
鸡KLF3基因的表达规律及其对脂肪细胞分化的影响研究
2015-03-22 08:13王海霞张志威王宇祥曹志平
畜牧兽医学报 2015年1期
王海霞,张志威,2,贺 綦,王 宁,王宇祥,曹志平,李 辉*
(1.东北农业大学动物科学技术学院,农业部鸡遗传育种重点实验室,黑龙江省普通高等学校动物遗传育种与繁殖重点实验室,哈尔滨 150030; 2.石河子大学医学院,石河子 832000)
鸡KLF3基因的表达规律及其对脂肪细胞分化的影响研究
王海霞1,张志威1,2,贺 綦1,王 宁1,王宇祥1,曹志平1,李 辉1*
(1.东北农业大学动物科学技术学院,农业部鸡遗传育种重点实验室,黑龙江省普通高等学校动物遗传育种与繁殖重点实验室,哈尔滨 150030; 2.石河子大学医学院,石河子 832000)
为研究鸡KLF3(GallusgallusKrüppel-like factor 3,gKLF3)基因的表达规律及其对脂肪细胞分化的影响,利用qRT-PCR检测了其在肉鸡脂肪组织和脂肪细胞中的表达特点,并利用基因过表达技术研究gKLF3基因对脂肪细胞分化的影响。结果显示,gKLF3在2~10周龄肉鸡腹部脂肪组织中持续表达,10周龄高脂肉鸡腹部脂肪组织gKLF3表达量显著高于低脂肉鸡(P<0.05);gKLF3在鸡成熟脂肪细胞的表达量低于前脂肪细胞(P<0.01);体外培养的鸡前脂肪细胞经油酸诱导分化后,gKLF3基因表达量均有低于对照组的趋势;过表达gKLF3基因有抑制脂肪细胞分化,以及抑制C/EBPα、FAS基因表达(P<0.01)的趋势。研究结果表明,gKLF3基因在鸡腹部脂肪组织生长发育过程中发挥重要作用,其可能是通过抑制C/EBPα、FAS基因表达进而抑制脂肪细胞分化实现的。
鸡;KLF3基因;脂肪细胞分化;基因表达
Krüppel-like factor 3(KLF3)具有3个典型的C2H2锌指结构,是Sp1/KLFs蛋白家族的一个成员,早在1996年,从小鼠红细胞(Murine erythroid cells)中克隆得到[1]。因为在其N端具有较多的碱性氨基酸,所以KLF3又被称为碱性KLF因子(Basic Krüppel like factor,bKLF)[1]。目前,KLF3已经被证实在哺乳动物的红细胞生成[2]、免疫反应[3]、自噬作用[4]、心肌细胞分化[5]和脂肪细胞分化[6]中发挥重要作用。
KLF3基因敲除小鼠(KLF3-/-)比正常小鼠拥有较少的白色脂肪组织,并且附睾脂垫中的脂肪细胞无论是数量,还是体积都小于正常小鼠[6-7]。细胞水平的研究证实KLF3在脂肪细胞分化中发挥作用。3T3-L1前脂肪细胞系的研究表明,KLF3基因的表达量随着脂肪细胞的分化而下降,强制表达KLF3基因抑制3T3-L1脂肪细胞的分化[6],进一步的机制研究显示,KLF3通过抑制C/EBPα基因的表达抑制脂肪细胞分化。在秀丽隐杆线虫中的研究表明,KLF3通过调控脂肪酸去饱和途径基因的表达水平,参与调控维持正常脂肪酸复合物的形成[8]; 此外,KLF3是调控脂蛋白组装、分泌和脂肪酸β-氧化的重要基因[9]。
目前还没有关于KLF3基因在鸟类中的研究报道。本研究以东北农业大学肉鸡高、低脂系为试验材料,利用qRT-PCR的方法检测gKLF3基因在2~10周龄高低脂系肉鸡腹部脂肪组织、12日龄Arbour Acres(AA)仔鸡成熟脂肪细胞和前脂肪细胞,以及脂肪细胞分化过程中的表达特点;同时,通过基因过表达试验研究了gKLF3基因对鸡前脂肪细胞分化的影响,为进一步研究gKLF3基因调控脂肪分化的机理奠定基础。
