
内蒙古典型草原蝗虫群落结构和生态位研究
2015-03-23 07:23秦兴虎吴惠惠黄训兵王广君曹广春农向群张泽华
植物保护 2015年5期
秦兴虎,吴惠惠,黄训兵,王广君,曹广春,农向群,张泽华*
(1.中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 100193;2.农业部锡林郭勒草原有害生物科学观测实验站,锡林浩特 026000)
内蒙古草原构成了欧亚大陆温带草原的东翼,其植被具有广泛的代表性。由于全球气候变化和人类活动,使草地退化,草原蝗灾暴发,导致草原生态系统结构和功能失调,群落多样性面临严重的威胁。草原蝗灾给畜牧业和粮食产量造成了巨大损失,因此,深入研究蝗虫的群落结构和生态位对于了解蝗虫在草原生态系统中的结构功能、地位与防控,以及正确评价人类活动和全球变化对物种的影响具有重要的理论意义。
目前,对于蝗虫的群落结构和生态位已有很多研究,康乐[1-2]发现蝗虫的分布和种群结构同植物群落类型和结构密切相关,并研究了11种草原蝗虫多维生态位关系、多样性格局、种间关联以及蝗虫与植物关系。贺达汉等[3]、康乐等[1-2]对蝗虫的营养生态位和时空生态位进行了探讨,发现草原蝗虫的发生与为害受植物群落结构、土壤的湿度、营养、物理结构及理化特性等的影响,但是并没有系统地探讨蝗虫群落结构和生态位对于宜生区的划分和蝗虫利用资源的分类以及蝗虫发生危害的潜在可能性。Grinnell[4-6]在20 世纪一二十年代就利用物种发生数据研究影响其分布的因素,并借此探讨物种空间分布与进化的关系。目前,生态位模型已被广泛应用于物种时空分布格局[7-9]、外来种入侵预警[10-13]、全球变化对物种分布或多样性格局的影响[14-15]等众多研究领域。本文研究了内蒙古草原蝗虫的群落结构和生态位,定量化地确定蝗虫在草原生态系统中的种群地位和空间地位,分析了蝗虫的优势类群和存在的优势资源以及存在的可能潜在危害,为蝗虫宜生区的划分和草原蝗虫发生监测预警与生态治理提供理论依据。
1 材料与方法
1.1 样地选择
试验地位于内蒙古自治区锡林郭勒盟镶黄旗东部地区(113°22′~114°45′E,41°56′~42°45′N),北起呼日敦高勒嘎查,南至后苏金高勒,长约10.5km;东起伊和德日苏嘎查,西至赛乌苏嘎查,约20km。镶黄旗地势南高北低,地形以低山丘陵为主,由东南向西北递减,平均海拔1 322m。属中温带半干旱大陆性季风气候,地区年降水量267.9mm,主要降水集中在夏季(6-8月),地区年日照时数3 031.6h,平均日照百分率68%,辐射能344.1kJ/m2。地区年平均气温3.1℃,最热月为7月份,平均气温20.4 ℃,最冷月为1月份,平均气温-16.5 ℃(引自内蒙古镶黄旗新宝拉格镇城市总体规划2010-2030年)。
调查地为典型草原,间有小面积荒地,以及人工林草地。根据地质地貌特性,调查地又进一步分成5种不同类型:(1)低丛生禾草地,主要以禾本科植被为主。草群盖度为20%~40%,草层高度4~15cm。(2)禁牧草地Ⅰ:植被主要以禾本科植物羊草为主。草群盖度50%以上。(3)禁牧草地Ⅱ:植被种类繁杂,多样性高。草群盖度35%~55%,草层高度4~35cm。(4)退化草地:优势种植被为冷蒿,草群盖度30%~40%。(5)特殊草地:植被单一,禾本科植物野燕麦为唯一绝对优势种,草群盖度80%以上。
1.2 蝗虫调查方法
