
基于CitespaceⅡ软件猫杯状病毒研究领域演进路径、研究热点及前沿的分析
2015-03-09 05:40赵艳丽董宏伟宋学洲孙宁宁杨松涛胡桂学
中国预防兽医学报 2015年9期
赵艳丽,董宏伟,张 鑫,宋学洲,孙宁宁,杨松涛,胡桂学
(1.吉林农业大学 动物科学技术学院,吉林 长春 130118;2.吉林农业大学 图书馆,吉林 长春 130118;3.吉林省农业广播电视学校,吉林 长春 130021;4.东北大学图书馆,辽宁 沈阳 110819;5.军事医学科学院 军事兽医研究所,吉林 长春 130062)
猫杯状病毒(Feline calicivirus,FCV)属于杯状病毒科水疮性病毒属的成员,为单股正链RNA 病毒,是杯状病毒科中少有的可以在体外培养的病毒。FCV 能够引起猫多发性口腔和呼吸道疾病,其发病率较高,几乎所有的猫科动物均易感,该病毒高度变异,其变异株还可以引起猫全身性致命性疾病,死亡率增加。CitespaceⅡ是由美国Drexel 大学开发的一款知识图谱可视化软件[1]。利用该软件的综合统计学分析功能可以显示出某一个病原学研究等领域发展的过程,特别是相关研究的知识转折点和关键点,而且能够探测一个领域的研究热点和前沿。近年来,随着大数据时代的到来,进一步掀起了综合统计学分析的浪潮,CitespaceⅡ软件有助于科研工作者快速地了解某一个研究领域最新成果及发展趋势。本研究主要应用CitespaceⅡ软件来展示FCV 研究领域的知识特征。
1 数据来源与方法
以FCV 为例,其相关数据来源为Science Citation Index-expanded(SCI-E),检索式为:主题="feline calicivirus",检索时间跨度为所有年份,检索结果为792 条,数据采集时间为2012 年11 月9 日。采用Citespace II 3.7.R7(64-bit)软件,选择最小生成树(Minimum spanning tree)算法,设定时间间隔为2 年,主题词来源选择标题、摘要和关键词,引文值、共被引值和共被引系数阈值分别设置为(2,3,15)、(3,3,20)、(3,3,20),运行Citespace II软件,对国家机构、作者共被引(ACA)、文献共被引(DCA)和关键词共现进行统计学分析。
2 FCV 研究领域统计学分析
2.1 国家和机构分析 在Citespace II 软件界面,网络节点选择“Country”和“Institution”,运行软件,显示FCV研究的重点国家和机构,发现该领域研究的国家和机构分布较集中,其中发文量最多的是美国,其次为英格兰、日本、德国、加拿大,发文最多的机构为加利福尼亚大学戴维斯分校(表1),而国内却鲜有报道,发文量少于10 篇。