1 材料与方法
1.1 材料
东北农业大学肉鸡高低脂选择系第11世代2~10周龄公鸡[10]。每个时间点,高脂系、低脂系各取3只为试验材料,采集新鲜的腹脂样品于-80 ℃保存备用。12日龄的AA肉仔鸡由本课题组饲养。鸡KLF3基因过表达质粒(pCMV-myc-gKLF3)由本实验室保存。
1.2 方法
1.2.1 鸡前脂肪细胞和成熟脂肪细胞分离 取12日龄AA肉仔鸡公鸡,超净台中取腹部脂肪组织(腹部及肌胃周围脂肪组织)放入装有磷酸盐缓冲液(PBS)的平皿中;用PBS将脂肪组织洗2遍,尽量除去血管和筋膜,用眼科剪剪碎组织,大约剪到1 mm3体积大小;用0.1%Ⅰ型胶原酶的消化液消化组织,37 ℃消化65 min(每5 min摇匀1次);消化完毕,加入全培养基终止消化,吸管吹打,静置分层,取上层油状物下的絮状物即为成熟脂肪细胞,其余部分分别经100和600目不锈钢筛网过滤,滤液分装入离心管中,2 000 r·min-1离心10 min;弃去上清,加入红细胞裂解液重悬细胞,室温静置10 min后,2 000 r·min-1离心10 min,弃上清,获得的沉淀即前脂肪细胞[11]。
1.2.2 鸡前脂肪细胞的培养和诱导分化 将分离的鸡前脂肪细胞沉淀加入适量的全培养基重悬、吹匀,置37 ℃、5% CO2培养箱中培养,每2 d换液1次,一般4~5 d长满,即可传代;当传代培养的鸡前脂肪细胞汇合至80%左右时,加入诱导培养基(含有160 μmol·L-1油酸浓度的培养基)进行诱导分化,每2 d换1次诱导培养基继续培养。
1.2.3 总RNA提取及反转录 按照TRIzol 试剂(Invitrogen)说明书提取鸡脂肪组织、成熟脂肪细胞和前脂肪细胞的总RNA。
1.2.4 油红O提取比色 在鸡前脂肪细胞诱导分化后的24、72、120 h,分别弃去培养基,用PBS洗3次,用10%甲醛固定30 min;然后用PBS洗3次,蒸馏水洗3次,再用0.5%的油红O染色液染色40 min;弃去多余的染色液,用PBS冲洗3次,室温干燥;加入100%异丙醇溶解细胞中的油红O,15 min后用分光光度计(510 nm)记录光吸收值。
1.2.5半定量PCR 根据NCBI数据库gKLF3基因预测序列XM_427367,利用Primer premier 5.0结合UCSC鸡基因组数据库跨内含子设计gKLF3基因表达分析引物,以gGAPDH基因为内参,半定量PCR用到的引物详见表1。扩增片段大小gKLF3基因为276 bp,gGAPDH基因为185 bp。循环次数分别为28和25。以脂肪组织和脂肪细胞的cDNA为模板,扩增片段,琼脂糖凝胶电泳获得电泳图片,利用Quantity one Bio-rad软件检测基因电泳条带的积分光密度IOD(Integrated option density,IOD)值。基因的相对表达量:

表1 用于PCR的引物
Table1PrimersusedforPCR

基因Gene引物(5'-3')PrimergKLF3F:CCAGCCAGTTCCTTTCATR:ACTTCCTGCGGAGACAATC/EBPαF:AGCTCGACCCGCTGTACR:TGTCTTTTTGGATTTGCFASF:AAGGAGGAAGTCAACGGR:TTGATGGTGAGGAGTCGβ-actinF:TCTTGGGTATGGAGTCCTGR:TAGAAGCATTTGCGGTGGgGAPDHF:CTGTCAAGGCTGAGAACGR:GATAACACGCTTAGCACCA