蝗虫调查于每年6-8月间进行,每7d调查1次,采用Evans描述的方法[16],以昆虫采集网“Z”字形快速扫网采样法取样,每个样点100 个往返网次(捕网直径30cm),每个样点重复5次,同时用1m2样方框随机选取5个样方,并调查植物群落特征,共15个样地。每个往返网次所捕蝗虫装入保鲜袋内,室内分类鉴定[17-18],统计龄期、种类及数量。植被、蝗虫生物量测定采用称重法。烘干温度≤65 ℃。每个时期蝗虫平均生物量为这个调查时期内调查的每种蝗虫生物量的平均值。
1.3 群落结构测定方法
1.3.1 蝗虫群落多样性
用两种常见的生物多样性指数分析植物和蝗虫多样性。
辛普森多样性指数(Simpson’s diversity index)[21]
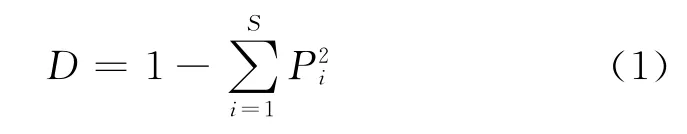
香农-威纳指数(Shannon-Weiner index)[22]
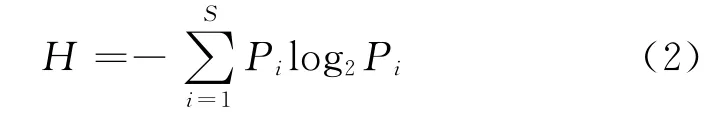
上述公式中,设种i的个体数占群落中总个体数的比例为Pi,S为物种数目。
1.3.2 蝗虫的生态位计算
Hutchinson提出目前广泛使用的基础生态位(fundamental niche)和实际生态位(realized niche)等概念[23-24],以BIOCLIM[25]、栖息地分析(HABITAT)[26]和主域分析(DOMAIN)[27]为代表的环境包络(environmental envelope)理论是最早的生态位量化理论。对一维生态位常用生态位宽度(niche breadth)和生态位重叠(niche overlap)两项定量指标,比较群落中各物种占据空间的大小或利用资源的多少。
多维生态位的构成,时间维指每年蝗虫发生调查6-8 月期间,以7d为单位所构成的“资源”序列,共分为5个“资源”;空间维指不同类型草地;时间-空间维指6—8月以7d为单位乘以不同草地所构成的两维“资源”序列。
生态位宽度公式用Levins(1968)度量公式[28]:
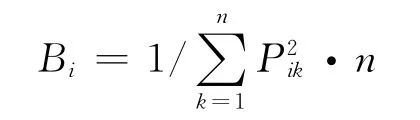
其中Bi为生态位宽度指数;n为资源单元数,Pik为第i物种所占第k资源单元数在所有可利用资源中的比例。生态位重叠公式主要有Hurlbert(1978)度量公式[29]
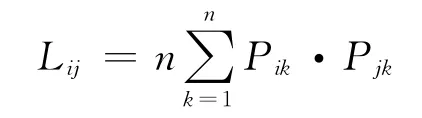
其中,Lij是第i物种重叠第j物种的生态位重叠指数;n为资源单元数,Pik、Pjk为第i、j物种所占第k资源单元数在所有可利用资源中的比例。Lij=0,不共享;Lij=1,平均利用;Lij>1,共同偏嗜利用某些资源。
当物种利用的各个资源类型独立时,多维生态位特征值可用各维生态位特征值的乘积表示[30]。时间-空间二维生态位是时间生态位和空间生态位的累积,反映物种的实际生态位。
1.4 数据分析方法
以上数据处理分析均采用Microsoft Office Excel 2007和SAS 9.2(Statistics Analysis System 9.2,SAS Institute Inc.)数据处理系统进行,显著性水平为P<0.05。
2 结果与分析
2.1 镶黄旗典型草原蝗虫群落组成
