
ISG15 蛋白促进猪源牛病毒性腹泻病毒2 型对I 型IFN 表达抑制的研究
2015-03-09 05:40:44廖金虎张信军朱国强
中国预防兽医学报 2015年9期
陶 洁,廖金虎,张 倩,张信军,朱国强
(1.扬州大学 兽医学院,江苏 扬州 225009;2.江苏省动物重要疫病与人兽共患病防控协同创新中心,江苏 扬州 225009;3.上海市农业科学院 畜牧兽医研究所,上海 201106)
猪源牛病毒性腹泻病毒(Bovine viral diarrhea virus,BVDV)的流行日益广泛,在国内外猪群中的感染现状已相当严重,成为一个不容忽视的问题[1]。通常,猪感染BVDV 不表现出临床症状,或表现出类似于猪瘟症状,因此猪BVDV 感染和CSFV 感染常被混淆[2]。这不仅给猪瘟检疫带来了困难,也给BVDV 防控添加了新的难题。BVDV 感染动物可以引起免疫抑制,以此逃避宿主先天免疫功能,并在宿主机体中呈持续性感染[3]。研究证实,只有非致细胞病变(NCP)型BVDV 实验性感染动物才能诱发持续性感染[4]。目前,关于BVDV 逃避机体先天免疫的机制尚不清楚,而了解这一机制有助于制定更全面的防控措施,从而更好的预防和控制BVDV 在猪群和牛群中的感染。
泛素样蛋白ISG15 是干扰素α/β(IFN-α/β)信号的一个负调控因子,该蛋白可以抑制多种DNA 病毒和RNA 病毒的增殖,属于宿主的天然免疫蛋白[5]。因此,阐明ISG15 和猪源BVDV-2 之间的相互作用,不仅可以更清楚的了解ISG15 抗病毒机制,也可以为深入研究猪源BVDV-2 逃避细胞先天免疫作用的机制奠定基础。
1 材料和方法
1.1 病毒株、细胞、多抗及质粒 猪源BVDV-2 SH-28 株由本实验室分离鉴定;牛肾细胞系(MDBK)和质粒pEC129[6]由美国宾夕法尼亚大学Dr.Leonard教授赠予;鼠抗ISG15 多克隆抗体由本实验室制备;E.coli DH5α 由本实验室保存。
1.2 主要试剂 限制性内切酶、T4 DNA 连接酶、r Taq DNA 聚合酶、DL2000 Marker 均购自TaKaRa公司;Donor Equine Serum(HS)购自Hyclone(Thermo scientific);G418 Sulfate 购自Sigma-Aldrich 公司;TRIzol Reagent、Lipofectamine LTX and PlusTMReagent、SuperScript First-Strand Synthesis System for RT-PCR购自Invitrogen 公司;FastStart Universal SYBR Green Master(Rox)购自Roche 公司;poly I:C 购自Sigma公司。
1.3 引物设计 根据GenBank 中登录的牛IFN 刺激因子ISG15 基因序列(AF069133)设计一对引物,并在上下游引物末端分别引入NheⅠ和Bam HⅠ酶切位点。引物由上海基康生物技术有限公司合成(表1)。
内参基因GAPDH、IFN-α、IFN-β 和ISG15 基因荧光定量PCR 引物由上海Invitrogen 公司设计并合成(表1)。
