
黄绿绿僵菌KM1104对灰飞虱的毒力
2015-03-23 07:23杜广祖杨卫秋周跃能和淑琪桂富荣李正跃
植物保护 2015年5期
杜广祖,杨卫秋,周跃能,2,和淑琪,陈 斌*,桂富荣,李正跃
(1.云南农业大学植物保护学院,云南省微生物发酵工程研究中心有限公司,农业生物多样性应用技术研究国家工程中心,昆明 650201;2.云南省河口县植检植保站,河口 661300)
灰飞虱[Laodelphaxstriatellus(Fallén)]属半翅目飞虱科,是水稻上的一种重要害虫,主要为害早、中稻秧田和本田分蘖期的稻苗[1]。国外主要分布于从菲律宾至西伯利亚一带的亚洲稻区和欧洲的温带地区[2],国内则几乎遍及各稻区。除以成虫、若虫刺吸为害水稻、小麦外,还传播水稻条纹叶枯病(Rice stripevirus,RSV)、水稻黑条矮缩病(Riceblackstreakeddwarfvirus,RBSDV)、小麦丛矮病(Northern cerealmosaicvirus,NCMV)等多种植物病毒病[3-4],且其传毒危害造成的损失远大于刺吸危害[5-6]。
绿僵菌作为重要的昆虫病原真菌种类,寄主广泛,对人畜及环境安全,是国内外害虫生防真菌研究中备受关注的虫生真菌种类之一[7],已广泛用于蝗虫的防治实践[8-10],也有发现在田间感染褐飞虱[11]。温度是影响虫生真菌生长发育与孢子萌发的重要因素之一,温度过高或过低均会影响其毒力[12]。目前关于黄绿绿僵菌对褐飞虱的毒力测定已有一些研究报道[13-15],但有关绿僵菌对灰飞虱毒力研究尚未见报道。本研究针对云南省本地灰飞虱发生为害严重的现状,生物测定黄绿绿僵菌在不同温度下对灰飞虱成虫及若虫的室内毒力,以期筛选灰飞虱生防黄绿绿僵菌菌株及其侵染适宜温度条件,从而为灰飞虱生物防治提供理论依据。
1 材料与方法
1.1 试验材料
供试黄绿绿僵菌(M.flavoviride)菌株KM1104分离自昆明水稻田的褐飞虱罹病虫体;采用SDAY 培养基分离纯化后4℃保存于云南农业大学植物保护学院昆虫病原体实验室。
1.2 试验方法
供试昆虫:室内采用张爱民等[16]的方法在人工气候室内用‘楚粳29’水稻苗饲养繁殖10代,获得龄期一致的灰飞虱健康种群。
菌株培养:将供试菌株分别在光照培养箱25 ℃,L∥D=16h∥8h条件下,SDAY培养基扩大培养采集分离获得的菌株,4℃保存于昆虫病原体实验室备用。
毒力测定:在SDAY 培养基上培养,待孢子粉布满全皿后,刮取分生孢子粉移入20mL含0.05%Tween-80湿润剂的无菌水中,置混匀器上充分振荡,然后按比例依次稀释配制1.01×108、1.01×107、1.01×106、1.01×105、1.01×104、1.01×103孢子/mL浓度的孢子悬浮液,以含0.05% Tween-80的无菌水为对照。选取龄期一致的灰飞虱3 龄若虫或成虫,采用浸渍法[17]接种,每次选取30头供试灰飞虱虫体置于200目尼龙纱网袋中再浸入相应浓度的孢子悬浮液2s,然后迅速取出,用灭过菌的干滤纸吸去虫体表面残留的悬浮液,分别放置于24、27、30 ℃,光周期L∥D =12h∥12h,湿度80%±5%的光照培养箱中饲养,每天定时观察记载死亡虫量,并将死亡虫体在25℃和L∥D=12h∥12h条件下保湿培养,鉴定死亡原因,每个处理重复3次。
1.3 数据统计
采用TDM 模型(时间-剂量-死亡率模型)处理生物测定数据,采用Duncan检验分析各数据的差异显著性,所有数据分析采用DPS软件[18]。