蝗虫的群落结构组成受栖息生境、植物群落的组成、气温、降水等环境因子共同作用,对草原生态系统的可持续发展有着重要的影响,蝗虫群落结构主要包括群落的组成、时间结构及空间结构等。我们经过调查将标本进行整理分析,得到主要的蝗虫4科9属11种,其种类及学名缩写如下:亚洲小车蝗(OedaleusdecorusasiaticusB.-Bienko)(OAB)、毛足棒 角 蝗[Dasyhippusbarbipes(Fischer von Waldheim)](DBF)、白边痂蝗[Bryodemaluctuosumluctuosum(Stoll)](BLS)、轮纹异痂蝗[Bryodemellatuberculatumdilutum(Stoll)](BTD)、黄胫异痂蝗[Bryodemellaholdereriholdereri(Krauss)](BHH)、鼓翅皱膝蝗(AngaracrisbarabensisPallas)(ABP)、红翅皱膝蝗[Angaracrisrhodopa(Fischer-Waldheim)](ARF)、华北雏蝗(ChorthippusbrunneushuabeiensisXiaetJin)(CBH)、短 星 翅 蝗(CalliptamusabbreviatusIkonnikov)(CAI)、宽须蚁蝗[Myrmeleotettixpalpalis(Zubovski)](MPZ)、邱氏异爪蝗(EuchorthippuscheuiHsia)(ECH)。各种群总生物量分布如图1。
在所调查的样地中,不同样地蝗虫种类和数量不同。由图1可知,典型草原蝗虫群落物种组成丰富,整个调查区域华北雏蝗和邱氏异爪蝗、短星翅蝗、亚洲小车蝗、宽须蚁蝗种群数量占整个群落比重较大,数量占总蝗虫数的90%以上,其种群在整个蝗虫群落中占有重要地位,华北雏蝗和邱氏异爪蝗是本地区蝗虫种群数量最大的优势种,华北雏蝗和邱氏异爪蝗对栖境选择具有特异性,喜欢栖居在植被盖度高、密度大的栖境,两者的分布与种群地位对镶黄旗植物群落和生境具有指示作用。
2.2 蝗虫种群地位与空间地位划分
内蒙古典型草原11种蝗虫在不同调查时期平均生物量随时间变化(表1)。从表1 中可以看出,不同蝗虫生物量时间变化明显不同。11种蝗虫中,变异系数较小的亚洲小车蝗是作为优势种存在的,宽须蚁蝗、亚洲小车蝗、短星翅蝗分别构成镶黄旗蝗虫早期、中期、晚期优势种。
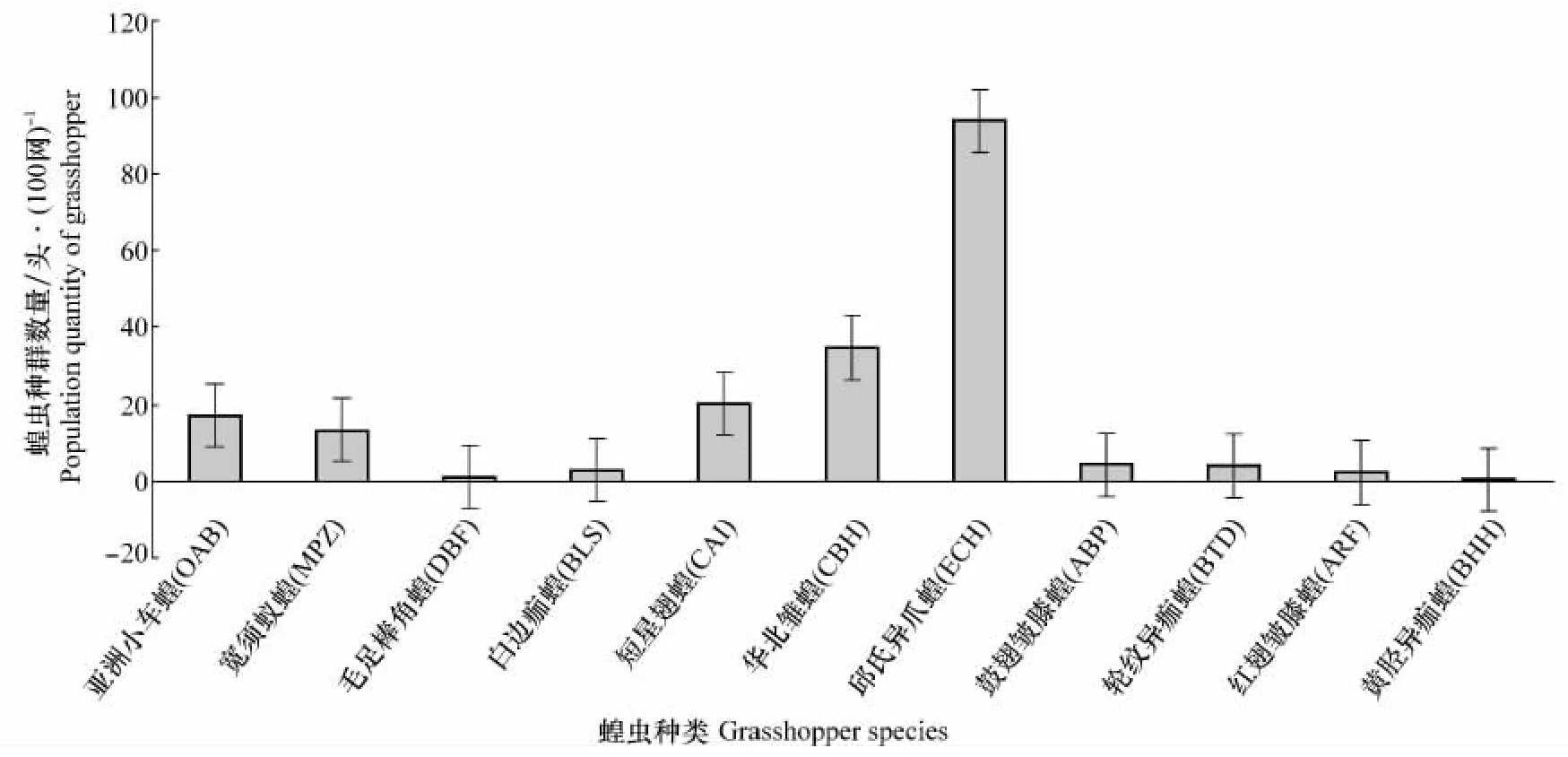
图1 镶黄旗草原蝗虫群落组成Fig.1 Grasshopper community composition in grasslands of Xianghuang Banner
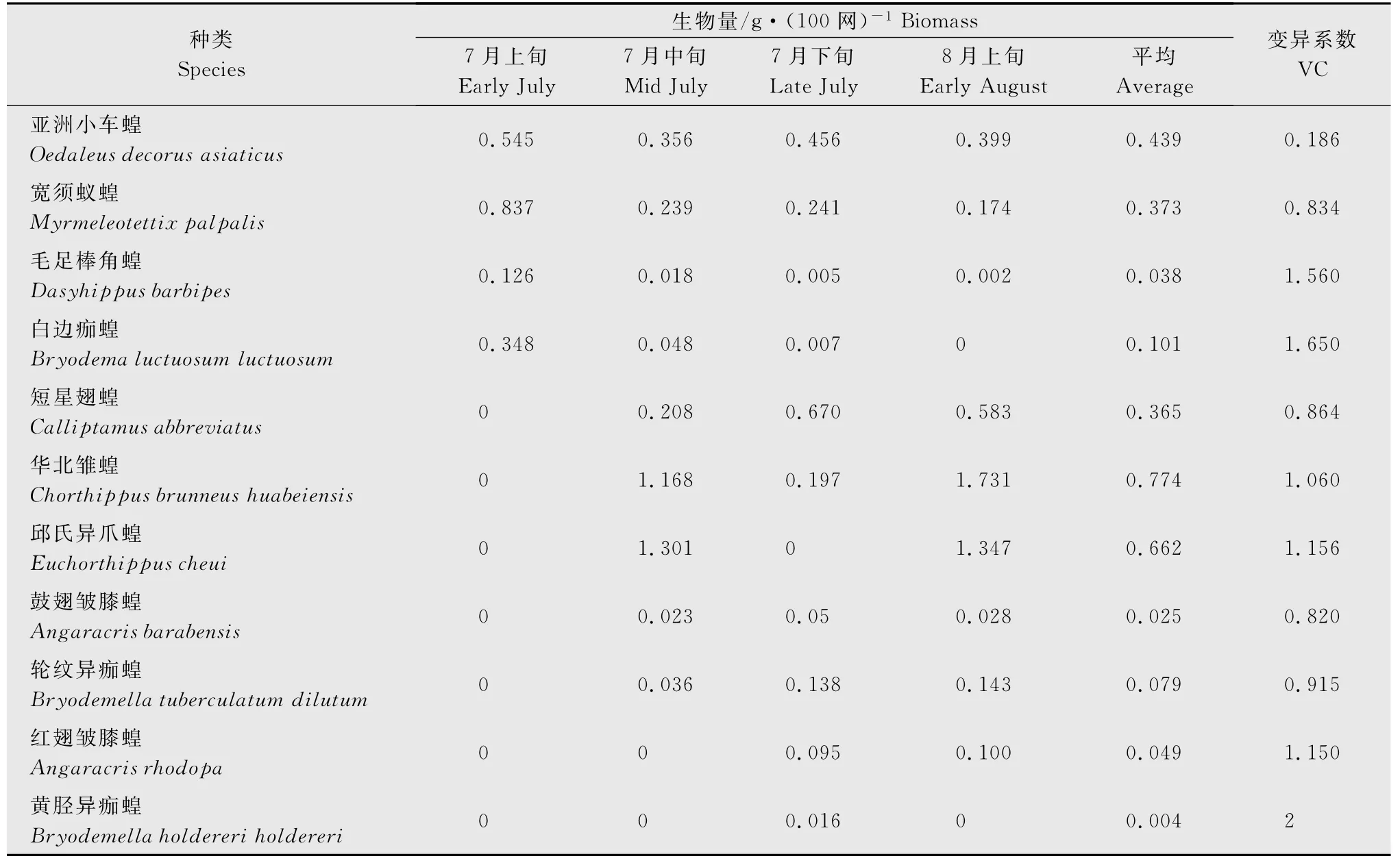
表1 典型草原蝗虫种群生物量季节变化Table 1 Seasonal change of the typical grassland locust population biomass
蝗虫种群动态符合发生期规律,5月中旬早期发生种蝗蝻开始出土,6月至7月上旬,早期发生种如宽须蚁蝗等蝗蝻数量上升,种群数量和生物量开始时较低,随蝗卵的孵化和虫体的增长,蝗虫的数量和生物量随之上升,到一定时间到达高峰,由于食物、栖境等因素的竞争,使蝗蝻存活率有所下降,但随着时间的推进蝗虫个体的生物量和种群的平均生物量上升至后期稳定。
2.2.1 不同种蝗虫种群地位的分析
蝗虫种群地位主要取决于蝗虫种群数量和取食量的大小,研究表明蝗虫取食量的大小往往与身体体重成正相关,同时,考虑到应用种群数量作为种群定量指标所产生的误差,我们以调查所得各蝗虫种群数量经转换为种群生物量将11种蝗虫采用主成分分析法进行了分析比较,结果见图2。

图2 典型草原11种蝗虫种群生物量主成分分析排序图Fig.2 Principal component analysis of eleven grasshopper species in typical grassland