1.2.7 Western blot 待细胞培养至预定时间,弃培养基,室温下用PBS清洗细胞。然后向6孔板中加入细胞裂解液(RIPA Buffer)(每孔0.15 mL),放置于冰上,轻轻摇动,作用15 min。裂解完成后,将含有细胞的裂解液移至1.5 mL离心管中。10 000 r·min-14 ℃离心10 min后,将上清(细胞总裂解物)转至新的离心管,与等体积凝胶加样缓冲液(2×)混合,在100 ℃加热3~5 min使蛋白质变性。每个样品取10~20 μL,采用BIO-RAD 的Mini-PROTEAN3电泳系统进行常规SDS-PAGE电泳。电泳结束后,采用BIO-RAD的Mini Trans-Blot将样品转移至PVDF膜,将膜置于封闭液(5%脱脂乳的PBST),室温反应1 h,或4 ℃过夜。洗去膜上的封闭液,将膜孵育在含一抗(Anti-HA Tag Mouse Monoclonal Antibody)的PBST溶液,置于摇床上,室温反应1 h;洗膜后将膜孵育在含二抗(Goat Anti-Mouse IgG,HRP Conjugated)的PBST溶液;再次洗膜后常规ECL显色。
1.2.8 细胞转染 细胞转染按照罗氏FuGENE HD转染试剂操作说明书进行。
1.2.9 数据分析 数据分析采用Student-t,数据表示为“平均数±标准差(Mean±SD)”,以P<0.05表示差异显著,P<0.01表示差异极显著。
2 结 果
2.1gKLF3基因在高低脂系11世代2~10周龄肉鸡脂肪组织的表达分析
利用半定量PCR分析了gKLF3基因在东北农业大学肉鸡高、低脂系11世代2~10周龄肉鸡脂肪组织中的表达。结果显示,gKLF3在2~10周龄肉鸡腹部脂肪组织的生长发育过程中持续表达,10周龄时,高脂肉鸡腹部脂肪组织gKLF3表达水平显著高于低脂肉鸡(P<0.05,图1)。
2.2 鸡脂肪细胞中gKLF3基因的表达规律分析
利用qRT-PCR方法分析AA肉鸡前脂肪细胞和成熟脂肪细胞中gKLF3基因的表达水平,结果显示,gKLF3基因在鸡前脂肪细胞(Stromal-vascular cell,SV)中的表达量高于成熟脂肪细胞(Fat cell,FC)(P<0.01, 图 2A)。此外,本研究发现,体外培养的鸡前脂肪细胞经油酸诱导分化后,gKLF3基因的表达量在4个时间点(24、48、72和96 h)均有低于对照组(未经油酸诱导)的趋势,但没有达到显著水平(图 2B)。
2.3 过表达gKLF3对鸡前脂肪细胞分化的影响
为了进一步研究gKLF3基因在鸡前脂肪细胞分化过程中的作用,本研究利用过表达技术分析了gKLF3基因对鸡前脂肪细胞分化的影响。在鸡前脂肪细胞中转染质粒pCMV-myc-gKLF3和pCMV-myc(Empty Vector,EV),48 h后回收细胞,用myc标签抗体进行Western blot,结果显示pCMV-myc-gKLF3质粒能在前脂肪细胞中成功表达蛋白(图3A)。将鸡KLF3基因过表达质粒(pCMV-myc-gKLF3)转染到原代培养的鸡前脂肪细胞中,培养24 h后,加入油酸诱导细胞分化,油红O提取比色结果显示,在诱导后的24、72和120 h,与转染空载体的对照组细胞相比,过表达gKLF3基因的鸡前脂肪细胞中脂滴沉积出现减少的趋势,但没有达到显著水平(图3B)。

