
倒木干扰对红松针阔混交林空间结构的影响1)
2015-03-08 05:45葛晓雯王立海侯捷建
东北林业大学学报 2015年7期
葛晓雯 王立海 侯捷建
(东北林业大学,哈尔滨,150040)
责任编辑:王广建。
倒木是树木在外力作用(通常是风、雨、雪等气候干扰)下,绝大部分树干主体倒落在地面上,即倒木。倒木一般是已经自然死亡的枯木,也有树冠巨大的壮年树木。主要包括干折、根倒和连根拔起3 种存在形式。森林空间结构是林分中树木及其属性在空间的分布;具体来说,就是林木空间分布格局、树种混交和树木竞争[1],森林空间结构是森林生长过程中的驱动因子,对森林未来的发展起决定性作用[2]。分析林分空间结构需要测定树木的位置坐标及其与相邻木的关系,这是区别于非空间结构的主要标志[2]。
目前,国内外关于森林生态系统中倒木的研究内容主要有:倒木对林分更新的影响[3-4]、倒木自身的生物贮量及分解研究[5-9]、倒木呼吸[10-12]、倒木对微立地环境的影响[13-16]、倒木对生物多样性的维持[17-19]以及风倒木力学机理的研究[20-23]等。而针对林分空间结构的研究主要集中在空间结构的分析比较和模拟。Aguirre 等[24]以50 m×50 m 的固定样地研究了墨西哥天然林的混交度。Pommerening A[25]利用多个空间结构指数对4 个大小不等的样地的空间结构进行了比较分析。由于树木生长周期长,短期内很难得出普适性结论,所以,国外不少学者尝试对森林空间结构及其动态发展进行模拟研究,如:将同龄林转化为异龄林[26]、纯林转化为混交林[27]以及对不同采伐方式的影响进行模拟[28]等。国内,安惠君[29]对阔叶红松林空间结构进行较为系统的研究,并首次提出“林层比”这一表达复层林林层结构的参数。惠刚盈等[30-31]在大量试验的基础上,提出角尺度、大小比数和混交度3 个空间结构参数,并对结构参数在森林空间结构分析中的应用和优选方法进行了探讨总结。汤孟平[32]率先提出以森林空间结构为目标的结构优化经营思想,并建立理论模型。吴蒙蒙等[33]探究了不同采伐强度(人为干扰)对阔叶红松林空间结构的影响。韩卫东等[34]针对南方强台风(极端自然天气干扰)前后风景区内林分空间结构的变化进行了研究。然而,针对倒木这种非极端自然干扰因子对森林空间结构的影响未见报道。
采用角尺度、混交度和大小比数3 个空间结构参数并结合树种组成,量化分析倒木产生前后对红松针阔混交林的空间结构的影响,为揭示自然干扰对林分空间结构的影响提供科学的参考依据。
1 试验地概况
试验地设在黑龙江省伊春市带岭区凉水国家级自然保护区,位于我国小兴安岭山脉的东南段——达里带岭支脉的东面,地理坐标为128°47'8″E~128°57'19″E,47°6'49″N~47°16'10″N。该区地形以低山丘陵地貌为主,坡度为10°~15°,平均海拔400 m。夏季温凉多雨,冬季严寒干燥,具有明显的温带大陆性季风气候特征,年平均气温0.5 ℃,年平均降水量676 mm,积雪期130~150 d,年平均蒸发量806 mm,无霜期100~120 d。该区春夏多西南风,秋冬多西北风。