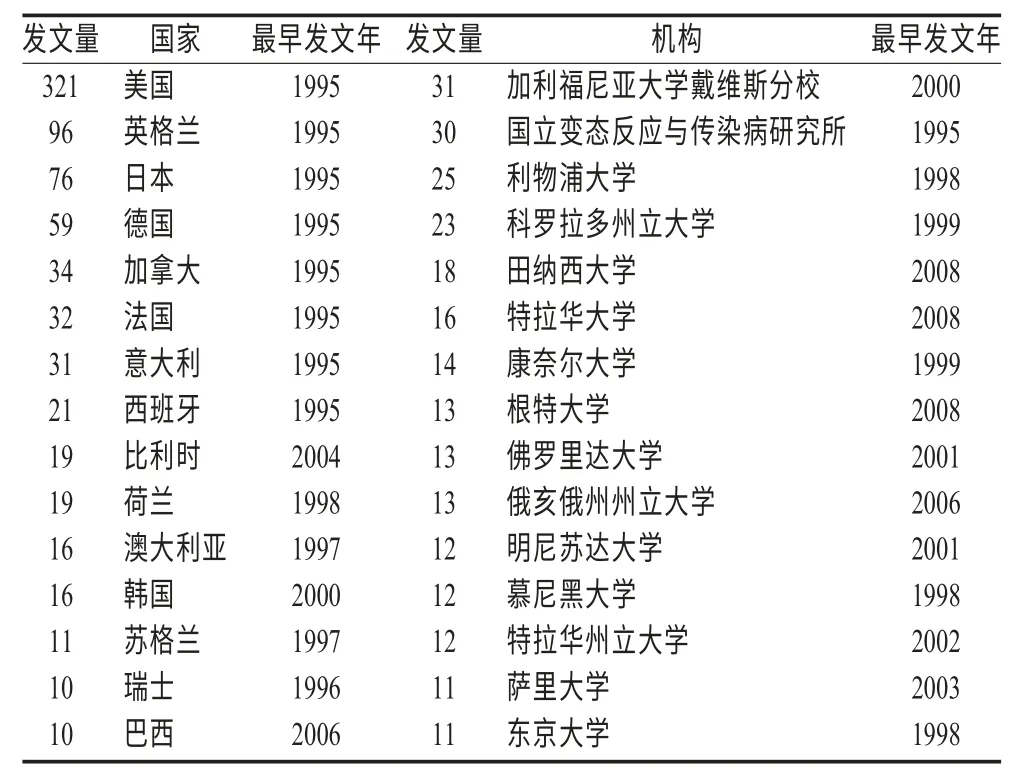
表1 FCV 研究领域国家、机构分布
2.2 作者共被引分析(ACA)在Citespace II 软件界面,网络节点选择“Cited author”,运行软件,进行聚类,发现FCV 研究作者分为4 个聚类,Hendrick MJ、Esplin DG为聚类#0 的成员,聚类标签为Postvaccinal;Pedersen NC、Radford AD、Dawson S、Povey RC 等为聚类#1 的高影响力作者,聚类标签为Feline;Carter MJ、Neill JD、Sosnovtsev SV、Jiang X 等为聚类#2 的高影响力作者,聚类标签为Organazation;Duizer E、Wobus CE、Doultree JC、Cannon JL 等为聚类#3 的高影响力作者,聚类标签为Norovirus。以下是具体的高被引频次、高突现值、高中心度作者:一个聚类向另一个聚类过渡的关键作者(表2)。
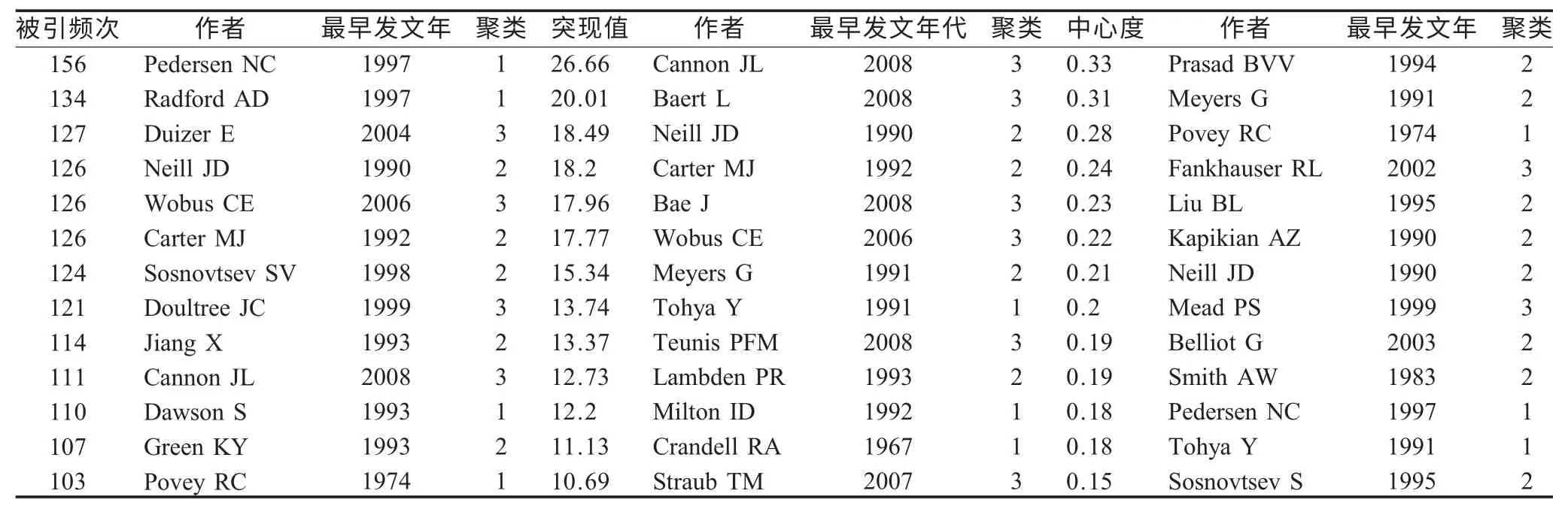
表2 高影响力作者
2.3 演进路径 根据软件的定义,文献共被引可视化图谱中的关键节点是图谱中连接两个以上不同聚类,并且相对中心度和被引频次较高的节点,这些节点可能成为网络中由一个时间段向另一个时间段过渡的关键节点,关键节点往往使研究领域产生变革,在领域知识发展过程中起承上启下的作用。因此,可以通过关键节点探知研究领域的演进路径。陈超美博士将关键节点定义为中心度大于0.1的节点[1],以下是中心度在0.1 以上的文献(表3),它们是整个网络关键节点文献,可以认为对FCV 领域的研究与发展起奠基性作用。
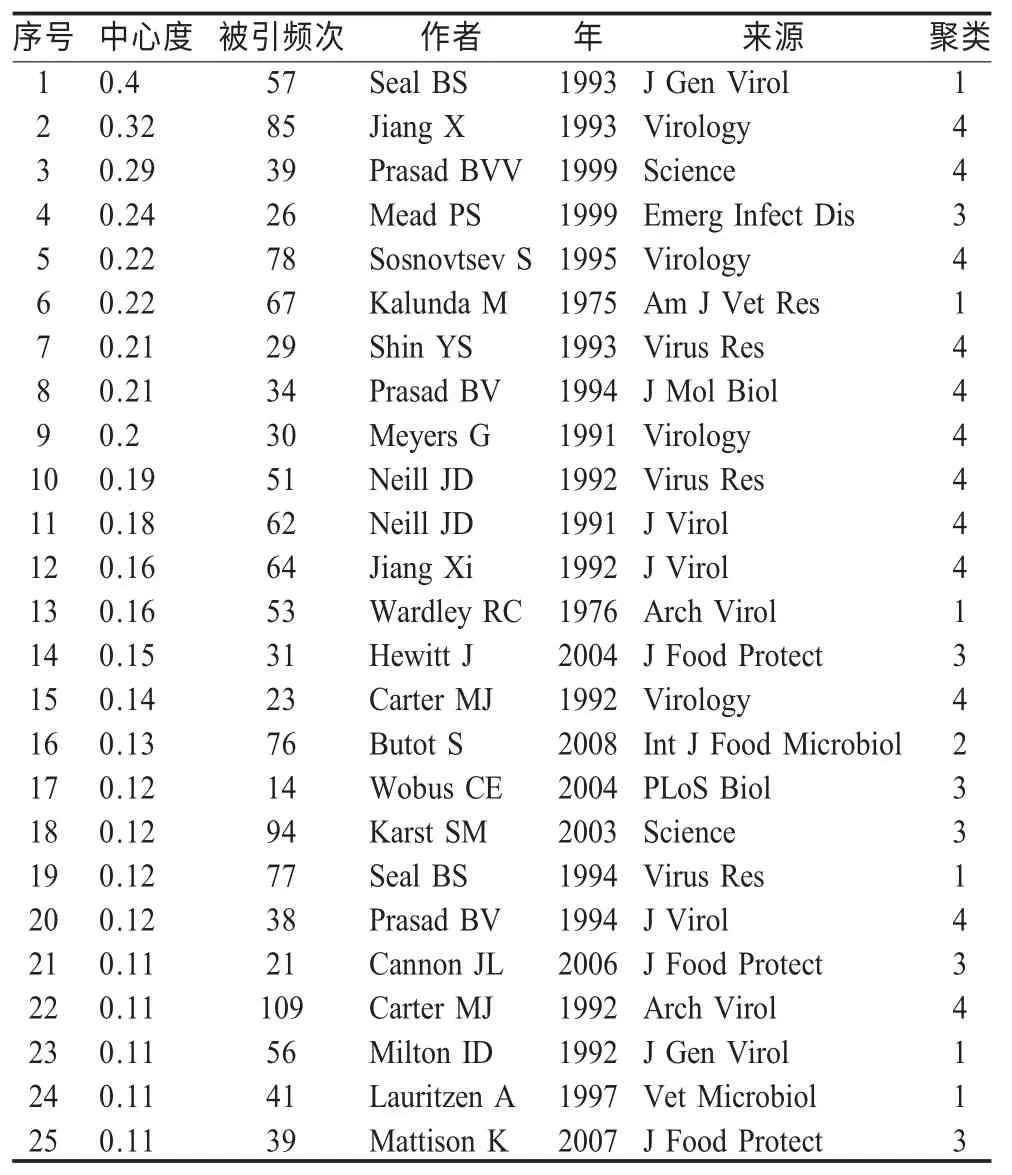
表3 关键节点文献(中心度>0.1)
1975 年:Kalunda 等提出F9 株可能成为参考株,并且FCV 只具有一个血清型。1976 年:Wardley 发现FCV的传播途径。1991 年:Neill 等报道了FCV 衣壳蛋白基因的核酸序列与表达;Meyers 等报道了RHDV 基因组的分子克隆和核苷酸序列。