柱状图表示gKLF3的相对表达量(平均数±标准差),*表示差异显著(P<0.05)The diagrams show the relative expression of gKLF3 gene(Mean±SD).* means significant difference(P<0.05)图1 gKLF3基因在脂肪组织发育过程中的表达Fig.1 Tissue expression pattern of gKLF3 gene in abdominal fat tissue during development

A.KLF3基因在原代分离(未经培养)的鸡前脂肪细胞和成熟脂肪细胞中的表达规律;B.KLF3基因在油酸诱导分化的前脂肪细胞和对照组(未经油酸诱导分化)中的表达规律。**.差异极显著(P<0.01)。下同A.KLF3 mRNA expression in chicken preadipocytes(SV) and mature adipocytes(FC) were detected;B.KLF3 mRNA expression in preadipocytes which induced into differentiation by oleate and control group(which not induced into differentiation) was detected .** indicates significant difference(P<0.01).The same as below图2 gKLF3基因在鸡前脂肪细胞和成熟脂肪细胞中表达特性Fig.2 Expression characteristics of gKLF3 gene in chicken preadipocytes and adipocytes
2.4 过表达gKLF3基因对鸡前脂肪细胞分化标志基因的影响
为了分析gKLF3基因对鸡前脂肪细胞分化的影响,利用qRTPCR检测了脂肪细胞分化过程的标志基因(FAS、C/EBPα)的表达变化。结果发现,过表达gKLF3基因,在24 h显著抑制C/EBPα基因的表达(P<0.01),在72 h显著抑制FAS、C/EBPα基因的表达(P<0.01,图4)。
3 讨 论
KLF家族的成员在哺乳动物脂肪发育中发挥着重要作用[12-14]。在3T3-L1细胞系中的研究显示,KLF3是脂肪细胞分化的负调控因子,但是KLF3基因敲除小鼠的脂肪组织减少[6],表明了KLF3在脂肪组织生长发育过程中具有多个调控功能,目前还没有关于鸟类KLF3的研究报道,本研究结果显示,gKLF3基因在高低脂系2~10周龄肉鸡腹部脂肪组织中均有表达(图1),暗示了与小鼠KLF3相似,gKLF3基因在鸡脂肪组织生长发育过程中具有调控作用;在10周龄时,高脂肉鸡gKLF3基因表达水平显著高于低脂肉鸡(P<0.05,图1),暗示gKLF3的表达水平可能是影响肉鸡腹部脂肪组织沉积的一个因素。
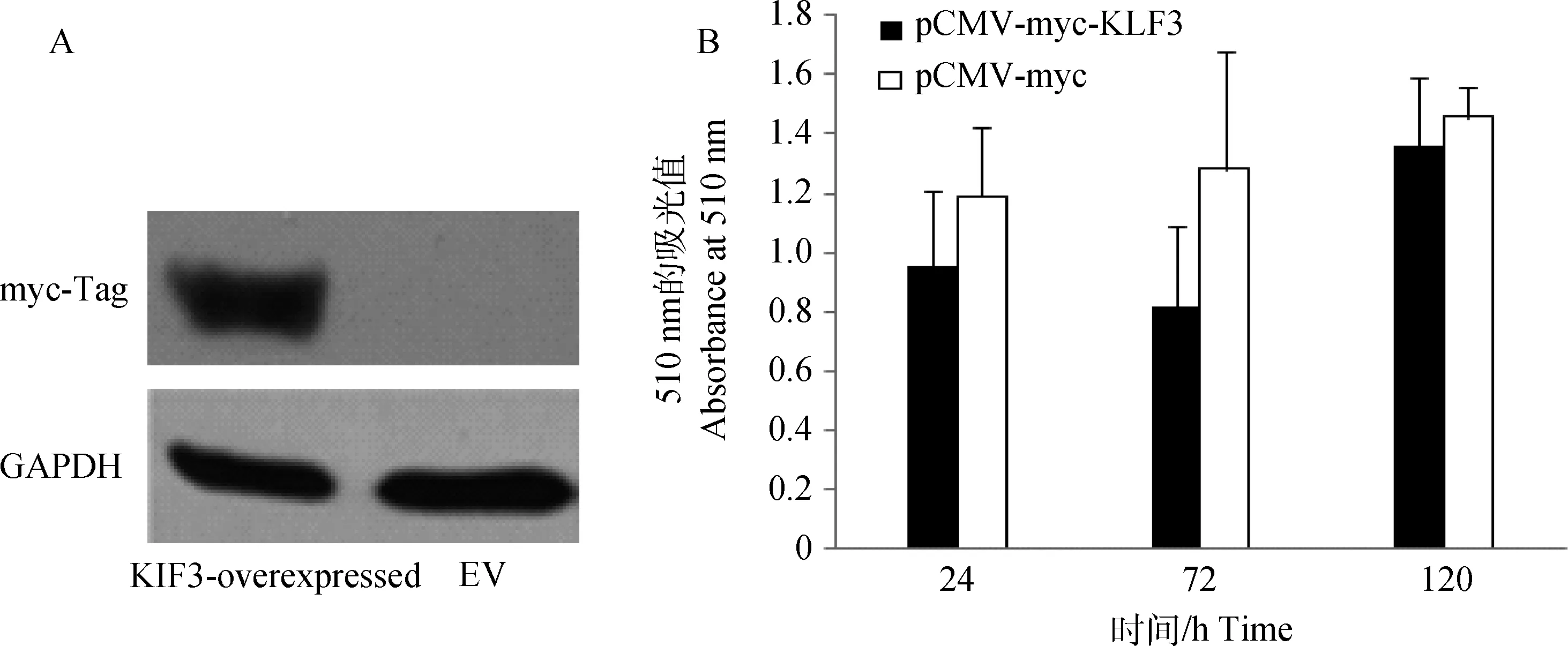