保护区内森林类型多样,物种丰富,以红松为主的针阔混交林是原生地带性顶级群落。主要树种有:冷杉(Abies fabri)、椴树(Tilia tuan)、青楷槭(Acer tegmentosum)、红松(Pinus koraiensis)、花楷槭(Acer ukurunduense)、毛赤杨(Alnus sibirica)、白桦(Betula platyphylla)、水曲柳(Fraxinus mandshurica)、云杉(Picea asperata)、榆树(Ulmus pumila)、暴马丁香(Syringa reticulate)等。
2 研究方法
2.1 样地设置与调查
2010年9月,利用全站仪在该保护区内建立固定监测样地约30 hm2,将样地划分为20 m×20 m 的连续样方,并在每个样方的四个顶点处埋设水泥桩标记。随机选取其中一块200 m×140 m(2.8 hm2)的样地,平均海拔350 m,坡度为5°~7°。分2 次进行调查,第1 次于2010年10月进行,全面调查样方内所有乔木的树种、相对坐标、胸径、树高及生长状况等因子,并挂编号牌;第2 次调查于2014年5月进行,清点挂牌的倒木,记录其树种、胸径、树高以及地理坐标等基本因子。经查阅气象资料,得知样地区域近4 a 并未发生会导致林木大面积倒伏或折断的极端天气状况。因此,所调查的倒木即为建立样地以来在正常自然干扰下产生的。
2.2 数据分析方法
林分的空间结构分析是以空间结构单元为基础进行的。林分中每株树(目标树)以及它的n 株最近相邻木,构成了林分内最基本的空间结构单元。选用n=4,即目标树i 与4 株最近相邻木组成的结构单元(见图1),该种结构单元具有中间过渡类型,较符合现实情况,生物学意义明显[35]。
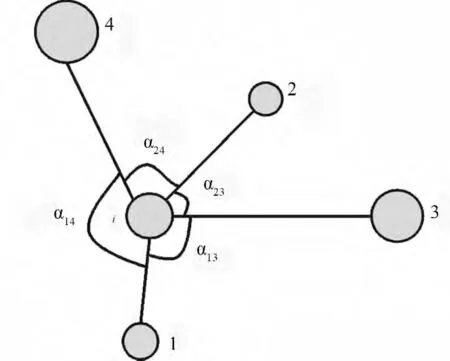
图1 空间结构单元示意图
角尺度(Wi)是α 角小于标准角(α0=72°)的个数占所考察的4 个α 角的比例。目标树i 与任意两株最近相邻木的夹角中的小角为α[35]。表达式为:当第j 个α 角小于标准角α0时,zij=1,否则zij=0。Wi为0、0.25、0.5、0.75、1.00 时,分别表示4株最近相邻木在目标树周围的分布情况为特别均匀、均匀、随机、不均匀和聚集,它反映的是林分中林木个体的分布格局。均值ˉW 可以反映一个林分的整体分布情况。
混交度(Mi)是4 株最近相邻木中与目标树i 不属于同一树种的个体比例[35]。表达式为vij,当参照树i 与j 株相邻木非同种时,vij=1,否则vij=0。Mi为0、0.25、0.5、0.75、1.00 时,分别表示第i 株目标树与周围4 株最近相邻木组成的结构单元是零度、弱度、中度、强度和极强度混交。
大小比数(Ui)是4 株最近相邻木中大于目标树的个体所占的比例[35]。表达式为当相邻木j 比参照树i 小时,kij=0,否则kij=1。Ui为0、0.25、0.5、0.75、1.00 时,分别表示4 株相邻木中比目标树小的有4 株、3 株、2 株、1 株和0 株,相应地目标树i 在该结构单元中所处的生态位分别为优势、亚优势、中庸、劣态和绝对劣态。
