为递呈外源抗原的展示载体的容耐性较大,可容纳至少42 aa的外源片段。
本研究以VP60-VLPs为基础,通过在VP60 N端、C端和306~307 aa位插入双串联FMDV B细胞表位序列构建嵌合VLPs,研究结果表明VP60的N端、C端和306~307 aa位置均适合外源氨基酸片段插入,嵌合VLPs均能够进行自我组装,且能够有效展示长度为42 aa的外源片段。此外,VP60作为载体所能容纳的最长外源氨基酸片段长度仍需进一步研究。
[1] FERREIRA P G,COSTA-E-SILVA A,MONTEIRO E,et al.Transient decrease in blood heterophils and sustained liver damage caused by calicivirus infection of young rabbits that are naturally resistant to rabbit haemorrhagic disease[J].Res Vet Sci,2004,76(1):83-94.
[2] XU Z J,CHEN W X.Viral haemorrhagic disease in rabbits:a review[J].Vet Res Commun,1989,13(3):205-212.
[3] ABRANTES J,VAN DER LOO W,LE PENDU J,et al.Rabbit haemorrhagic disease(RHD)and rabbit haemorrhagic disease virus(RHDV):a review[J].Vet Res,2012,10(43):12.
[4] COOKE B D.Rabbit haemorrhagic disease:field epidemiology and the management of wild rabbit populations[J].Rev Sci Tech,2002,21(2):347-358.
[5] CHEN M,SONG Y,FAN Z,et al.Immunogenicity of different recombinant rabbit hemorrhagic disease virus-like particles carrying CD8+T cell epitope from chicken ovalbumin(OVA)[J].Virus Res,2014,183:15-22.
[6] CRISCI E,ALMANZA H,MENA I,et al.Chimeric calicivirus-like particles elicit protective anti-viral cytotoxic responses without adjuvant[J].Virology,2009,387(2):303-312.
[7] VALICEK L,SMID B,RODAK L,et al.Electron and immunoelectron microscopy of rabbit haemorrhagic disease virus(RHDV)[J].Arch Virol,1990,112(3-4):271-275.
[8] 金明兰,侯继波,郑其升,等.兔出血症病毒VP60蛋白T细胞表位优势区的筛选[J].江苏农业科学,2013,41(4): 186-188.
[9] 王 芳,胡 波,任雪枫,等.兔出血症病毒衣壳蛋白在昆虫细胞中的表达及对家兔的免疫保护效果[J].畜牧兽医学报,2008,39(10):1382-1387.
[10]PLANA-DURAN J,BASTONS M,RODRIGUEZ M J,et al.Oral immunization of rabbits with VP60 particles confers protection against rabbit hemorrhagic disease[J].Arch Virol,1996,141(8): 1423-1436.
[11]FARNOS O,RODRIGUEZ M,CHIONG M,et al.The recombinant rabbit hemorrhagic disease virus VP60 protein obtained from Pichia pastoris induces a strong humoral and cell-mediated immune response following intranasal immunization in mice[J].Vet Microbiol,2006,114(3-4):187-195.
[12]FERNANDEZ-FERNANDEZ M R,MOURINO M,RIVERA J,et al.Protection of rabbits against rabbit hemorrhagic disease virus by immunization with the VP60 protein expressed in plants with a potyvirus-based vector[J].Virology,2001,280(2):283-291.
[13]LAURENT S,KUT E,REMY-DELAUNAY S,et al.Folding of the rabbit hemorrhagic disease virus capsid protein and delineation of N-terminal domains dispensable for assembly[J].Arch Virol,2002,147(8):1559-1571.
[14]BARCENA J,VERDAGUER N,ROCA R,et al.The coat protein of rabbit hemorrhagic disease virus contains a molecular switch at the N-terminal region facing the inner surface of the capsid[J].Virology,2004,322(1):118-134.
[15]DIMARCHI R,BROOKE G,GALE C,et al.Protection of cattle against foot-and-mouth disease by a synthetic peptide[J].Science,1986,232(4750):639-641.
[16]BAXTER R,CRAIGMILE S C,HALEY C,et al.BoLA-DR peptide binding pockets are fundamental for foot-and-mouth disease virus vaccine design in cattle[J].Vaccine,2009,28(1):28-37.
[17]ZHANG H Y,SUN S H,GUO Y J,et al.Immune response in mice inoculated with plasmid DNAs containing multiple-epitopes of foot-and-mouth disease virus[J].Vaccine,2003,21(32): 4704-4707.
[18]WONG H T,CHENG S C,CHAN E W,et al.Plasmids encoding foot-and-mouth disease virus VP1 epitopes elicited immune responses in mice and swine and protected swine against viral infection[J].Virology,2000,278(1):27-35.
[19]蔡少平,王 芳,贾华敏,等.兔出血症病毒胶体金免疫层析试纸条诊断方法的建立及初步应用[J].畜牧兽医学报,2012,43 (11):1795-1801.
[20]杨廷亚,王 芳,姜 平,等.应用噬菌体展示技术筛选兔出血症病毒抗原模拟表位[J].畜牧兽医学报,2012,43(8): 1281-1286.
[21]李超美,王 芳,蔡少平,等.检测兔出血症病毒抗体间接ELISA方法的建立[J].江苏农业学报,2010,26(3):546-550.
[22]DI MARTINO B,MARSILIO F,ROY P.Assembly of feline calicivirus-like particle and its immunogenicity[J].Vet Microbiol,2007,120(1-2):173-178.
[23]NICOLLIER-JAMOT B,OGIER A,PIROTH L,et al.Recombinant virus-like particles of a norovirus(genogroup II strain)administered intranasally and orally with mucosal adjuvants LT and LT(R192G)in BALB/c mice induce specific humoral and cellular Th1/Th2-like immune responses[J].Vaccine,2004,22(9-10): 1079-1086.
[24]HAN M G,CHEETHAM S,AZEVEDO M,et al.Immune responses to bovine norovirus-like particles with various adjuvants and analysis of protection in gnotobiotic calves[J].Vaccine,2006,24(3):317-326.
[25]SOUZA M,COSTANTINI V,AZEVEDO M S,et al.A human norovirus-like particle vaccine adjuvanted with ISCOM or mLT induces cytokine and antibody responses and protection to the homologous GII.4 human norovirus in a gnotobiotic pig disease model[J].Vaccine,2007,25(50):8448-8459.
[26]PEREZ-FILGUEIRA D M,RESINO-TALAVAN P,CUBILLOS C,et al.Development of a low-cost,insect larvae-derived recombinant subunit vaccine against RHDV[J].Virology,2007,364 (2):422-430.
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
西藏农业科技(2019年3期)2019-11-04
温州医科大学学报(2019年4期)2019-04-28
食品科学(2018年10期)2018-05-23
现代园艺(2018年3期)2018-02-10
上海农业学报(2017年3期)2017-04-10
中国免疫学杂志(2017年1期)2017-01-17
西南医科大学学报(2015年1期)2015-08-22
畜牧兽医学报(2015年3期)2015-07-05
医学研究杂志(2015年6期)2015-07-01