A.Western blot检验pCMV-myc-gKLF3在前脂肪细胞中的过表达效果;B.油红O提取比色分析过表达gKLF3基因对脂肪细胞脂滴沉积的影响A.Western blot analysis of gKLF3 overexpression in chicken preadipocytes transfected with pCMV-myc-gKLF3 or EV;B.The lipid content of chicken preadipocytes which were transfected with pCMV-myc-gKLF3 and pCMV-myc,induced into differentiate by oleate,and absorbance was measured at 510 nm图3 过表达gKLF3基因对鸡前脂肪细胞分化的影响Fig.3 Effects of gKLF3 gene over-expression on chicken preadipocyte differentiation

图4 过表达gKLF3基因对鸡前脂肪细胞分化标志基因的影响Fig.4 Expression analysis of differentiation marker genes in gKLF3-overexpressing chicken preadipocytes induced into differentiation by oleate
进一步分析gKLF3基因在前脂肪细胞和成熟脂肪细胞中的表达发现,gKLF3基因在前脂肪细胞中的表达量高于成熟脂肪细胞(P<0.01,图2A),并且在油酸诱导分化的鸡脂肪细胞中,gKLF3表达水平有普遍低于对照组的趋势,暗示与哺乳动物相似[6],gKLF3基因在脂肪细胞分化中可能具有负调控作用。为了进一步研究gKLF3基因对鸡脂肪细胞分化的作用,笔者在鸡前脂肪细胞中过表达gKLF3基因,油红O提取比色显示,在油酸诱导后的24、72和120 h,过表达gKLF3基因的脂肪细胞呈现脂滴形成减少的趋势(图3B),进一步暗示gKLF3基因与小鼠KLF3类似[6],在体外具有抑制前脂肪细胞分化的作用。
C/EBPα是哺乳动物和鸟类脂肪形成的重要调控因子[12,14-16]。C/EBPα基因敲除小鼠的研究表明,C/EBPα表达是白色脂肪组织发育的必须条件[17]。此外,C/EBPα在脂肪细胞终末分化中发挥重要调控作用[18-20]。FAS基因在动物体脂沉积中发挥了重要的作用[21],它可以将碳水化合物合成脂肪酸,并以甘油三酯的形式储存[22]。本研究发现,过表达gKLF3基因,在72 h会显著抑制FAS、C/EBPα基因的表达,在24 h显著抑制C/EBPα基因的表达(图4,P<0.01)。笔者推断,与哺乳动物相似[6],gKLF3基因可能通过抑制C/EBPα基因的表达抑制脂肪细胞分化。而gKLF3对FAS具有转录抑制作用,则暗示了与线虫KLF3[8]相似,KLF3可能具有抑制脂肪酸从头合成和调控脂类沉积的功能。
综上所述,本研究结果显示gKLF3和已经报道的哺乳动物及线虫中KLF3相似,在肉鸡脂肪组织生长发育中至少具有2个功能,即抑制脂肪细胞分化和调控脂类合成代谢。此外,gKLF3是否直接调控C/EBPα、FAS基因的表达等具体作用机制,还有待荧光素酶活性检测、染色质免疫共沉淀等试验的进一步分析和验证。
[1] CROSSLEY M,WHITELAW E,PERKINS A,et al.Isolation and characterization of the cDNA encoding BKLF/TEF-2,a major CACCC-box-binding protein in erythroid cells and selected other cells[J].MolCellBiol,1996,16(4):1695-1705.