1992 年:Carter 等报道了FCV 的全基因序列,衣壳蛋白的识别和序列决定,确定了成熟的衣壳蛋白和衣壳蛋白前体的切割位置;Jiang 等报道了诺如病毒(NoV)衣壳蛋白的表达、自我组装和抗原性;Neill等报道了杯状病毒科成员圣米格尔海狮病毒的两个血清型的衣壳蛋白核苷酸序列,识别了衣壳蛋白中保守和非保守氨基酸序列;Milton 等报道了FCV 衣壳蛋白3 个单克隆抗体(MAb)连接位点的位置,一个非中和MAb 识别C 末端36 个氨基酸内的区域,两个中和MAb 连接在蛋白中心37 个氨基酸的不同区域,分析表明该区域是衣壳蛋白主要的可变区。1993 年:Seal 等识别了FCV 衣壳蛋白可变抗原决定区,Jiang 等报道了NoV 的序列和基因组结构,确定其属于杯状病毒科家族的成员;Shin 等研究发现3 个中和表位具有一个构象的特征,其它的4 个线性表位在蛋白的381 和454 间存在高变区。1994 年:Prasad 等报道了杯状病毒的三维结构;Seal 报道了不同FCV 分离株间衣壳蛋白基因变异分析;Prasad 等报道了杆状病毒表达NoV衣壳的三维结构。1995 年:Sosnovtsev 开启了FCV 反向遗传学研究[2]。1997 年:Lauritzen 等报道了分离自美国和英国的FCV 毒株血清学分析结果[3]。1999 年:Mead 等报道了美国与食物相关的疾病和死亡,与以前的估量相比,食源性疾病能够引起更多的疾病但更少的死亡[4];Prasad等报道了NoV 衣壳蛋白X-射线的晶体结构[5]。2003 年:Karst 等研究发现对NoV 抗性至关重要的是依赖先天性免疫的信号转导因子和转录因子1 激活剂,而不是依赖适应性免疫的T、B 细胞。对鼠诺如病毒(MNV)抗性至关重要的宿主分子的识别可能为一个重要的人类NoV 的预防或控制提供目标[6]。2004 年:Wobus 等首次报道了MNV-1在细胞培养中复制,揭示了其对树突状细胞和核巨噬细胞的嗜性,提供了NoV 重要的生物学特性[7];Hewitt 等报道了NoV、甲型肝炎病毒(HAV)、FCV 在腌制的贻贝中的存活和持续感染[8]。2006 年:Cannon 发现MNV 可以在细胞中培养,确定其有望成为研究人类NoV 存活环境的替代病毒[9]。2007 年:Mattison 等报道了在代表性食品和金属表面上杯状病毒的生存:FCV 作为一个NoV 的替代实验,研究数据为流行病学的监测以及食品加工实践的发展,灭活NoV 合适的策略,控制它通过食物和表面传播提供有价值的信息[10]。2008 年:Butot 等报道了在浆果和草本植物中卫生、冷冻和冷藏储存对肠道病毒的影响[11]。FCV 研究领域知识演进路径(图1)。
