
极端粒形水稻品种GS5基因的序列及效应分析
2015-03-15 12:32梁彦丽张亚东王才林
江苏农业学报 2015年5期
郑 佳, 丁 丹,2, 梁彦丽,2, 张亚东, 王才林
(1.江苏省农业科学院粮食作物研究所/江苏省优质水稻工程技术研究中心/国家水稻改良中心南京分中心,江苏 南京210014;2.南京农业大学农学院,江苏 南京 210095)
水稻是中国人赖以生存的重要粮食作物,提高水稻产量一直以来都是科研人员研究的重点之一,矮化育种和杂交育种这两大历史性的科学研究成果使水稻产量在上世纪得到了较大幅度的提高[1-3]。随着分子生物学日新月异的发展,研究者们开始从分子生物学这一角度思考如何提高水稻产量。水稻的产量主要取决于每株水稻的穗数,每穗的谷粒数以及粒质量,而粒质量主要由粒型决定[4]。
目前利用分子生物学手段定位和克隆到一些与粒型密切相关的基因如 GW2[5]、qSW5[6]/GW5[7-8]、GS3[9-10]、GS5[11]、GW8[12]和 qGL3[13]/GL3.1[14]等。GS5 是控制水稻粒宽、粒质量和充实度的主效QTL。Li等利用 zhenshan97和H94的杂交种构建的单倍体加倍性群体发现位于第5条染色体短臂上的GS5基因。通过形态学比较不同等位基因构成的近等基因系的水稻发现拥有大粒单倍型GS5基因的水稻幼穗的内稃,外稃在横切面上拥有更多的细胞数目。与野生型相比,过表达大粒型GS5基因的植株在细胞周期G1/S阶段的一些相关基因表达量上调。这些证据证明GS5作为细胞周期基因的上游正调控因子,其高表达量可以通过调节有丝分裂行为来增加细胞数目,从而控制籽粒大小。通过分析GS5基因ORF区和启动子区域的序列发现位于启动子区域的序列多态性是对粒形产生影响的原因,而非其编码区[11]。
本研究以江苏省农业科学院粮食作物研究所杂交籼稻项目组选育获得的1个新的特大粒粳稻材料TD70和小粒籼稻Kasalath为研究对象,对2个水稻品种的GS5基因的基因组序列和cDNA序列进行克隆和分析,并设计1对SSR分子标记,对2个亲本及来源于这2个亲本的240个重组自交系(RILs)群体的GS5基因型和表型进行检测,为明确粒型控制基因GS5在2种极端粒型水稻中的序列差异和作用奠定基础。
1 材料与方法
1.1 研究材料
来源于天鹅谷///9520//(72-496/御糯)的特大粒粳稻TD70和小粒籼稻Kasalath,其粒宽2年平均分别为4.42 cm和2.55 cm;TD70/Kasalath的240个重组自交系(RILs)。用游标卡尺(精确到0.01 mm)测量240个RILs及其亲本的单个籽粒长度。每个品种或株系随机取5个单株,每株随机选取10粒饱满籽粒测定,取平均值作为粒宽的表型值。
1.2 植物DNA、RNA提取及cDNA合成
水稻DNA提取以新鲜叶片为材料,总RNA提取以生长旺盛期的根、茎、幼穗的混合样为材料,DNA、RNA提取试剂盒采用 OMEGA 公司的 Plant DNA Kit和 Plant RNA Kit,提取的RNA用DNa se I处理样本,琼脂糖凝胶电泳和分光光度计检测RNA质量后进行第一链c DNA的合成。第一链cDNA合成试剂盒采用Fermentas公司RevertAid First Strand cDNA synthesis kit,胶回收采用 Axygen公司的胶回收试剂盒。
1.3 引物设计、合成与序列测定
特异性引物利用Primer Premier 5.0软件设计,并由英潍捷基贸易有限公司合成,序列测定由英潍捷基贸易有限公司完成。
1.4 GS5基因的DNA序列的克隆
以提取的TD70和Kasalath基因组为模版,特异性基因GS5G1F、GS5G1R、GS5G2F和GS5G2R扩增引物通过聚合酶链式反应分段扩增GS5基因,测序结果利用DNAman进行拼接后即为完整的GS5基因DNA序列。以提取的 TD70和 Kasalath的总RNA为模版,反转录为cDNA后利用特异性扩增引物GS5F、GS5R通过聚合酶链式反应扩增GS5基因的cDNA全长。
1.5 PCR产物的克隆、纯化与筛选
PCR产物在1%的琼脂糖凝胶中电泳,确定片段大小正确无误后,切胶回收,纯化。连接到pGEM-T克隆载体上,热激法导入大肠杆菌DH5α感受态细胞中,进行氨苄抗性法和X-gal/IPTG蓝白斑筛选,挑取阳性菌落在含有氨苄的培养基中培养后进行菌液PCR,扩增程序为:94℃预变性5 min;94 ℃ 1 min,55 ℃ 1 min,72 ℃ 1 min,30 个循环;72℃5 min。电泳后将检测为阳性的菌落送公司测序,约10个重复样。
2 结果
2.1 GS5基因DNA、cDNA全长克隆与序列分析
已知的GS5基因(Os05g0158500)序列长4 466 bp,分段设计扩增引物 GS5G1F:5'-CTCCCATGGAATTACTAGAGAA-3',GS5G1R:5'-TTGGACATGTTTACATCCACAT-3';GS5G2F: 5'-TGTATGGTCTAACATTCAGAATTC-3',GS5G2R:5'-GCGCACTTGAAATTGATTTG-3'。利用 NCBI的 Primerblast检测引物对在水稻中仅可以扩出1个片段。利用引物对在TD70和Kasalath的全基因组中扩增出的片段测序拼接后发现TD70品种中该片段TGS5g长度为 4 449 bp,Kasalath品种中该片段KGS5g长 度 为 4 441 bp,它 们 与 已 知 GS5(Os05g0158500)基因的序列相似性达99.82%。