2 结果与分析
2.1 接种黄绿绿僵菌KM1104后灰飞虱的累积死亡率
2.1.1 灰飞虱成虫的累积死亡率
从图1(左)可以看出,在24~30 ℃下,黄绿绿僵菌KM1104菌株对灰飞虱成虫具有一定的侵染致病能力,用孢子悬浮液接种灰飞虱成虫后,累积死亡率随接种浓度的增加及处理时间的延长而增加,不同浓度及温度下累积死亡率差异明显。在24、27和30 ℃下,用1.01×103~1.01×108孢子/mL 绿僵菌孢子悬浮液接种灰飞虱成虫后,第4天时的累积死亡率分别为11.11%~35.56%、14.44%~28.89%和17.78%~53.33%,表明在30 ℃下灰飞虱成虫的累积死亡率高于24 ℃(F=21.32,P<0.01)和27 ℃(F=19.45,P<0.01),但24 ℃和27 ℃下的累积死亡率间无明显差异(F=3.47,P>0.015)。到接种后第12 天时,于24、27 和30 ℃下接种了1.01×103~1.01×108孢子/mL 的灰飞虱成虫的累积死亡率分别为26.67%~90.00%、34.44%~94.44%和52.22%~100%。24 和27 ℃下用低浓度1.01×103孢子/mL 接种后,灰飞虱成虫的累积死亡率均低于50%,而在30 ℃下用1.01×103孢子/mL接种后灰飞虱成虫的累积死亡率达50%以上,明显高于24 ℃(F=31.45,P<0.01)和27 ℃(F=23.39,P<0.01)下的死亡率。
2.1.2 灰飞虱若虫的累积死亡率
从接种处理后灰飞虱若虫的累积死亡率(图1:右)可以看出,在24~30 ℃下,黄绿绿僵菌对灰飞虱3龄若虫均具有致病力,其中27 ℃和30 ℃下致病力较强,用浓度为1.01×103~1.01×108孢子/mL孢子悬浮液接种后第4 天时,累积死亡率分别为8.89%~24.44%和10.00%~30.00%,而到第12天时累积死亡率均达50%以上,分别为51.11%~100%和61.11%~100%。在24 ℃下,用浓度为1.01×103~1.01×108孢子/mL 绿僵菌孢子悬浮液接种3龄若虫后第4天时累积死亡率仅为5.56%~20.00%,低于27 ℃和30 ℃下的累积死亡率(27 ℃:F=9.32,P<0.05;30 ℃:F=11.15,P<0.05),用1.01×108孢子/mL 浓度接种后第12天时累积死亡率达72.22%,仍明显低于30 ℃下的累积死亡率(F=37.28,P<0.01)。

图1 不同温度下黄绿绿僵菌KM1104对灰飞虱的累积死亡率Fig.1 The cumulative mortality of Laodelphax striatellus after infection by Metarhizium flavoviride at different temperatures
2.2 黄绿绿僵菌KM1104对灰飞虱成虫和若虫的致死中浓度LC50
从表1 可以看出,不同温度下黄绿绿僵菌KM1104对灰飞虱成虫和3龄若虫侵染致病的致死中浓度不同,其中30 ℃下对灰飞虱成虫和3龄若虫的致死中浓度LC50明显低于24和27 ℃下的LC50(24 ℃:成虫F=21.31,P<0.05;若虫F=25.45,P<0.05;27 ℃:成虫F=15.27,P<0.05;若虫F=19.37,P<0.05)。24℃时,黄绿绿僵菌KM1104对灰飞虱成虫和3龄若虫的LC50分别为7.14×104孢子/mL和3.69×105孢子/mL,与27 ℃下成虫和若虫的LC50间无明显差异(成虫:F=2.24,P>0.05;若虫:F=1.19,P>0.05),但显著大于30 ℃下对成虫和3龄若虫的LC50(成虫:F=15.34,P<0.05;若虫:F=16.41,P<0.05),因而在24 ℃下的致病力低于27 ℃和30 ℃下的致病力。