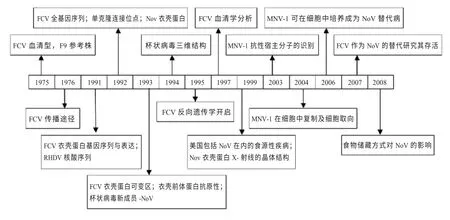
图1 FCV 研究领域知识演进路径
2.4 FCV 领域的研究热点分析
2.4.1 关键词共现网络 文献的关键词是作者依据文章内容高度概括产生的,虽然数量不多,但其出现的频次在一定程度上能够反映出某领域研究的热点。在Citespace II软件界面,网络节点选择“keywords”,运行软件。
将相关词进行合并,发现该领域的研究热点如下:FCV、NoV、杯状病毒、灭活、猫、胃肠炎、识别、甲型肝炎病毒、RT-PCR、核苷酸序列、疾病、MNV、暴发、美国、诺瓦克样病毒、感染、出血性疾病病毒、表达、肠道病毒、流行、暴发、替代物、衣壳蛋白基因(表4)。
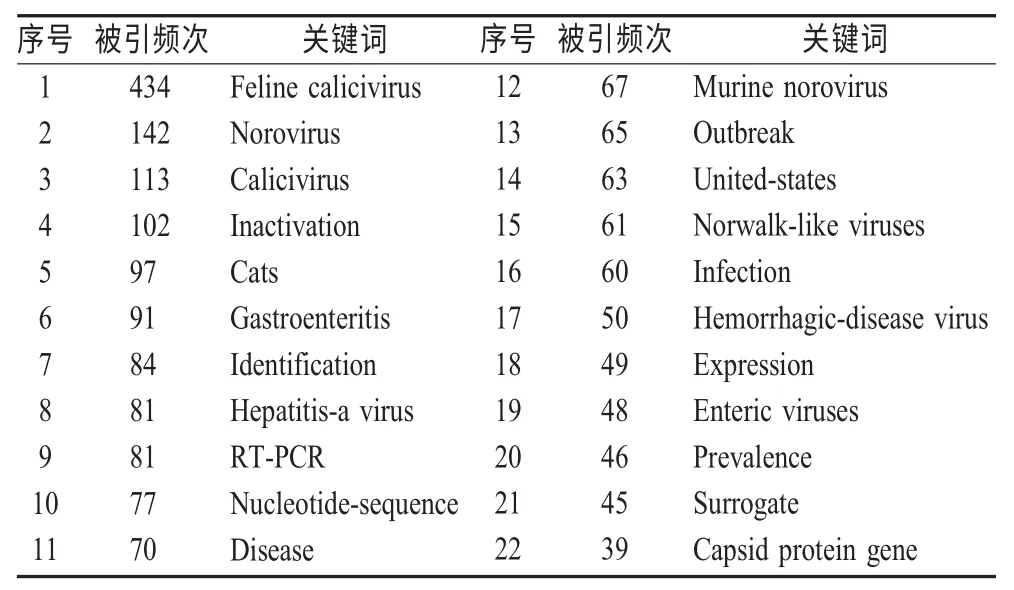
表4 高被引关键词列表
2.4.2 高被引文献内容分析 从科学计量学和知识计量学的角度,一段时间内被引频次高的一系列文献可以体现该文献所在研究领域中在这一时间段内的研究热点问题。因此,可以将文献被引频次的排名情况作为揭示该领域研究热点问题的一个指标。在CitespaceⅡ软件界面,网络节点选择“cited reference”,运行CitespaceⅡ软件,进行聚类,用题名词进行标签,下面列出了被引频次≥50 的高影响力文章(表5)。
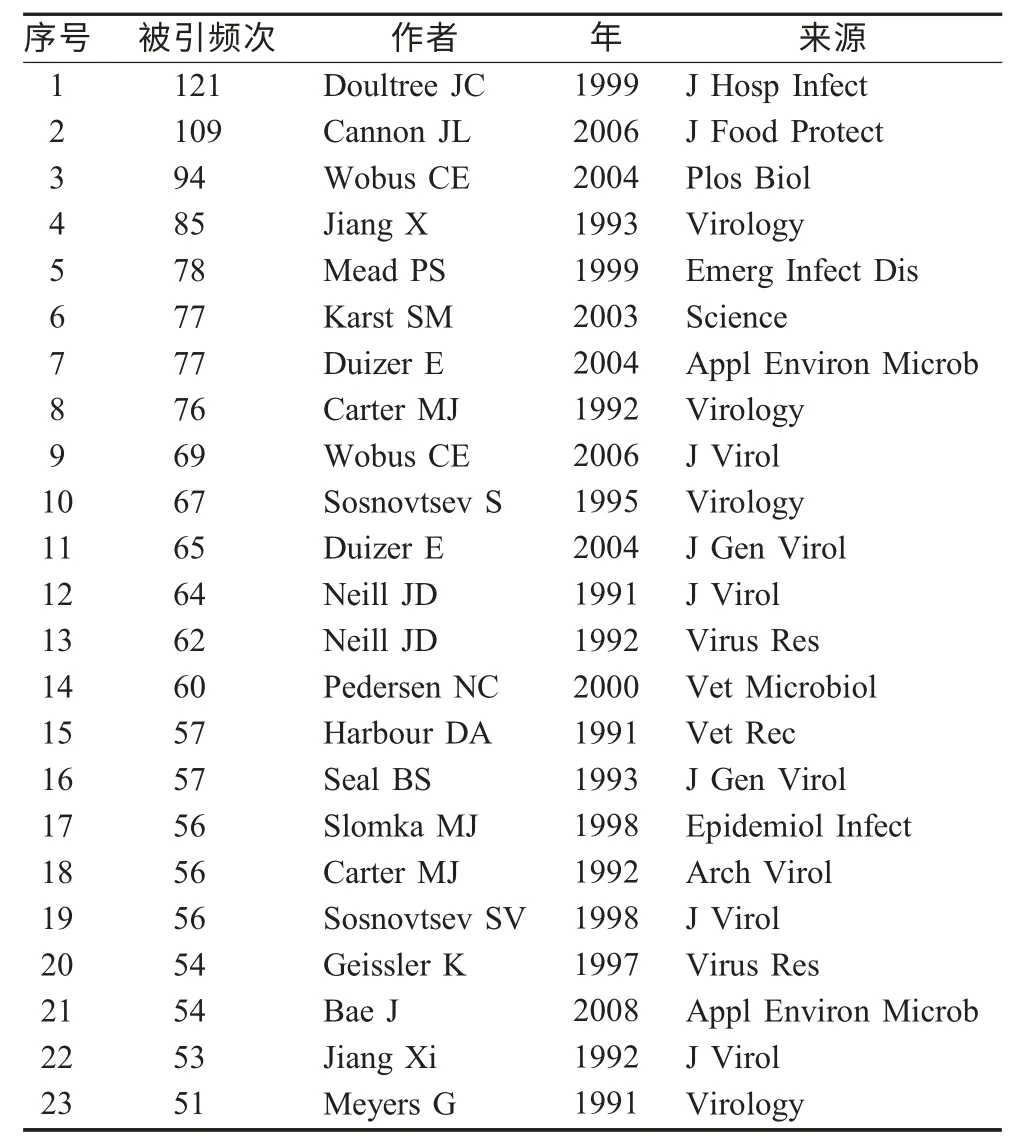
表5 被引频次≥50 的高影响力文章
根据FCV 领域高被引文献的内容分析,归纳出FCV领域研究热点主要有以下聚类:
(1)FCV:基于FCV 自身的研究领域。
FCV 作为研究领域的检索词无疑会成为最大的研究热点,该聚类下包含FCV、猫、核苷酸序列、表达、衣壳蛋白基因等关键词,该聚类的高被引文献如下:
该聚类包含表5 中8、10、12、14、15、16、18、19、20 号高被引文献,其中8、10、12、16、18 号文献前面已述,文献14:报道了一个新的高致病的FCV 株--FCV-Ari 引起猫出血热样孤立流行病[12]。文献15:将1980~1989 年间从6 866 个猫的咽试纸分离到的FCV 和猫疱疹病毒(FHV)的分离率进行回顾性研究。FCV 分离率为19.9 %,FHV 的分离率为4.2 %。文献19:研究发现FCV 衣壳蛋白前体的裂解是由病毒编码的蛋白酶介导[13]。文献20:报道了引起不同疾病表现的FCV 分离株的基因和抗原异质性[14]。