本文利用空间结构分析软件Winkelmass 对倒木产生前后的样地调查数据进行处理,利用Excel2010 和SPSS16.0 对原始数据和处理结果进行统计相关分析及非参数检验。
3 结果与分析
3.1 样地林分树种组成
倒木产生前,该样地内共有22 个乔木树种(见表1),顶级树种红松的相对多度为8.31%,相对显著度却高达42.68%,在林分中处于绝对优势地位;冷杉和椴树无论从相对多度还是相对显著度来讲都占有一定的优势;青楷槭的相对显著度很小,主要以中、小径个体居多;林分中稠李、鱼鳞云杉、杨树、鼠李、臭冷杉、山杨和榛树的相对多度在1%以下,相对显著度也很小;从各树种的平均胸径来看,杨树和红松的平均胸径在40 cm 以上,分别为47.7 cm 和42.9 cm,鱼鳞云杉、山杨和臭冷杉的平均胸径分别达到了34.2、33.0 和27.1 cm,但这3 个树种的相对多度和相对显著度并不高。
相对多度=(某个树种的株数/全部树种的株数)×100%;相对显著度=(某个树种的断面积/全部树种的断面积)×100%;某个树种的断面积,i 为某个树种的株数。

表1 倒木产生前样地树种组成的数量特征
经调查,样地内共有211 株倒木,其树种组成见表2,共有15 个树种出现倒木,以中小径木存在的青楷槭倒木最多,共42 株;其次,冷杉、毛赤杨和椴树倒木株数均超过20 株,分别为37、25 和24 株,而且这4 个树种在样地内的立木多度也较高;稠李、鱼鳞云杉、鼠李、臭冷杉、山杨和榛树等相对多度极低和根系较发达的水曲柳未出现倒木。经相关分析得到,各树种的倒木数量与其立木相对多度呈极显著正相关(r=0.816,P=0),如图2所示。倒木株数占立木总株数的7.15%,倒木现象在样地中很明显。倒木中最粗的是1 株胸径为100.1 cm 的杨树,其次是胸径为90.7 cm 的1 株红松,其余胸径均在40 cm以下,最细的1 株胸径仅为5.4 cm;倒木的平均胸径为16.7 cm,说明样地内倒木以中等胸径树木居多。从存在形式看,样地内连根拔起的倒木最多,其次是根倒,干折木最少。主要是由于试验样地坡度平缓、土壤湿度相对较大,降低了林木根系固持土壤的能力,使得林木在一定的外力作用下极易连根拔起;干折倒木最少则是由于样地地势低,所受风力相对较小;不同树种的倒木存在形式之间的差异则与每个树种本身的生长特性密切相关。

表2 样地内倒木树种组成的数量特征

图2 倒木株数与立木相对多度之间的关系
3.2 林分空间结构
角尺度表征林木的空间分布格局,有随机、均匀和团状3 种分布情况,当角尺度取值[0.475,0.517]时,属于随机分布,小于0.475 时为均匀分布,大于0.517 时则为团状分布[36]。混交度表示林分的树种隔离程度,值越大表明同种树相邻生存的状态就越少,林木对整个林分空间的利用越充分,森林生态系统整体越稳定。大小比数此处代表林木胸径指标的大小分化程度。