图中G(1)=50.64%;G(2)=81.01%,将11种蝗虫划分为3大类:
(1)优势种,包括亚洲小车蝗(OAB)、宽须蚁蝗(MPZ)、短星翅蝗(CAI)。在典型草原蝗虫群落中种群数量大,其种群的发生对整个群落结构与功能起着重要的影响作用。位于图中第Ⅰ、Ⅱ象限。
(2)附属种,是指优势种以外的发生量相对较少的种类。包括毛足棒角蝗(DBF)、白边痂蝗(BLS)、轮纹异痂蝗(BTD)、黄胫异痂蝗(BHH)、鼓翅皱膝蝗(ABP)、红翅皱膝蝗(ARF)。在图中位于第Ⅲ象限。这些种类由于种群数量较少,或者发生期短暂,而对整个群落的影响作用较小。另一方面,这些蝗虫往往受外界的干扰、气候变化或人类活动的影响,种群数量变化很大,甚至会上升为主要害虫。
(3)稀少种,包括华北雏蝗(CBH)和邱氏异爪蝗(ECH)。在图中位于第Ⅳ象限。这些蝗虫仅在局部地区分布,它们对整个蝗虫群落的作用不够明显,或仅局部偏重发生。
2.2.2 典型草原蝗虫种群空间地位分析
表2是取不同时间过程中,11种典型草原蝗虫在5种不同草地类型中的种群平均生物量。采用主成分分析法对表2 数据进行分析,结果见图3。其中二维坐标图:G(1)=53.44%;G(2)=77.72%。将11种蝗虫划分为4类。

表2 典型草原不同草地类型蝗虫种群生物量的变化1)Table 2 Changes in grasshopper population biomass in different types of typical steppes

图3 典型草原不同草地类型11种蝗虫主成分分析排序图Fig.3 Principal component analysis of eleven grasshopper species in different grassland types
(1)禾草地类,包括亚洲小车蝗(OAB)、宽须蚁蝗(MPZ)。这些种类主要分布于丛生短禾草地和禁牧草场禾草地中。图中位于第Ⅳ象限。
(2)荒草地类,这一类分布多趋于有菊科、百合科等杂草丛生的地方,或湿润的荒滩地。包括白边痂蝗(BLS)、轮纹异痂蝗(BTD)、黄胫异痂蝗(BHH)、鼓翅皱膝蝗(ABP)、红翅皱膝蝗(ARF)、毛足棒角蝗(DBF)。
(3)特殊地类,华北雏蝗(CBH)。仅在局部地区,即调查时的特殊草地类型中分布。
(4)全域类,其食量很杂,发生数量较大,没有明显的分布域差别,主要是短星翅蝗(CAI)、邱氏异爪蝗(ECH)图中位于第Ⅰ、Ⅱ象限。
2.3 草原蝗虫时空生态位分析
以蝗虫发生期6-8月期间所调查的15块样地范围为空间资源,以7d为单位构成时间序列资源,共分为5个“资源”计算典型草原蝗虫生态位,计算结果如表3。
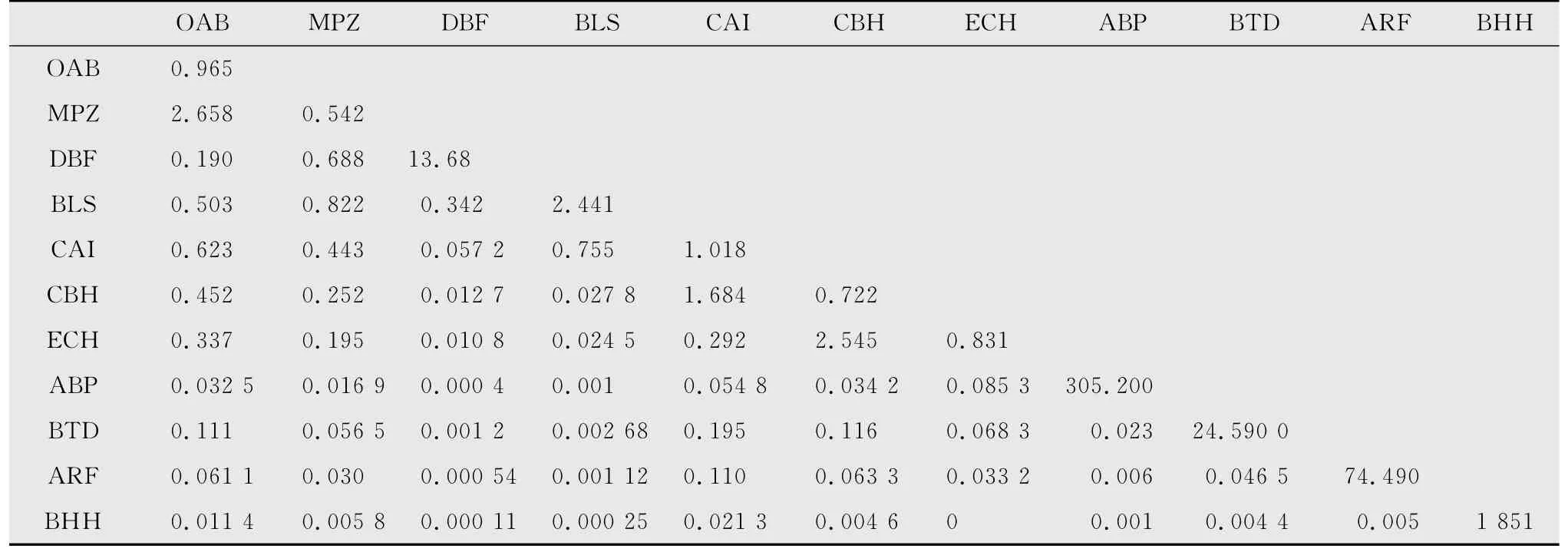
表3 内蒙古镶黄旗草原11种蝗虫时间-空间生态位宽度和生态位重叠1)Table 3 Time-space niche breadth and niche overlap of the eleven grasshopper species in Xianghuang Banner,Inner Mongolia
物种生态位宽度、物种之间的生态位重叠值与物种的竞争与生存对策密切相关,由上表可知,在调查的某些特殊类草地中,时空生态位宽度最大的前4种蝗虫分别是黄胫异痂蝗、鼓翅皱膝蝗、轮纹异痂蝗、红翅皱膝蝗;优势种蝗虫中生态位宽度最大的是短星翅蝗,生态位重叠程度最高的是亚洲小车蝗与宽须蚁蝗,其次是短星翅蝗与亚洲小车蝗,说明它们的可利用资源最为丰富且有很强的利用资源的能力,宽须蚁蝗发生盛期在6 月至8 月,与亚洲小车蝗有较大的时间重叠,两种蝗虫都以取食禾本科牧草为主[31],因此有较大的食物和空间竞争,而短星翅蝗主要嗜食菊科植物,并对亚洲小车蝗喜食的禾本科牧草羊草有所喜食,食性相对较广,发生期从7月至8月末,因此,占有较大的时空生态位并与亚洲小车蝗有很大的生态位重叠;邱氏异爪蝗与华北雏蝗的生态位重叠程度较高,因为两种蝗虫在同一草地区域内大量聚集分布,使两者生态位重叠度较高。