(2)FCV、MNV 作为NoV 替代病毒的相关研究:包括FCV、MNV、NoV、灭活、替代物等关键词。
该聚类包含1、2、3、6、7、9、11、17、21 号高被引文献,其中2、3、6 前面已述,文献1:通过对作为NoV 替代病毒的FCV 进行灭活试验,为建立减少诺瓦克病毒样颗粒(NVLPs)在环境中传播的消毒程序提供一个基础[15]。文献7:为了探究引起食源-水源胃肠炎的NoV,报道了肠道犬杯状病毒No.48(CaCV)和FCV F9(FeCV)和减少FeCV、CaCV 和NoV PCR 单位的相关灭活结果[16]。文献9:报道了一个研究NoV 生物学和发病机制的模型系统:MNV[17]。文献11:通过NoV 体外培养的实验发现试图建立NoV 的培养方法是不成功的[18]。文献17:报道了FCV 作为贝类小的圆的结构病毒的热灭活研究的一个模型系统[19]。文献21:评价了MNV、FCV、脊髓灰质炎病毒和MS2 在表面水和地下水病毒持续感染的模型中作为人NoV 的替代物。发现MNV 依据基因相似和环境稳定性显示了作为人NoV 替代物的巨大潜力,FCV 不太稳定,作为替代物遭到了质疑[20]。
(3)NoV:包含衣壳蛋白、表达、食物、疾病等关键词。该聚类包含4、5、22 号高被引文献,前面已述。
(4)杯状病毒科其它成员:包含出血性疾病病毒、海狮病毒等关键词。
该聚类包含表5 中13、23 号高被引文献,前面已述。
上述各文献的被引频次详见表5。通过热点文献可以看出FCV 领域的研究,除了FCV 感染、传播、基因结构、蛋白结构功能的研究外,对NoV 的研究也比较突出,因为NoV 是感染人类肠道的一个重要的非细菌性病原,对人有一定的危害,由于它不能够在体外培养,很多都是将FCV、MNV 作为其替代病毒进行研究。
2.5 FCV 领域的研究前沿分析 利用Citespace 提供的词频探测技术,通过考察词频的时间分布,将其中频次变化率高的词(Burst term)从大量的关键词中探测出来,依靠词频的变动趋势,而不仅仅是频次的高低,来确定前沿领域和发展趋势。认为突现率(Burst 值)高的关键词可以代表前沿领域。运行Citespace Ⅱ,发现核酸序列、MNV[21-22]、反转录PCR[23-25]、鉴定、出血性疾病病毒、表达、肠病毒、NoV、中和、组织、分子克隆、衣壳蛋白基因、胃肠炎、3C 样蛋白酶[26]、白血病病毒、三维结构、衣壳蛋白[27]、基因组、FCV、免疫缺陷病毒、高变区、裂解位点等突现值高,可以代表该领域的前沿,结合关键词的突现年份可以看出目前的研究前沿为MNV,反转录PCR,肠道病毒,NoV[28-29](表6)。