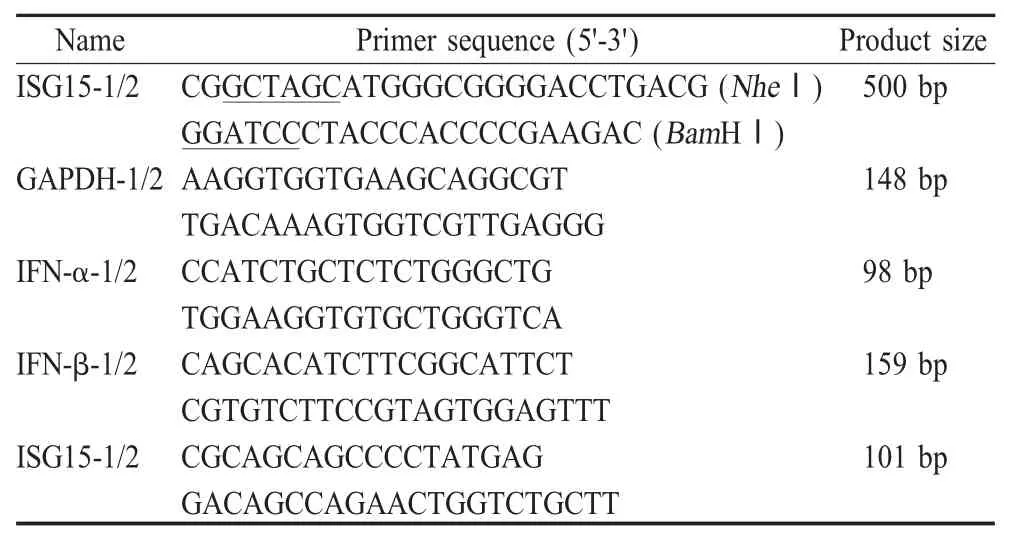
表1 各基因的PCR 引物Table 1 PCR primers for amplification of the genes
1.4 ISG15 基因的扩增及重组真核表达质粒的构建以人源IFN-α-2A 刺激MDBK 细胞后[7],提取细胞总RNA 并反转录制备cDNA,采用引物ISG15-1/2 PCR 扩增ISG15 基因。以NheⅠ和Bam HⅠ双酶切后克隆于真核表达载体pEC129 中,构建重组真核表达质粒pEC-ISG15,并进行测序鉴定。
1.5 稳定表达ISG15 蛋白细胞系的构建与筛选将G418 设定10 个浓度梯度,以100 μg/mL 为递增值,分别为100 μg/mL~1 000 μg/mL,每组设3 个平行样。连续培养10 d~14 d,待MDBK 细胞全部死亡时G418 的最低浓度为MDBK 细胞最佳筛选浓度[8]。筛选稳定细胞系时使用的浓度比最佳筛选浓度高一个稀释度,维持液浓度为最佳筛选浓度的一半。
参照Lipofectamine LTX and PlusTMReagent 说明书,将pEC129-ISG15 转染MDBK 细胞;培养6 h后,将细胞上清液换成含有10 % HS 的DMEM 完全培养液。同时设置空质粒对照。参照文献[9]方法,将转染24 h 后的MDBK 细胞传代至新的6 孔细胞板中(约2×104个/孔)。待细胞贴壁后,将培养液更换为含最佳工作浓度G418 的完全培养液进行抗性筛选;每2 d~3 d 更换一次培养基,直至阴性对照组中细胞全部死亡;将转染质粒细胞孔内筛选培养基更换为维持培养基,即调整G418 终浓度为300 μg/mL,继续培养细胞,直至6 孔板中长出单克隆细胞团;将其进行胰酶消化传代,以含300 μg/mL G418 培养液中继续培养,并逐渐扩大培养,形成稳定的传代细胞系,将其命名为E53。
1.6 稳定表达ISG15 蛋白细胞系的鉴定
1.6.1 间接免疫荧光(IFA)鉴定 将第10 代建立的细胞系E53 铺于24 孔培养板,以鼠抗ISG15 多克隆血清(1∶2 500)为一抗,FITC 标记的羊抗鼠IgG(1∶5 000)为二抗进行ISG15 蛋白表达的IFA 鉴定[10]。
1.6.2 Western blot 鉴定 将第10 代建立的细胞系E53 细胞裂解后,进行收集细胞总蛋白,进行SDS-PAGE 电泳,转膜后以鼠抗ISG15 多克隆抗体(1∶2 500)为一抗,HRP 标记的羊抗鼠IgG(1∶5 000)为二抗,进行ISG15 蛋白表达的western blot 鉴定。
1.7 病毒增长曲线的绘制 将病毒(1 MOI)分别感染单层MDBK 细胞和E53 细胞。在感染后不同时间点分别收集细胞上清液,并分别测定其TCID50,绘制病毒在两种细胞中的病毒滴度增长曲线。