2.4 典型草原植物群落与蝗虫群落聚类分析及潜在危害种的预测
群落相似性是用来测定不同生境、群落之间的相似程度,划分群落及探讨群落组成异同的重要依据。将蝗虫群落与植物群落聚类可以分析蝗虫的优势类群和存在的优势资源,是探讨蝗虫生态位与预测蝗虫种群发生与稳定的重要依据。优势种蝗虫的分布与优势种植被类型有密切的关系,优势种植被的数量变化直接体现了植被的变化,植物的优势度、相对密度,地上部分相对生物量基本上反映了其在群落中的地位。以这3个特征因子作为植物群落聚类分析的运算单位,用蝗虫的相对多度、生物量作为蝗虫群落聚类分析的运算单位,利用欧氏距离将16个样地植被群落和蝗虫群落进行系统聚类分析,得到16块样地植被群落与蝗虫群落的系统聚类结果。通过样地的聚类树状图可直观地看出各样地之间在组成上的相近程度,图4对样地植物群落参数之间的相似性进行聚类,取λ=0.70,植被群落可根据聚类结果分为禾本科杂草优势样地(包括针茅、羊草、隐子草等,OB1、OB2、OB3、OB4、OB5、OB6、OB7、OB8、OB9、OB10、OB11、OB12、OB14、OB15号),阔叶草优势样地(包括冷蒿、扁蓿豆等,OB16号样地)和特殊类样地(OB13号样地)三类,禾草草原样地之间的相似度指数均较高,在不同的样地中大体可以将蝗虫种群分为4类不同的群落,以亚洲小车蝗占优势种的群落样地(OB1、OB2、OB3、OB4、OB6号样地),以宽须蚁蝗占优势种的样地(OB1、OB3、OB14号样地)和以短星翅蝗占优势种的样地(OB2、OB5、OB7、OB16号样地),以及其他类蝗虫样地(雏蝗,OB6号样地)。性质相同或者相近的生境中,蝗虫的物种相似度较高,物种间存在着交流,而不同性质的生境常分布着对环境有特殊生态要求的物种,显示出较低的相似性。由图4 可知,OB1、OB2、OB3、OB4、OB14号样地植被特征非常相近,以禾本科草占优势,与之相对应,图5 显示,OB1、OB2、OB3、OB4号样地的蝗虫群落特征也非常相近,以亚洲小车蝗占优势,所以,可以预测亚洲小车蝗在14号地发生危害的可能性较大,同理,宽须蚁蝗在OB2、OB4号样地大量暴发的可能性也很大,由于短星翅蝗本身生态位宽度较大,与亚洲小车蝗和宽须蚁蝗、短星翅蝗生态位重叠较高,占有的“资源”潜力大,根据聚类结果,可以预测,短星翅蝗在OB3、OB8、OB9号样地大量发生的可能性较大。植被群落分类大体上对应了蝗虫群落分类,但并非一一对应,一方面在小生境中,植被群落特征影响了蝗虫群落的不同分布,另一方面大生境中影响蝗虫群落特征的因素不只是植被特征,小生境中,植被群落特征往往与蝗虫群落特征呈现显著的相关性,植被的种类和特征参数影响了蝗虫群落结构,而大生境中,影响蝗虫群落的因素比较复杂,包括温度、降水等气象因素及土壤、人类活动和小生境气候等因素的共同作用,因此,我们可以根据聚类的草地类型和气候条件,预测蝗虫在某一地区的发生与危害。
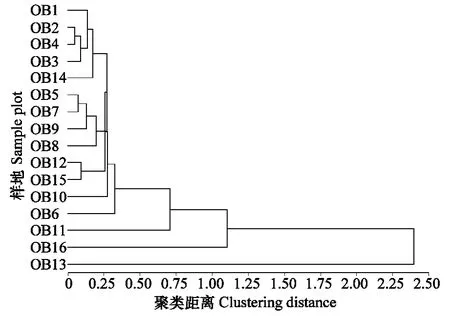
图4 不同样地植物群落聚类图Fig.4 Clustering analysis of phytocoenosium in different sample plots