前沿领域主要是围绕引起人肠道胃肠炎的NoV 展开,由于其不能在体外细胞中培养,通过对其替代病毒FCV[30]、MNV[31],尤其是MNV 的核酸、蛋白功能、作用机制等研究,来探究人NoV[32]的致病机理,从而抑制疾病。
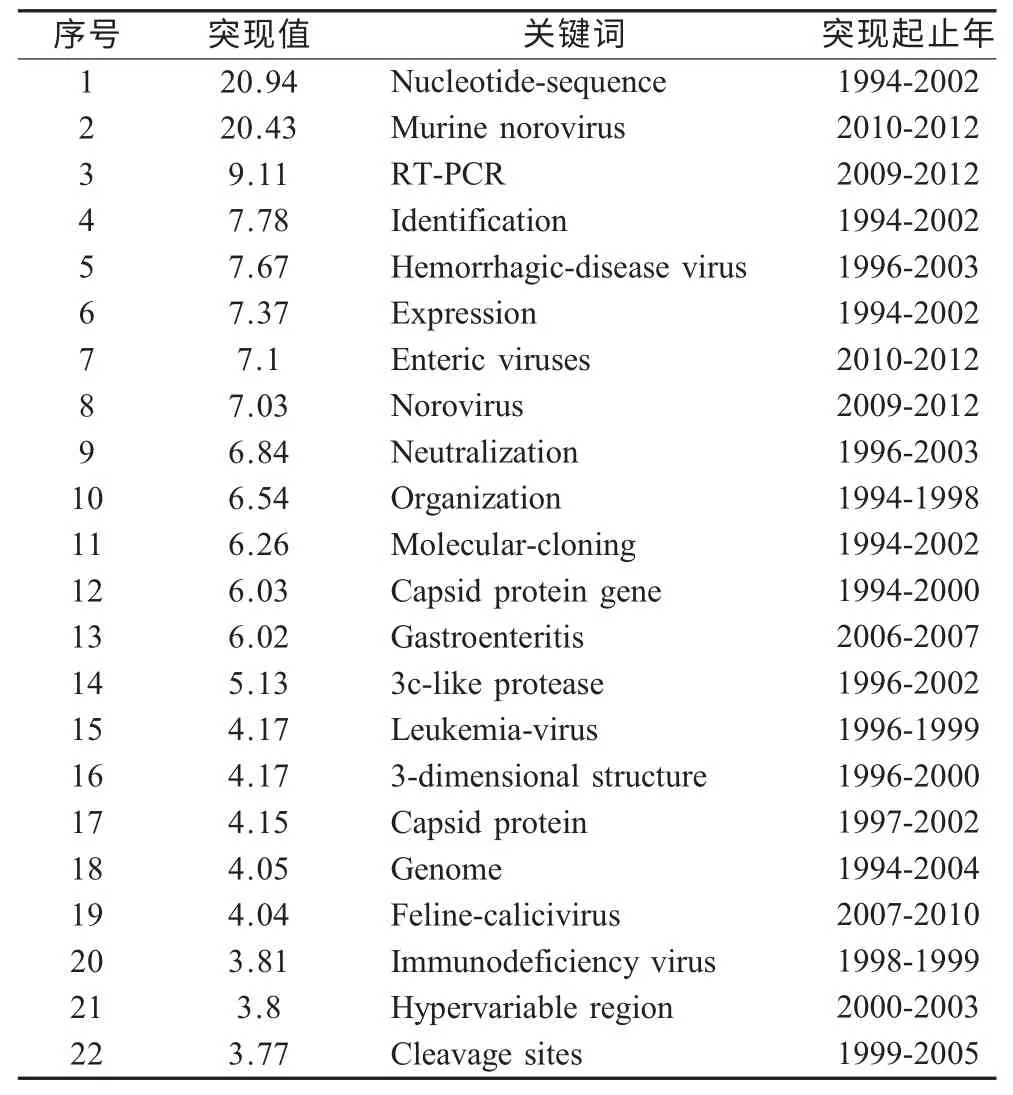
表6 突现值关键词列表
3 结论
CitespaceⅡ是一款可以帮助科研人员迅速地了解某一研究领域的可视化工具。通过CitespaceⅡ知识图谱可视化软件,我们较为全面地探知了FCV 整个研究领域的发展态势,包括其研究的国家、机构、作者、研究基础、研究热点及前沿,望广为传播,助力科研。
[1]Chen Chao-ming.CiteSpace Ⅱ:Detecting and visualizing emerging trends and transient patterns in scientific literature[J].J Am Soc Inf Sci Tec,2006,57(3):359-377.
[2]Sosnovtsev S,Green K Y.RNA transcripts derived from a cloned full-length copy of the feline calicivirus genome do not require VpG for infectivity[J].Virology,1995,210(2):383-390.
[3]Lauritzen A,Jarrett O,Sabara M.Serological analysis of feline calicivirus isolates from the United States and United Kingdom[J].Vet Microbiol,1997,56(1-2):55-63.
[4]Mead P S,Slutsker L,Dietz V,et al.Food-related illness and death in the United States[J].Emerg Infect Dis,1999,5(5):607-625.
[5]Prasad B V,Hardy M E,Dokland T,et al.X-ray crystallographic structure of the Norwalk virus capsid[J].Science,1999,286(5438):287-290.
[6]Karst S M,Wobus C E,Lay M,et al.STAT1-dependent innate immunity to a Norwalk-like virus[J].Science,2003,299(5612):1575-1578.
[7]Wobus C E,Karst S M,Thackray L B,et al.Replication of Norovirus in cell culture reveals a tropism for dendritic cells and macrophages[J].PLoS Biol,2004,2(12):e432.
[8]Hewitt J,Greening G E.Survival and persistence of norovirus,hepatitis A virus,and feline calicivirus in marinated mussels[J].J Food Prot,2004,67(8):1743-1750.
[9]Cannon J L,Papafragkou E,Park G W,et al.Surrogates for the study of norovirus stability and inactivation in the environment:A comparison of murine norovirus and feline calicivirus[J].J Food Prot,2006,69(11):2761-2765.
[10]Mattison K,Karthikeyan K,Abebe M,et al.Survival of calicivirus in foods and on surfaces:experiments with feline calicivirus as a surrogate for norovirus[J].J Food Prot,2007,70(2):500-503.
[11]Butot S,Putallaz T,Sánchez G.Effects of sanitation,freezing and frozen storage on enteric viruses in berries and herbs[J].Int J Food Microbiol,2008,126(1-2):30-35.