1.8 I 型IFN 的荧光定量PCR 检测 弃去细胞上清,加入1 mL 细胞裂解液,4 ℃作用30 min,收集单层细胞,12 000 r/min 离心10 min;吸取400 μL上清液,参照PrimeScriptTMRT reagent Kit with gDNA Eraser 说明书,提取细胞总RNA 并反转录。将cDNA 稀释3 倍后作为模板,按FastStart Universal SYBR Green Master(Rox)说明书,采用SYBR Green I 法进行荧光定量PCR 扩增,以GAPDH 基因为内参,检测各样品中I 型IFN 的相对表达量。同时设阴性对照,检测数据利用2-△△CT方法进行处理[11]。采用SPASS13.0 软件进行差异显著性分析。
1.9 ISG15 对病毒诱导I 型IFN 表达影响的检测取等量的E53 细胞和MDBK 细胞分别铺于6 孔细胞板中,24 h 后各孔内加入终浓度为50 μg/mL 的poly I:C;作用6 h 后,各孔加入1 MOI BVDV-2;吸附1 h 后,弃去细胞上清,分别加入DMEM 维持液继续培养;24 h 后,收集细胞,利用荧光定量PCR 检测细胞内IFN-α 和IFN-β 的表达情况。
2 结果
2.1 ISG15 基因的扩增及重组真核表达质粒的鉴定利用ISG15-1/2 引物PCR 扩增ISG15 基因,结果显示,扩增片段约为500 bp,与预期相符(图1)。将PCR 扩增产物经双酶切后克隆至真核表达载体pEC129 中,经鉴定,重组质粒pEC-ISG15 构建正确。
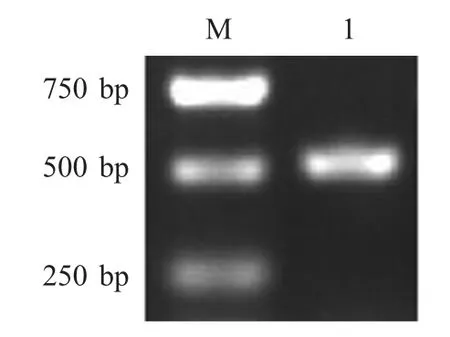
图1 ISG15 的PCR 扩增Fig.1 Amplification of ISG15 gene by PCR
2.2 稳定表达ISG15 蛋白细胞系的鉴定 经筛选,MDBK 细胞的最佳筛选浓度为600 μg/mL G418,获得单克隆细胞团后,即利用300 μg/mL G418 的维持液进行扩大培养。
以ISG15 多克隆抗体为一抗,对第10 代稳定细胞系E53 进行IFA 和western blot 检测,同时设MDBK 细胞作为阴性对照。结果显示,稳定细胞系E53能够稳定表达ISG15 蛋白(图2)。
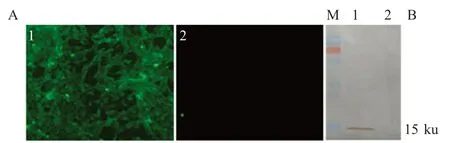
图2 过表达ISG15 细胞系E53 的IFA(A)和western blot(B)的鉴定Fig.2 Idetification of the ISG15 over-expressed cell line E53 by IFA(A)and western blot(B)
2.3 ISG15 蛋白对BVDV-2 复制的影响 将BVDV-2以1 MOI 的剂量分别感染E53 和MDBK 细胞,于不同时间点分别测定其病毒滴度,并绘制病毒增长曲线。结果表明,病毒在MDBK 细胞中滴度呈逐渐递增趋势,60 h 时病毒滴度达到峰值(TCID50为10-6.5);在E53 细胞中,病毒滴度虽有所增长,但变化趋势不大,60 h 时TCID50为10-3.8(图3)。两者差异显著(p<0.05),表明过表达ISG15 蛋白对BVDV-2的复制具有明显抑制作用。
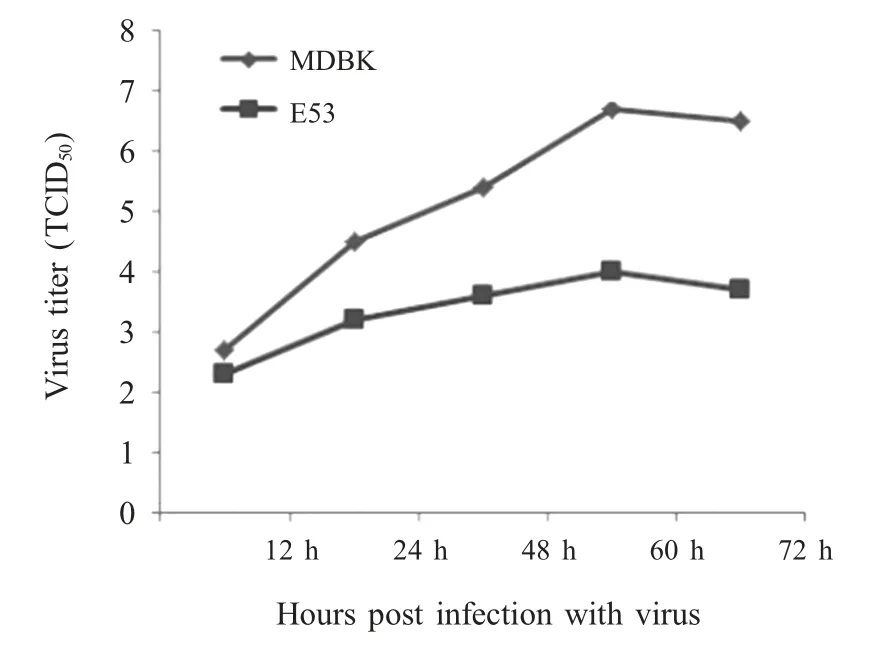
图3 ISG15 蛋白对猪源BVDV-2 复制的影响Fig.3 The effect of ISG15 protein on the replication of BVDV-2