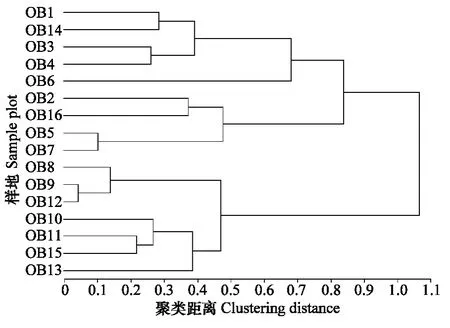
图5 不同样地蝗虫群落聚类图Fig.5 Clustering analysis of grasshopper community in different sample plots
3 讨论
草原生态系统中蝗虫种群与植被多样性的研究是阐明物种多样性对生态系统功能作用的重要途径之一。不同种类的蝗虫个体之间,以及同种蝗虫不同发育阶段和性别的个体之间生物量差异很大。所以单纯使用种群个体数量来描述蝗虫种群的动态,以及种群间的相互关系和群落中的作用和地位是不准确的。以种群生物量作为衡量种群作用与功能的定量指标相对较为客观、实际。不同的蝗虫种群各自选择适宜的时间-空间生态位,是为了避免种群之间的竞争,或者是尽可能充分利用有限的时间-空间作为营养、繁殖或栖息场所。物种生态位可以从竞争物种对时间、空间和食物资源的分配策略来探讨竞争共存机制、协同进化、多物种群落的演替及多样性等问题[32-35]。镶黄旗11种蝗虫种群生物量随时间和植物多样性呈现规律一致性变化,该地区宽须蚁蝗、亚洲小车蝗、短星翅蝗分别作为早、中、晚优势种存在,分析样地植物群落特征与蝗虫群落特征关系则发现,植被群落影响蝗虫群落的结构和分布,在较大生态群落范围内,蝗虫群落随着生境的破碎化与人类活动的影响而变化,蝗虫群落与植被群落的相互作用与影响正在发生着变化,本文把蝗虫种群依据不同空间地位和草地类型划分,因为蝗虫的分布与样地的植被特征有着较高的相关性,蝗虫种团资源利用的分化来自于蝗虫适应性的分化和蝗虫—植物协同进化的关系[36],这说明优势种植被对优势种蝗虫的分布具有显著影响。如此可以根据蝗虫生物量与样地植被特征预测并划分蝗虫宜生区。生态位的保守性(conservatism)与等价性(equivalency)[37-42],是一直争论的问题,Peterson认为基础生态位在小时间尺度上保持稳定,在大时间尺度上缓慢变化[37,39],但是,Broennimann的研究则认为,物种的基础生态位会随着其生活的群落不同而发生变化[38]。时空间生态位的大小主要取决于生境环境条件的复杂程度,本文通过大面积长时间调查,分析不同优势种蝗虫时间动态、不同区域内的空间分布,植物群落“资源”与蝗虫群落之间的聚类分析,认为,蝗虫在时间和空间“资源”的利用上是相互关联的。蝗虫对时间和空间“资源”的利用随着季节变化而转移,其转移速度因种类而异,受环境因子的明显影响。我们可以利用蝗虫群落与植物群落聚类分析蝗虫的优势类群和存在的优势资源,并可根据群落的相似性和蝗虫生态位预测蝗虫的迁移与危害。优势种蝗虫种群时空生态位变化,也在揭示着草原蝗虫群落与植被群落的演替、人类活动及气候的变化。
群落类型实际上是空间生态位的一种表现方式,主要反映群落中物种的空间配置情况,一般来说,禾草地草场蝗虫种群数量较高;荒漠地带种类数较多,个体数较低。生态位宽度可以反映生物对环境的适应性以及利用环境资源的广泛性,即物种对环境资源利用能力的强弱。蝗虫种群在时空资源利用上的分化十分明显。不同草地类型有着不同的种团组合,同一种团中同等地位的种群在时间上又有着明显的分化,反映了蝗虫群落对环境条件的适应特性及充分合理地利用资源,以达到种群发生与资源利用相平衡的生态对策。
本文对草原蝗虫群落的研究,将为估测蝗虫潜在危害和监测预警及草原蝗害的生态治理提供理论依据,并对了解草原生态系统的结构功能以及正确评价人类活动和全球气候变化对物种的影响具有重要的理论意义。