[12]Pedersen N C,Elliott J B,Glasgow A,et al.An isolated epizootic of hemorrhagic-like fever in cats caused by a novel and highly virulent strain of feline calicivirus[J].Vet Microbiol,2000,73(4):281-300.
[13]Sosnovtsev S V,Sosnovtseva S A,Green K Y.Cleavage of the feline calicivirus capsid precursor is mediated by a virus-encoded proteinase[J].J Virol,1998,72(4):3051-3059.
[14]Geissler K,Schneider K,Platzer G,et al.Genetic and antigenic heterogeneity among feline calicivirus isolates from distinct disease manifestations[J].Virus Res,1997,48(2):193-206.
[15]Doultree J C,Druce J D,Birch C J,et al.Inactivation of feline calicivirus,a Norwalk virus surrogate[J].J Hosp Infect,1999,41(1):51-57.
[16]Duizer E,Bijkerk P,Rockx B,et al.Inactivation of caliciviruses[J].Appl Environ Microbiol,2004,70(8):4538-4543.
[17]Wobus C E,Thackray L B,Virgin H W.4th Murine norovirus:a model system to study norovirus biology and pathogenesis[J].J Virol,2006,80(11):5104-5112.
[18]Duizer E,Schwab K J,Neill F H,et al.Laboratory efforts to cultivate noroviruses[J].J Gen Virol,2004,85(1):79-87.
[19]Slomka M J,Appleton H.Feline calicivirus as a model system for heat inactivation studies of small round structured viruses in shellfish[J].Epidemiol Infect,1998,121(2):401-407.
[20]Bae J,Schwab K J.Evaluation of murine norovirus,feline calicivirus,poliovirus,and MS2 as surrogates for human norovirus in a model of viral persistence in surface water and groundwater[J].Appl Environ Microbiol,2008,74(2):477-484.
[21]Huang Run-ze,Li Xin-hui,Huang Yao-xin,et al.Strategies toenhance high pressure inactivation of murine norovirus in strawberry puree and on strawberries[J].Int J Food Microbiol,2014,185:1-6.
[22]Fraisse A,Temmam S,Deboosere N,et al.Comparison of chlorine and peroxyacetic-based disinfectant to inactivate feline calicivirus,murine norovirus and hepatitis A virus on lettuce[J].Int J Food Microbiol,2011,151(1):98-104.
[23]Liu Peng-bo,Kim M,Schlesinger D,et al.Immunomagnetic separation combined with RT-qPCR for determining the efficacy of disinfectants against human noroviruses[J].J Infect Public Health,2015,8(2):145-154.
[24]Li Dan,De Keuckelaere A,Uyttendaele M.Application of longrange and binding reverse transcription-quantitative PCR to indicate the viral integrities of noroviruses[J].Appl Environ Microbiol,2014,80(20):6473-6479.
[25]Tong H I,Connell C,Boehm A B,et al.Effective detection of human noroviruses in Hawaiian waters using enhanced RT-PCR methods[J].Water Res,2011,45(18):5837-5848.
[26]Kim Y,Shivanna V,Narayanan S,et al.Broad-spectrum inhibitors against 3C-like proteases of feline coronaviruses and feline caliciviruses[J].J Virol,2015,89(9):4942-4950.
[27]Yao Lin,Li Feng-ling,Wang Lian-zhu,et al.Function of VP2 protein in the stability of the secondary structure of virus-like particles of genogroup II norovirus at different pH levels:function of VP2 protein in the stability of NoV VLPs[J].J Microbiol,2014,52(11):970-975.
[28]Arias A,Emmott E,Vashist S,et al.Progress towards the prevention and treatment of norovirus infections[J].Future Microbiol,2013,8(11):1475-1487.
[29]Hall A J,Eisenbart V G,Etingüe A L,et al.Epidemiology of foodborne norovirus outbreaks,United States,2001-2008[J].Emerg Infect Dis,2012,18(10):1566-1573.
[30]Aboubakr H A,Williams P,Gangal U,et al.Virucidal effect of cold atmospheric gaseous plasma on feline calicivirus,a surrogate for human norovirus[J].Appl Environ Microbiol,2015,81(11):3612-3622.
[31]Royall E,Doyle N,Abdul-Wahab A,et al.Murine norovirus 1(MNV1)replication induces translational control of the host by regulating eIF4E activity during infection[J].J Biol Chem,2015,290(8):4748-4758.
[32]Bozkurt H,D'souza D H,Davidson P M.Thermal inactivation of human norovirus surrogates in spinach and measurement of its uncertainty[J].J Food Prot,2014,77(2):276-283.
猜你喜欢
军事文摘(2022年17期)2022-09-24
解放军医学院学报(2022年1期)2022-04-02
科学导报(2021年82期)2021-12-17
家庭影院技术(2021年8期)2021-11-02
计算机世界(2020年50期)2020-01-15
铁道通信信号(2019年6期)2019-10-08
青年生活(2019年23期)2019-09-10
医药前沿(2018年27期)2018-01-16
雷达学报(2017年6期)2017-03-26
互联网天地(2016年1期)2016-05-04