样地内倒木出现前后林分的空间结构变化(见表3)。由表3可知,角尺度、混交度和大小比数三个指标变化量都极小。虽然倒木产生前后,角尺度和大小比数减小、混交度增大,林分空间结构呈现出好的变化趋势,但样地内空间结构的整体状态并未发生改变,即林分整体分布仍属于团状分布,林分仍处于由中度混交向强度混交过渡的状态,样地内林木在胸径分化程度上仍都处于中庸状态。
为了明确倒木对林分整体空间结构的影响是否具有统计意义,本文分别利用Mann-Whitney U 检验和Kolmogorov-Smirnov Z 检验两种方法对倒木产生前后的3 个空间结构指标进行两组独立样本的非参数检验,结果显示各指标的概率值P 均大于显著性水平0.05,因此接受原假设,认为在倒木产生前后林分的角尺度、混交度和大小比数均没有显著性差异。也就是说在小兴安岭红松针阔混交林中,短期内自然干扰对林分整体空间结构影响不显著。

表3 倒木产生前后各空间结构指标变化情况
3.3 不同树种的空间结构
3.3.1 角尺度
由于角尺度体现的是林分内林木的空间分布格局,只与林木的坐标位置有关,按树种分析没有实际意义,因此仍对其作整体分析。如图3所示为样地内的林木分布情况,经计算,倒木的角尺度值为0.574 5,同样属于团状分布。因此,自然干扰下倒木的产生十分类似于农业生产中的“间苗”,虽然能够使林分整体更趋近随机分布,但短期内并不能改变团状分布的本质。
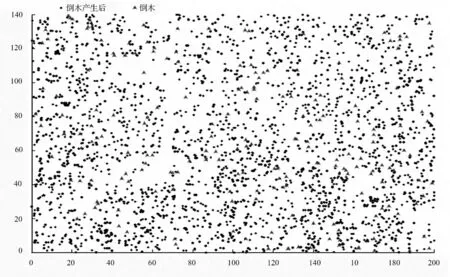
图3 样地内林木空间分布情况
3.3.2 混交度
为进一步了解样地内各树种的混交情况,对倒木出现前后各个树种的混交度进行比较分析(见表4)。在22 个树种中,鱼鳞云杉、杨树、鼠李、臭冷杉、山杨和榛树6 个树种的混交度前后无变化,其中榛树处于由强度混交向极强度混交过渡的状态,其余5 个树种均为极强度混交。主要是这6 个树种在样地内相对多度极低、生长位置较分散,而且多数树种未出现倒木(仅杨树有1 棵),所以其树种隔离程度一般不会发生改变。
由表4可知,倒木的产生使大部分树种的混交度都发生了不同程度的增减变化。冷杉、椴树、青楷槭、红松、毛赤杨、山桃和暴马丁香7 个树种的混交度增大,倒木比例较高(10%以上)的暴马丁香、毛赤杨、青楷槭和山桃的混交度增幅十分显著,另外3个树种的倒木比例较低(5%左右),其混交度增量不显著;枫桦、色木、花楷槭、白桦、水曲柳、云杉、稠李、红皮云杉和榆树9 个树种的混交度减小,未出现倒木的稠李混交度减幅最显著,倒木比例较低(6%以下)的色木、枫桦、云杉、水曲柳和红皮云杉的混交度减幅也比较显著,而倒木比例较高的花楷槭和白桦,其混交度减幅不显著。经相关分析,各树种混交度均值的增量(若混交度减小,则增量为负值)与该树种倒木比例呈极显著正相关(r=0.715,P=0.002)(见图4),这是因为某一树种倒木比例越高,该树种剩余林木个体与其他树种相邻生存的几率就越大,因此,混交度增量也越大。
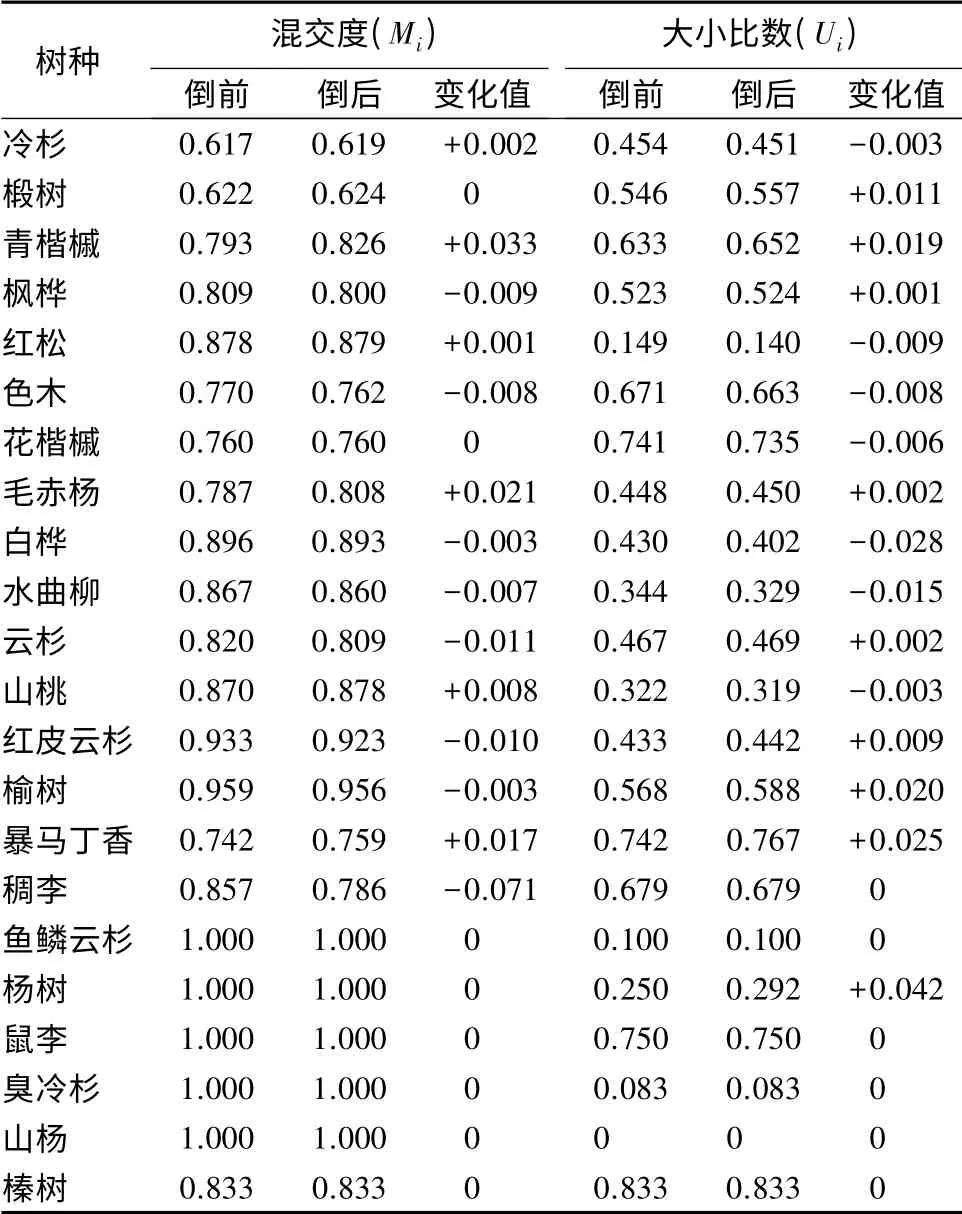
表4 倒木产生前后不同树种的混交度和大小比数变化情况
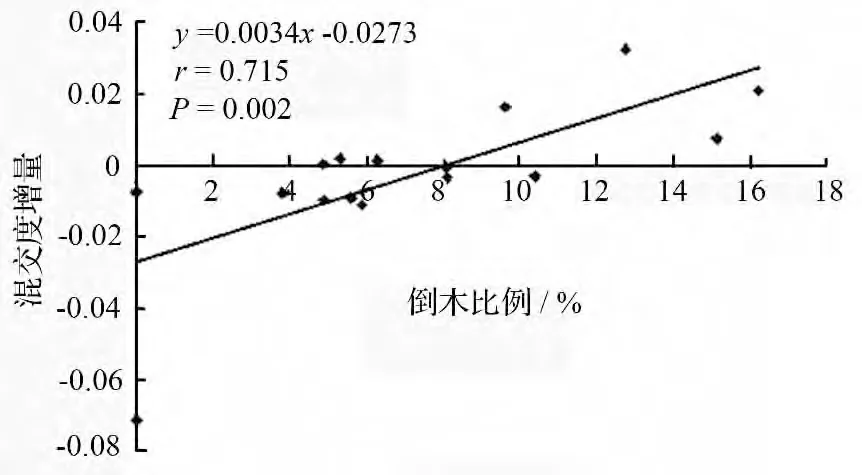