2.4 BVDV-2 感染对ISG15 基因表达的影响 将BVDV-2 以1 MOI 的剂量感染E53 细胞同时设未感染的E53 细胞空白对照;分别在不同时间点收集细胞并提取其总RNA,利用荧光定量PCR 检测各时间点细胞内ISG15 基因表达情况;结果显示,与空白对照相比,病毒感染的E53 细胞中ISG15 基因表达量明显高于空白对照E53 细胞,48 h 时ISG15 基因的表达差异显著(p<0.05),表明BVDV-2 感染细胞后能够促进ISG15 的表达(图4)。

图4 猪源BVDV-2 感染细胞对ISG15 基因表达的影响Fig.4 The effect of pig BVDV-2 infection on the relative mRNA transcription levels of ISG15 gene
2.5 ISG15 基因对BVDV-2 诱导I 型IFN 表达的影响 病毒感染MDBK 和E53 细胞,结果表明均能够抑制由poly I:C 诱导产生的IFN-α 和IFN-β。MDBK细胞中,IFN-α 和IFN-β 表达量分别下降约2.49 和3.31 倍;而E53 细胞中,IFN-α 和IFN-β 分别下降约6.29 倍和9.71 倍,表明ISG15 蛋白的存在能够提高BVDV-2 抑制I 型IFN 表达的能力(图5)。

图5 ISG15 促进BVDV-2 抑制I 型IFN 表达Fig.5 The role of ISG15 on BVDV-2 inhibited IFN-I
3 讨论
研究报道,ISG15 在宿主天然免疫反应中发挥重要作用,它能够抑制多种病毒的复制,从而发挥抗病毒功能[12]。因此,研究ISG15 蛋白和猪源BVDV-2 之间相互作用关系,不仅有助于了解ISG15 的抗病毒机制,也为猪源BVDV-2 逃避细胞先天免疫机制的研究奠定基础,从而为BVDV 持续性感染机制的研究提供依据,有助于制定更全面的BVDV 防控措施及BVDV 新型疫苗的研制。
SH-28 株是由本实验室分离鉴定的一株NCP 型猪源BVDV-2 分离株,以此为基点,结合以前所做的基础工作,本实验展开猪源BVDV-2 和ISG15 相互作用研究。结果显示BVDV-2 SH-28 株在E53 细胞系中增殖速度明显下降,表明过量表达ISG15 能够抑制猪源BVDV-2 复制。此外,感染BVDV-2 可以促进细胞内ISG15 的表达,这可能是由于BVDV-2感染细胞,激活了细胞的先天免疫,使ISG15 表达量升高,从而发挥其抗病毒功能。此外,我们还研究了ISG15 蛋白对猪源BVDV-2 抑制I 型IFN 表达的影响,将SH-28 株分别感染经poly I:C 处理的MDBK 细胞和E53 细胞,利用荧光定量PCR 检测不同细胞内I 型IFN 的表达情况,结果表明经SH-28株感染后,E53 细胞内I 型IFN 表达下降的倍数高于MDBK 细胞内I 型IFN 下降的倍数,表明ISG15能够促进猪源BVDV-2 抑制I 型IFN 的表达,为探索BVDV-2 的免疫抑制机制提供了线索。
[1]戴益民,张文波,刘文峰,等.江西部分地区猪瘟疑似病例中牛病毒性腹泻病毒感染情况的初步调查[J].中国兽医杂志,2010,46(7):12-14.