[1]康乐,陈永林.草原蝗虫时空异质性的研究[M]∥中国科学院内蒙古草原生态系统定位站.草原生态系统研究(第四集).北京:科学出版社,1992:109-123.
[2]康乐,陈永林.草原蝗虫营养生态位的研究[J].昆虫学报,1994,37(2):178-189.
[3]贺达汉.荒漠草原蝗虫时空生态位的研究[J].宁夏农学院学报,1997,18(2):2-8.
[4]Grinnell J.Barriers to distribution as regards birds and mammals[J].American Naturalist,1914,48:248-254.
[5]Grinnell J.The niche-relationships of the California Thrasher[J].The Auk,1917,34:427-433.
[6]Grinnell J.Geography and evolution [J].Ecology,1924,5:225-229.
[7]Pulliam H R.On the relationship between niche and distribution[J].Ecology Letters,2000,3:349-361.
[8]Sobek-Swant S,Kluza D A,Cuddington K,et al.Potential distribution of emerald ash borer:what can we learn from ecological nichemodels using Maxent and GARP-Forest[J].Rangeland Ecology and Management,2012,281:23-31.
[9]Monahan W B,Tingley M W.Niche tracking and rapid establishment of distributional equilibrium in the house sparrow show potential responsiveness of species to climate change[J].PLoS ONE,2012,7:e42097.
[10]Taylor S,Kumar L,Reid N.Impacts of climate change and land-use on the potential distribution of an invasive weed:a case study ofLantanacamarain Australia [J].Weed Research,2012,52:391-401.
[11]Ebeling S K,Welk E,Auge H,et al.Predicting the spread of an invasive plant:combining experiments and ecological niche model[J].Ecography,2008,31:709-719.
[12]Václavík T,Meentemeyer R K.Invasive species distribution modeling(iSDM):are absence data and dispersal constraints needed to predict actual distributions[J].Ecological Modelling,2009,220:3248-3258.