此外,从表1还可看出,在24~30 ℃下,黄绿绿僵菌KM1104 菌株对灰飞虱成虫的致死中浓度LC50均小于对3龄若虫的LC50,表明对成虫的致病力强于若虫。如在27 ℃时,对成虫的LC50为1.33×104孢子/mL,显著低于对3 龄若虫的LC50值1.05×105孢子/mL(F=36.43,P<0.01);同样,在30 ℃时对成虫的LC50小于对3龄若虫的LC50(F=29.27,P<0.01)。

表1 不同温度下黄绿绿僵菌KM1104对灰飞虱的毒力1)Table1 Virulence of Metarhizium flavoviride to Laodelphax striatellus at different temperatures
综合以上结果,黄绿绿僵菌KM1104对灰飞虱成虫的致病力高于对3龄若虫的致病力,不同温度下的致病力大小顺序为30 ℃>27 ℃>24 ℃,对成虫和3龄若虫的LC50分别为1.45×103~7.14×104和7.25×103~3.69×105孢子/mL。
2.3 黄绿绿僵菌KM1104对灰飞虱的致死中时LT50
黄绿绿僵菌菌株KM1104对灰飞虱成虫和3龄若虫的侵染致死中时LT50不同(表2)。用较低浓度黄绿绿僵菌菌株1.01×105和1.01×106孢子/mL于24、27和30 ℃下接种处理灰飞虱成虫和3龄若虫的LT50分别为5.13~11.53和8.49~11.58d,总体表现为在该浓度下黄绿绿僵菌更易于侵染灰飞虱成虫(F=18.35,P<0.01)。当用较高浓度的1.01×107和1.01×108孢子/mL 接种后对成虫和3龄若虫的LT50分别为3.02~7.01d 和6.85~10.05d,即表现对成虫的致病速度明显快于若虫。综上所述,供试黄绿绿僵菌KM1104菌株对灰飞虱成虫的侵染致病速度比若虫快。
不同温度下黄绿绿僵菌KM1104菌株对灰飞虱成虫和3龄若虫的侵染致死中时LT50不同(表2),24和27 ℃下较高接种浓度对成虫的LT50间无明显差异,30 ℃下对成虫的LT50明显小于对3龄若虫的LT50值,因此在30 ℃下,该菌株对灰飞虱成虫的侵染致病力强于3 龄若虫,接种1.01×105~1.01×108孢子/mL 后对成虫和3 龄若虫的LT50分别为3.02~6.86d和6.85~9.35d。
3 讨论
黄绿绿僵菌作为一种传统的虫生真菌,一直是害虫生防真菌筛选和开发研究的重要种类之一[7,19],不同菌株的生长温度范围不同,如黄绿绿僵菌菌株SM076能在20~29 ℃温度范围内生长发育,其最适生长发育温度为26 ℃[20]。本研究发现黄绿绿僵菌KM1104 菌株在24、27 和30 ℃下对灰飞虱成虫和3龄若虫均具有一定的致病作用,但在27 ℃和30 ℃下的致病力高于24 ℃,这也与灰飞虱最适生长、繁殖的温度范围一致[1-21],因而该菌株KM1104是一株对灰飞虱具有良好生防潜力的菌株。然而耿博闻研究发现在25 ℃条件下黄绿绿僵菌对褐飞虱成虫的侵染致死率较高[22],造成这种差异的原因可能与菌株来源和昆虫种类不同有关。