[2]王新平,周绪斌.猪感染牛病毒性腹泻病毒的研究进展[J].动物医学进展,1998,19(1):1-4.
[3]骆延波,张绍学,柴家前,等.牛病毒性腹泻病毒免疫机理研究进展[J].山东农业大学学报:自然科学版,2000,31(2):214-216.
[4]Peterhans E,Schweizer M.BVDV:a pestivirus inducing tolerance of the innate immune response[J].Biologicals,2013,41(1):39-51.
[5]Sadler A J,Williams B R.Interferon-inducible antiviral effectors[J].Nat Rev Immunol,2008,8(7):559-568.
[6]Wang Lin-shu,Sunyer J O,Bello L J.Fusion to C3d enhances the immunogenicity of the E2 glycoprotein of type 2 bovine viral diarrhea virus[J].J Virol,2004,78(4):1616-1622.
[7]蔡新斌.ISG15 的原核表达和多克隆抗体的制备及其与猪瘟病毒相互作用的初步研究[D].南宁:广西大学,2011.
[8]林美娟,薛志平,陈平华,等.抗生素G418 在甘蔗组织培养中的最佳筛选浓度[J].福建农林大学学报:自然科学版,2006,35(1):6-10.
[9]龙鼎新,李小玲,温元武.稳定表达绿色荧光蛋白的COS7细胞系的筛选和鉴定[J].中南医学科学杂志,2012,40(1):11-13.
[10]任敏,焦海宏,林燕清,等.基因2 型牛病毒性腹泻病毒新疆及山东分离株的鉴定[J].中国动物传染病学报,2009,(4):20-25.
[11]韩猛立,黄新,钟发刚.牛病毒性腹泻病毒感染牛外周血单核细胞对IFN-α,β,γ mRNA 转录的影响[J].中国 兽医学报,2012,32(008):1110-1115.
[12]刘畅乔,王琛,耿运琪.类泛素蛋白ISG15 及其在先天免疫中的作用[J].生物化学与生物物理进展,2006,33(11):1023-1029.
猜你喜欢
环球时报(2022-09-20)2022-09-20 15:18:57
今日农业(2022年14期)2022-09-15 01:43:28
猪业科学(2021年3期)2021-05-21 02:05:54
今日农业(2020年24期)2020-12-15 16:16:00
湖南畜牧兽医(2016年3期)2016-06-05 08:37:56
兽医导刊(2016年12期)2016-05-17 03:51:50
山东医药(2015年14期)2016-01-12 00:39:43
江苏大学学报(医学版)(2015年2期)2015-04-17 06:49:51
中国医药导报(2015年26期)2015-02-28 22:07:44
现代检验医学杂志(2015年4期)2015-02-06 02:02:06