利用已扩增出的基因组序列结合NCBI上下载的已知GS5基因序列信息,设计cDNA扩增引物GS5F: 5'-ATGGCGGTGGCGG-3',GS5R: 5'-TCATCTGCTTGTCGGAAG-3',在 TD70 和Kasalath反转录的cDNA模版上分别扩增出1条约1 500 bp大小的片段。测序结果显示,TD70品种中该片段TGS5长度为1 449 bp,编码482个氨基酸残基,Kasalath品种中该片段KGS5长度为1 443 bp,编码480个氨基酸残基,它们与已知 GS5(Os05g0158500)序列相似性达99.84%。TGS5和Os05g0158500的开放阅读框在碱基的+92位是G,因此第31位编码的氨基酸为甘氨酸;KGS5的开放阅读框在碱基的+92位是C,该位点编码的氨基酸为丙氨酸,且在+106~111 bp缺失GGCGGC6个碱基,即第36、37位编码的氨基酸缺失2个甘氨酸(图1)。
比较GS5基因的DNA序列和cDNA序列发现TGS5和KGS5基因在基因结构上包含10个外显子和9个内含子(图2)。
2.2 GS5基因分子标记设计及验证
分析TD70和Kasalath的GS5基因序列上的多处差异,根据KGS5基因第一外显子中GCGGCG的缺失设计SSR(Simple sequence repeats)标记,正向引物GS5SSR-F:5'-AGGCGGCGAGATGTCTCTTGTC-3',反 向 引 物 GS5SSR-R:5'-CTGCGCCTCGAAGAACCAGTAG-3'(图3)。该引物在 TD70中可以扩增出216 bp的条带,在Kasalath中可以扩增出210 bp的条带(图4)。
2.3 功能标记对TD70/Kasalath RIL群体的鉴定
利用上述分子标记检测TD70与Kasalath为亲本构建的重组自交系群体的结果显示,240个株系中含有GS5-T基因(表现TGS5基因型)的株系有122个,含有GS5-K(表现KGS5基因型)基因的株系有118个。含GS5-T基因的株系平均粒宽为 (3.26±0.37)mm,平均千粒质量为 (32.61±7.66)g,含GS5-K基因的株系平均粒宽为 (2.98±0.28)mm,平均千粒质量为 (29.74±6.36)g,两者分别相差0.47 mm和2.87 g,粒宽存在极显著差异(表1),说明GS5是水稻粒宽性状的主效调控基因。此外,GS5对水稻的粒长、粒厚都有一定的影响,具有多效调节作用。
2.4 GS5基因对水稻粒宽的作用及与已知其他基因的比较
GW2基因是控制水稻粒宽的主效基因,某些大粒型水稻品种中存在的功能缺失型是籽粒变宽的重要调节因素,我们之前的研究已经证实本试验所用研究材料TD70品种拥有稀有的GW2功能缺失型,是TD70的超宽籽粒的正向调控因子[15],且在TD70和 Kasalath构建的重组自交系中,GW2对粒宽的贡献较大,含GW2-T基因的株系平均粒宽比含GW2-K基因的株系平均粒宽分别高出0.47~0.51 mm,千粒质量高出5.54~6.05 g[15]。本试验发现同一重组自交系群体中,含GS5-T基因的株系平均粒宽比含GS5-K基因的株系平均粒宽高出0.47 mm,千粒质量高出2.87 g。说明GS5对粒宽调控效应与GW2相当,但是对千粒质量的调控效应没有GW2强。

图1 TD70品种中GS5基因片段及预测编码的氨基酸Fig.1 GS5 sequence fragment and encoded protein prediction in TD70

图2 TGS5和KGS5的基因结构Fig.2 Structure of TGS5 and KGS5 genes
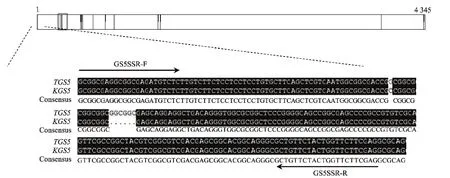
图3 TGS5和KGS5差异位点及引物序列所处的位置Fig.3 The polymorphic loci and primers locations of TGS5 and KGS5

图4 GS5分子标记对TD70、Kasalath和RIL群体中的扩增结果Fig.4 PCR amplification of GS5 with SSR marker in TD70,Kasalath and RILs

表1 含不同来源的GS5基因的株系的粒型表现Table 1 Grain phenotype of lines with different GS5 genotypes
3 讨论
GS5是控制水稻粒宽、粒质量和充实度的主效QTL。已有研究结果表明GS5基因启动子区域的序列多态性是影响水稻粒宽的原因,而非其编码区[11]。本试验通过对极端粒型水稻TD70和Kasalath 2个品种的GS5基因的研究并未发现文献报道的启动子区域的差异,但是编码区存在GGCGGC的插入缺失位点。宽粒TD70型的GS5基因较窄粒Kasalath型GS5基因所编码的氨基酸序列中存在1个甘氨酸向丙氨酸的变化和2个丙氨酸的增加。本研究利用该差异位点成功设计区分2种类型GS5基因的SSR标记,对重组自交系群体分型后发现TD70型的GS5基因对水稻粒宽具有显著的正调控作用,效应与GW2相当,对粒宽、粒厚和千粒质量也有一定的正向调控效应。在高粱、玉米等其他禾本科植物中也发现了水稻GS5基因的直系同源基因,有些物种内还存在侧向同源基因 ,说明GS5基因对籽粒的调控作用并不单一存在于水稻中。
[1] XINGY Z,ZHANGQF.Genetic and molecular bases of rice yield[J].Annu Rev Plant Biol,2010,61:421-442.
[2] 付立东,隋 鑫,任 海,等.不同取秧量与穴距对机插水稻产量的影响[J].江苏农业科学,2014,42(12):70-72.