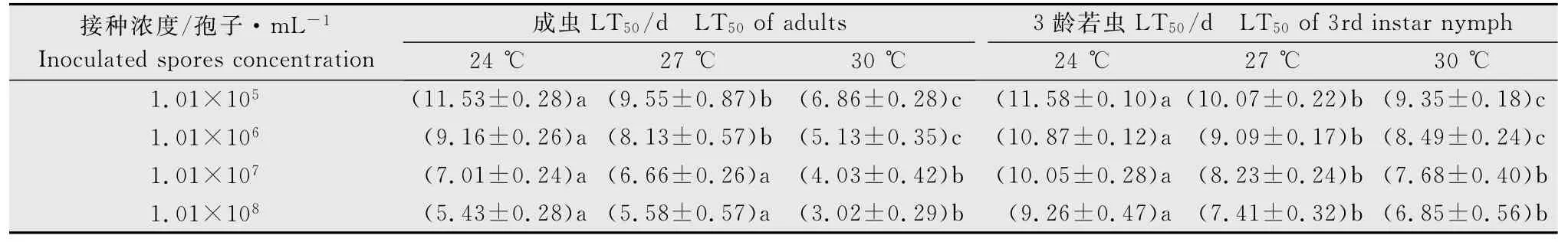
表2 不同温度下黄绿绿僵菌对灰飞虱成虫及3龄若虫的致死中时LT501)Table 2 The median lethal time(LT50)of Metarhizium flavoviride to adults and 3rd instar nymph of Laodelphax striatellus at different temperatures
同时,本研究发现黄绿绿僵菌菌株KM1104对灰飞虱成虫及3龄若虫均有致病力,但对成虫的致病力高于3龄若虫,30 ℃时,接种浓度为1.01×108孢子/mL时对灰飞虱成虫的致死中时LT50(3.02d)显著低于3龄若虫(6.85d),表明灰飞虱成虫对黄绿绿僵菌的敏感性高于3龄若虫,该结果与李茂业等报道的黄绿绿僵菌Mf82菌株对褐飞虱成虫的致病性高于5 龄若虫的结果一致[14]。由于黄绿绿僵菌主要在体表以芽管或产生附着胞侵染褐飞虱[13],若虫蜕皮会影响到分生孢子在体表的附着,这也是造成若虫死亡率低于成虫的原因之一。
黄绿绿僵菌对昆虫具有较强的致病性而对其他节肢动物无害[23],已广泛用于害虫生防制剂的开发,本研究中的菌株分离自罹病褐飞虱成虫,说明该菌株在自然条件下可以感染褐飞虱成虫;同时研究表明对灰飞虱成虫及3 龄若虫均存在一定的致病力。因此,当田间2种稻飞虱共同存在时,施用绿僵菌制剂则可对两种稻飞虱都起到良好的控制作用,因而黄绿绿僵菌KM1104可用于稻飞虱生防制剂的开发。
[1]张爱民,刘向东,翟保平,等.温度对灰飞虱生物学特性的影响[J].昆虫学报,2008,51(6):640-645.
[2]Kisimoto R.Flexible diapause response to photoperiod of a laboratory selected line in the small brown planthopper,LaodelphaxstriatellusFallén[J].Applied Entomology and Zoology,1989,24(1):157-159.
[3]Hadidi A,Khetarpal R K,Koganezawa H.Plant virus disease control[M].US:American Phytopathological Society Press,1998.
[4]Kimura I,Murao K,Isogai M.Tissue culture of vector planthopper,Laodelphaxstriatellus(Fallen)ofRiceblack-streaked dwarfvirus(RBSDV),and inoculation of the virus to their cell monolayers[J].Annals of the Phytopathological Society of Japan,1996,62(3):338.
[5]林凌伟,董国堃,汪恩国.水稻黑条矮缩病传毒昆虫的防治实践与研究[J].昆虫知识,2001,38(6):426-428.
[6]朱龙粉,傅华欣,毛华方,等.水稻条纹叶枯病发生规律与防治技术研究[J].植保技术与推广,2003,23(5):3-6.