[13]Gallien L,Douzet R,Pratte S,et al.Invasive species distribution models—how violating the equilibrium assumption can create new insights[J].Global Ecology and Biogeography,2012,21:1126-1136.
[14]Pearson R G,Dawson T P.Predicting the impacts of climate change on the distribution of species:are bioclimate envelope models useful[J].Global Ecology and Biogeography,2003,12:361-371.
[15]Swab R M,Regan H M,Keith D A,et al.Niche models tell half the story:spatial context and life-history traits influence species responses to global change[J].Journal of Biogeography,2012,39(7):1266-1277.
[16]Evans E.Grasshopper(Insecta:Orthoptera:Acrididae)assemblages of tallgrass prairie:influences of fire frequency,topography and vegetation[J].Canadian Journal of Zoology-Revue Canadienne De Zoologie,1988,66:1495-1501.
[17]吴虎山,能乃扎布.呼伦贝尔市草地蝗虫[M].北京:中国农业出版社,2009:1-175.
[18]白文辉,徐绍庭.内蒙古草原蝗虫名录[J].中国草地,1985(1):41-47.
[19]Taylor L R.Aggregation,variance and the mean[J].Nature,1961,189:732-735.
[20]庞雄飞,尤民生.昆虫群落生态学[M].北京:中国农业出版社,1996:1-147.
[21]Simpson E H.Measurement of diversity[J].Nature,1949,163:688.
[22]Chao A,Shen T J.Nonparametric estimation of Shannon’s index of diversity when there are unseen species in sample[J].Environmental and Ecological Statistics,2003,10(4):429-443.
[23]Hutchinson G E.An introduction to population ecology[M].New Haven:Yale University Press,1978.
[24]Colwell R K,Rangel T F.Hutchinson’s duality:the once and future niche[J].Proceedings of the National Academy of Sciences of the United States of America,2009,106:19651-19658.
[25]Busby J.BIOCLIM—a bioclimate analysis and prediction system[J].Plant Protection Quarterly,1991,6:8-9.
[26]Walker P A,Cocks K D.HABITAT:aprocedure for modelling a disjoint environmental envelope for a plant or animal species[J].Global Ecology and Biogeography Letters,1991,1:108-118.
[27]Carpenter G,Gillison A N,Winter J.DOMAIN:a flexible modelling procedure for mapping potential distributions of plants and animals[J].Biodiversity and Conservation,1993,2:667-680.
[28]Levins,R.Evolution in changing environments:some theoretical explorations[M].Princeton,N J:Princeton University Press,1968.
[29]Hurlbert Stuart H.The measurement of niche overlap and some relatives[J].Ecology,1978,59:67-77.
[30]Cody M L.Competition and the structure of bird communities[M].New Jersey:Princeton University Press,1974:1-326.
[31]卢辉,余鸣,张礼生,等.不同龄期及密度亚洲小车蝗取食对牧草产量的影响[J].植物保护,2005,31(4):55-58.
[32]汪信庚,刘树生,吴晓晶,等.杭州郊区菜蚜种群的空间动态[J].应用生态学报,1997,8(6):599-604.
[33]李志胜,黄咏俏,尤民生.芥蓝田主要害虫种群生态位研究[J].昆虫知识,2005,42(4):409-412.
[34]王智,曾伯平,王文彬.黄鳝体内寄生虫生态位研究[J].湖南农业大学学报(自然科学版),2008,3(2):204-206.
[35]贺达汉.荒漠草原蝗虫群落特征研究[M].银川:宁夏人民出版社,1997:1-135.
[36]Kang L,Chen Y L.Multidimension analysis of resource utilization in assemblages of rangeland grasshoppers(Orthoptera:Acrididae)[J].Insect Science,1994(3):264-282.
[37]Peterson A T,Soberón J,Sánchez-Cordero V.Conservatism of ecological niches in evolutionary time[J].Science,1999,285:1265-1267.
[38]Broennimann O,Treier U A,Müller-Schrer H,et al.Evidence of climatic niche shift during biological invasion[J].Ecology Letters,2007,10:701-709.
[39]Peterson A T.Ecological niche conservatism:a time-structured review of evidence[J].Journal of Biogeography,2011,38:817-827.
[40]Warren D L,Glor R E,Turelli M.Environmental niche equivalency versus conservatism:quantitative approaches to niche evolution[J].Evolution,2008,62:2868-2883.
[41]Wiens J J,Graham C H.Niche conservatism:integrating evolution,ecology,and conservation biology[J].Annual Review of Ecology Evolution and Systematics,2005,36:519-539.
[42]Graham C H,Ron S R,Santos J C,et al.Integrating phylogenetics and environmental niche models to explore speciation mechanisms in dendrobatid frogs[J].Evolution,2004,58:1781-1793.
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
小哥白尼(野生动物)(2022年5期)2022-08-15
贵州畜牧兽医(2022年3期)2022-06-28
现代园艺(2021年23期)2021-12-01
新农业(2020年18期)2021-01-07
花卉(2020年24期)2021-01-04
今日农业(2020年19期)2020-12-14
科学大众(2020年10期)2020-07-24
安徽农学通报(2020年7期)2020-05-26
故事作文·低年级(2019年6期)2019-06-20