图4 不同树种混交度增量与倒木比例之间的关系
3.3.3 大小比数
对样地内不同树种的平均大小比数进行比较分析(见表4),稠李、鱼鳞云杉、鼠李、臭冷杉、山杨和榛树等6 个相对多度极低的未倒树种平均大小比数没有发生改变。平均胸径较大(分别为33、27.1 和34.2 cm)的前3 个树种在空间结构单元中均处于绝对优势地位,而平均胸径较小(分别为8.5、11.5 和6.5 cm)的后3 个树种均处于劣态地位。红松较其他树种的平均大小比数小,表明在空间结构单元中红松总体上较其他树种更具优势。倒木产生前后,不同树种的大小比数增减不一,冷杉、红松、色木、花楷槭、白桦、水曲柳和山桃等7 个树种的大小比数减小,椴树、青楷槭、枫桦、毛赤杨、云杉、红皮云杉、榆树、暴马丁香和杨树等9 个树种的大小比数增大;大小比数增大的树种组,倒木平均胸径明显大于大小比数减小的树种组。经相关分析得知,平均大小比数的增量与该树种倒木的平均胸径呈显著正相关(r=0.579,P=0.024)(见图5~6),即倒木平均胸径越大,该树种大小比数增量越大,在空间结构单元的生态位越恶劣;各树种平均大小比数的增减与该树种倒木和立木平均胸径之比呈极显著正相关(r=0.806,P=0)。一般地,比值大于1(即倒木平均胸径大于同树种的立木平均胸径)时,平均大小比数增大,比值小于1 时则减小。这是因为倒木平均胸径大于立木时,表明该树种有优势地位的个体减少,整体生态位恶化,故而平均大小比数增大。

图5 不同树种大小比数增量与倒木平均胸径之间的关系

图6 不同树种大小比数增量与倒木和立木平均胸径之比的关系
4 结论与讨论
利用角尺度、混交度和大小比数3 个空间结构指标并结合树种组成,对小兴安岭红松针阔混交林样地倒木产生前后的空间结构进行量化分析,得出以下结论:
红松针阔混交林样地中,倒木株数比例为7.15%,倒木现象在样地内很明显,而且各树种倒木数量与其立木相对多度呈极显著正相关(r=0.816,P=0),倒木以中等胸径木居多。从存在形式来看,掘根倒木最多、根倒次之、干折最少,这主要与地势、根系、土壤强度及树木自身生长特性有关。
倒木对林分整体空间结构的影响极小,虽然使角尺度和大小比数减小、混交度增加,是朝一个好的方向发展,但是在统计意义上差异并不显著。
倒木的角尺度为0.574 5,属于团状分布。可以将自然干扰下倒木的产生理解为农业生产中的“间苗”,虽能够使林分整体更趋近随机分布,但短期内并不能改变林分团状分布的本质。
倒木的产生使不同树种的平均混交度发生改变,混交度增量与该树种的倒木比例呈极显著正相关(r=0.715,P=0.002)。因为某一树种倒木比例越高,该树种剩余林木个体与其他树种相邻生存的几率就越大,故而混交度增量也越大。
倒木的产生使大多数树种的大小比数发生增减变化,而且树种平均大小比数的增量与其倒木的平均胸径呈显著正相关(r=0.579,P=0.024)。倒木平均胸径越大,该树种大小比数增量越大,在空间结构单元的生态位越恶劣;各树种平均大小比数的增减与该树种倒木和立木平均胸径之比呈极显著正相关(r=0.806,P=0)。一般地,比值大于1(即倒木平均胸径大于同树种的立木平均胸径)时,平均大小比数增大,比值小于1 时则减小。倒木平均胸径大于立木时,表明该树种有优势地位的个体减少,整体生态位恶化,故而平均大小比数增大,反之亦然。