[7]蒲蛰龙,李增智.昆虫真菌学[M].合肥:安徽科技出版社,1996:93-111.
[8]雷仲仁,问锦曾.绿僵菌治蝗研究进展[J].植物保护,2004,30(4):14-17.
[9]Thomas B,Gbongboui C,Lomer C J.Between season survival of the grasshopper pathogenMetarhiziumflavoviridein the Sahel[J].Biocontrol Science and Technology,1996,6(4):569-573.
[10]Lomer C J,Prior C,Kooyman C.Development ofMetarhiziumspp.for the control of grasshoppers and locusts[J].Memoirs of the Entomological Society of Canada,1997,129(171):265-286.
[11]Rombach M C,Aguda R M,Shepard B M,et al.Infection of rice brown planthopper,Nilaparvatalugens(Homoptera:Delphacidae),by field application of entomopathogenic Hyphomycetes(Deuteromycotina)[J].Environmental Entomology,1986,15(5):1070-1073.
[12]岳梅,雷仲仁,孟涛,等.温度对绿僵菌LA 菌株孢子萌发、酶活力及毒力的影响[J].植物保护,2010,36(2):56-60.
[13]李茂业,林华峰,金立,等.黄绿绿僵菌对褐飞虱侵染过程的扫描电镜观察[J].应用昆虫学报,2011,48(5):1412-1416.
[14]李茂业,林华峰,李世广,等.黄绿绿僵菌Mf82 菌株对不同虫态褐飞虱的毒力[J].昆虫学报,2012,55(3):316-323.
[15]林华峰,张松影,李茂业,等.黄绿绿僵菌Mf82 悬乳剂对褐飞虱作用的时间-剂量-死亡率模型分析[J].菌物学报,2013,32(2):239-247.
[16]张爱民,刘向东.室内饲养灰飞虱的一种简便方法[J].昆虫知识,2008,45(2):314-315.
[17]Feng M G,Johnson J B,Kith L P.Virulence ofVerticilliumlecaniiand an aphid-derived isolate ofBeauveriabassiana(Fungi:Hyphomycetes)for six species of cereal-infesting aphids(Homoptera:Aphididae)[J].Environmental Entomology,1990,19(3):815-820.
[18]唐启义,冯明光.实用统计分析及其DPS 数据处理系统[M].北京:科学出版社,2002:295-304.
[19]Roberts D W,St Leger R J.Metarhiziumspp.,cosmopolitan insect-pathogenic fungi:mycological aspects[J].Advances in Applied Microbiology,2004,54:1-70.
[20]李佳颖,徐奇爱,代晓彦,等.黄绿绿僵菌SM0706菌株生物学特性研究[J].广西农业科学,2013,40(9):71-73.
[21]Hachiya K.Effect of temperature on the developmental velocity of small brown planthopperLaodelphaxstriatellusFallén[J].Annual Report of the Society of Plant Protection of North Japan,1990(41):112-113.
[22]耿博闻.黄绿绿僵菌防治稻飞虱的研究[D].广州:中山大学,2004.
[23]Peveling R,Demba S A.Virulence of the entomopathogenic fungusMetarhiziumflavovirideGams and Rozsypal and toxicity of diflubenzuron,fenitrothion-esfenvalerate and profenofos-cypermethrin to nontarget arthropods in Mauritania[J].Archives of Environmental Contamination and Toxicology,1997,32(1):69-79.
猜你喜欢
湖北植保(2022年5期)2022-12-24
上海农业科技(2022年4期)2022-08-15
环境昆虫学报(2021年6期)2022-01-05
农药科学与管理(2021年10期)2021-12-07
昆虫学报(2021年8期)2021-09-06
ELLE世界时装之苑(2021年4期)2021-04-07
昆虫学报(2020年8期)2020-10-15
浙江农业学报(2019年8期)2019-08-29
小溪流(画刊)(2015年9期)2016-03-03
小雪花·成长指南(2015年5期)2015-05-25