[2] FUNNELL A P,MALONEY C A,THOMPSON L J,et al.Erythroid Krüppel-like factor directly activates the basic Krüppel-like factor gene in erythroid cells[J].MolCellBiol,2007,27(7):2777-2790.
[3] ULGIATI D,SUBRATA L S,ABRAHAM L J.The role of Sp family members,basic Krüppel-like factor,and E box factors in the basal and IFN-gamma regulated expression of the human complement C4 promoter[J].JImmunol, 2000,164(1):300-307.
[4] GUO L,HUANG J X,LIU Y,et al.Transactivation of Atg4b by C/EBP beta promotes autophagy to facilitate adipogenesis[J].MolCellBiol,2013,33(16):3180-3190.
[5] HIMEDA C L,RANISH J A,PEARSON R C,et al.KLF3 regulates muscle-specific gene expression and synergizes with serum response factor on KLF binding sites[J].MolCellBiol,2010,30(14):3430-3443.
[6] SUE N,JACK B H,EATON S A,et al.Targeted disruption of the basic Krüppel-like factor gene(Klf3) reveals a role in adipogenesis[J].MolCellBiol,2008,28(12):3967-3978.
[7] BELL-ANDERSON K S,FUNNELL A P,WILLIAMS H,et al.Loss of Krüppel-like factor 3(KLF3/BKLF) leads to upregulation of the insulin-sensitizing factor adipolin(FAM132A/CTRP12/C1qdc2)[J].Diabetes,2013,62(8):2728-2737.
[8] ZHANG J,YANG C,BREY C,et al.Mutation in Caenorhabditis elegans Krüppel-like factor,KLF-3 results in fat accumulation and alters fatty acid composition[J].ExpCellRes,2009,315(15):2568-2580.
[9] ZHANG J,HASHMI S,CHEEMA F,et al.Regulation of lipoprotein assembly,secretion and fatty acid beta-oxidation by Krüppel-like transcription factor,klf-3[J].JMolBiol,2013,425(15):2641-2655.
[10] GUO L,SUN B,SHANG Z,et al.Comparison of adipose tissue cellularity in chicken lines divergently selected for fatness[J].PoultSci,2011,90(9):2024-2034.
[11] ZHANG Z,WANG H,SUN Y,et al.Klf7 modulates the differentiation and proliferation of chicken preadipocyte[J].ActaBiochimBiophysSin(Shanghai),2013,45(4):280-288.
[12] ROSEN E D,MACDOUGALD O A.Adipocyte differentiation from the inside out[J].NatRevMolCellBiol,2006,7(12):885-896.
[13] LEFTEROVA M I,LAZAR M A.New developments in adipogenesis[J].TrendsEndocrinolMetab,2009,20(3):107-114.
[14] FARMER S R.Transcriptional control of adipocyte formation[J].CellMetab,2006,4(4):263-273.
[15] WANG Y,MU Y,LI H,et al.Peroxisome proliferator-activated receptor-gamma gene:a key regulator of adipocyte differentiation in chickens[J].PoultSci,2008,87(2):226-232.
[16] LIU S,WANG Y,WANG L,et al.Transdifferentiation of fibroblasts into adipocyte-like cells by chicken adipogenic transcription factors[J].CompBiochemPhysiolAMolIntegrPhysiol,2010,156(4):502-508.
[17] LINHART H G,ISHIMURA-OKA K,DEMAYO F,et al.C/EBPalpha is required for differentiation of white,but not brown,adipose tissue[J].ProcNatlAcadSciUSA,2001,98(22):12532-12537.