[3] 唐 成,陈 露,安敏敏,等.稻瘟病诱导水稻幼苗叶片氧化还原系统的特征谱变化[J].江苏农业科学,2014,42(12):141-144.
[4] XING Z,TAN F,HUA P,et al.Characterization of the main effects,epistatic effects and their environmental interactions of QTLs on the genetic basis of yield traits in rice[J].Theor Appl Genet,2002,105(2-3):248-257.
[5] SONG X J,HUANGW,SHIM,et al.A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase[J].Nat Genet,2007,39(5):623-630.
[6] SHOMURA A,IZAWA T,EBANA K,et al.Deletion in a gene associated with grain size increased yields during rice domestication[J].Nat Genet,2008,40:1023-1028.
[7] WAN X,WENGJ,ZHAIH,et al.Quantitative trait loci(QTL)analysis for rice grain width and fine mapping of an identified QTL allele gw-5 in arecombination hotspot region on chromosome5[J].Genetics,2008,179(4):2239-2252.
[8] WENG J,GU S,WAN X,et al.Isolation and initial characterization of GW5,a major QTL associated with rice grain width and weight[J].Cell Res,2008,18(12):1199-1209.
[9] FAN C C,XING Y Z,MAO H L,et al.GS3,a major QTL for grain length and weight and minor QTL for grain width and thickness in rice,encodes a putative transmembrane protein[J].Theor Appl Genet,2006,112(6):1164-1171.
[10] MAO H,SUN S,YAOJ,et al.Linking differential domain functions of the GS3 protein to natural variation of grain size in rice[J].Proc Natl Acad Sci USA,2010,107:19579-19584.
[11] LI Y,FAN C,XING Y,et al.Natural variation in GS5 plays an important role in regulating grain size and yield in rice[J].Nat Genet,2011,43:1266-1269.
[12] WANG S,WU K,YUAN Q,et al.Control of grain size,shape and quality by OsSPL16 in rice[J].Nat Genet,2012,44(8):950-954.
[13] ZHANG X,WANG J,HUANG J,et al.Rare allele of OsPPKL1 associated with grain length causes extra-large grain and a significant yield increase in rice[J].Proc Natl Acad Sci USA,2012,109(52):21534-21539.
[14] QI P,LIN Y S,SONGX J,et al.The novel quantitative trait locus GL3.1 controls rice grain size and yield by regulating Cyclin-T1 [J].Cell Research,2012,22:1666-1680.
[15]张亚东,梁彦丽,郑 佳,等.极端粒形水稻粒宽基因 GW2的序列分析和效应[J].中国水稻科学,2014,28(6):581-588.
[16]张 帅,王金朋,张 宪,等.禾本科植物中调控产率基因 GS5的进化[J].河北联合大学学报:自然科学版,2012,34(4):65-68.
猜你喜欢
亚热带植物科学(2022年1期)2022-05-17
江苏农业学报(2020年2期)2020-05-27
四川农业科技(2019年5期)2019-07-01
北京农学院学报(2019年1期)2019-02-22
山东农业科学(2018年11期)2018-01-17
河南农业科学(2017年4期)2017-04-12
江苏农业科学(2016年6期)2016-07-25
西南农业学报(2016年5期)2016-05-17
西南农业学报(2016年6期)2016-04-16
浙江柑橘(2016年1期)2016-03-11