[18] EL-JACK A K,HAMM J K,PILCH P F,et al.Reconstitution of insulin-sensitive glucose transport in fibroblasts requires expression of both PPARgamma and C/EBP alpha[J].JBiolChem,1999,274(12):7946-7951.
[19] LIN F T,LANE M D.CCAAT/enhancer binding protein alpha is sufficient to initiate the 3T3-L1 adipocyte differentiation program[J].ProcNatlAcadSciUSA,1994,91(19):8757-8761.
[20] MACDOUGALD O A,HWANG C S,FAN H,et al.Regulated expression of the obese gene product(leptin) in white adipose tissue and 3T3-L1 adipocytes[J].ProcNatlAcadSciUSA,1995,92(20):9034-9037.
[21] SMITH S,WITKOWSKI A,JOSHI A K.Structural and functional organization of the animal fatty acid synthase[J].ProgLipidRes,2003,42(4):289-317.
[22] CHIRALA S S,WAKIL S J.Structure and function of animal fatty acid synthase[J].Lipids,2004,39(11):1045-1053.
(编辑 郭云雁)
Expression Pattern of Chicken Krüppel-like Factor 3 Gene and Its Effect on Adipocyte Differentiation
WANG Hai-xia1,ZHANG Zhi-wei1,2,HE Qi1,WANG Ning1, WANG Yu-xiang1,CAO Zhi-ping1,LI Hui1*
(1.KeyLaboratoryofChickenGeneticsandBreedingofMinistryofAgriculture,KeyLaboratoryofAnimalGenetics,BreedingandReproductionatEducationDepartmentofHeilongjiangProvince,CollegeofAnimalScienceandTechnology,NortheastAgriculturalUniversity,Harbin150030,China; 2.SchoolofMedicine,ShiheziUniversity,Shihezi832000,China)
This experiment was conducted to study the expression pattern of Krüppel-like factor 3(GallusgallusKLF3,gKLF3) gene,and its effect on adipocyte differentiation.The expression pattern ofgKLF3 in the adipose tissue and adipocytes of broilers were analyzed by using qRT-PCR.The effect ofgKLF3 on adipocyte differentiation was analyzed by over-expression technology.The result showed thatgKLF3 was consecutively expressed during the chicken adipose tissue development from 2- to 10-week-old.At 10-week-old,thegKLF3 expression in the abdominal fat tissues was significantly higher in fat line chicken than that in lean line chicken(P<0.05).Additionally,the significantly highergKLF3 expression level was observed in preadipocytes in contrast to that in mature adipocytes(P<0.01).The expression levels ofgKLF3 in preadipocytes induced by oleate were lower than that in control.Moreover,over-expression ofgKLF3 inhibited adipocyte differentiation and down-regulated expression ofFASandC/EBPα(P<0.01).The result indicate thatgKLF3 play an important role in the growth and development of chicken abdominal adipose tissue,and might restrain adipocyte differentiation by inhibiting the expression ofC/EBPαandFAS.
chicken;KLF3 gene;adipocyte differentiation;gene expression
10.11843/j.issn.0366-6964.2015.01.004
2014-05-07
国家高技术研究发展计划(863计划)(2013AA102501);国家肉鸡产业技术体系建设项目(CARS-42);黑龙江省高等学校科技创新团队建设项目(2010td02)
王海霞(1987-),女,山东烟台人,硕士生,主要从事家禽遗传育种研究,E-mail:haixiawang1025@126.com
*通信作者:李 辉,教授,E-mail:lihui@neau.edu.cn
S831.2
A
0366-6964(2015)01-0026-06
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28
特产研究(2021年6期)2021-12-08
畜禽业(2021年11期)2021-11-22
中老年保健(2021年9期)2021-08-24
昆明医科大学学报(2021年8期)2021-08-13
新农业(2020年18期)2021-01-07
河南农业科学(2019年9期)2019-09-24
农村百事通(2016年6期)2016-05-14
中国民族医药杂志(2016年4期)2016-05-09
中国卫生标准管理(2015年25期)2016-01-14