总之,在小兴安岭红松针阔混交林,倒木这种自然干扰因素对林分整体空间结构影响不显著,但对不同的树种有明显影响。
[1] 汤孟平,陈永刚,徐文兵,等.森林空间结构分析[M].北京:科学出版社,2013.
[2] 汤孟平.森林空间结构研究现状与发展趋势[J].林业科学,2010,46(1):117-122.
[3] 班勇,徐化成,李湛东.兴安落叶松老龄林落叶松林木死亡格局以及倒木对更新的影响[J].应用生态学报,1997,8(5):449-454.
[4] 王俊峰,郑小贤,王铁牛,等.长白山云冷杉林倒木贮量及倒木对更新的影响[J].华北农学报,2004,19(S1):45-49.
[5] 杨丽韫,代力民,张扬建.长白山北坡暗针叶林倒木贮量和分解的研究[J].应用生态学报,2002,13(9):1069-1071.
[6] 辛魏巍,铁牛,李卓凡,等.大兴安岭兴安落叶松原始林倒木养分含量研究[J].林业资源管理,2012(3):94-99.
[7] 康玲.内蒙古大兴安岭兴安落叶松与白桦倒木分解研究[D].呼和浩特:内蒙古农业大学,2012.
[8] Garrett l G,Oliver G R,Pearce M R,et al.Decomposition of Pinus radiata coarse woody debris in New Zealand[J].Forest Ecology and Management,2008,255(11):3839-3845.
[9] Yang F F,Li Y L,Zhou G Y,et al.Dynamics of coarse woody debris and decomposition rates in an old-growth forest in lower tropical China[J].Forest Ecology and Management,2010,259(8):1666-1672.
[10] 吴家兵,关德新,韩士杰,等.长白山地区红松和紫椴倒木呼吸研究[J].北京林业大学学报,2008,30(2):14-19.
[11] Jomura M,Kominami Y,Tamai K,et al.The carbon budget of coarse woody debris in a temperate broad-leaved secondary forest in Japan[J].Tellus,2007,59(2):211-222.
[12] Woodall C W.Carbon flux of down woody materials in forests of the North Central United States[J].International Journal of Forestry Research,2010,2010:1-10.
[13] 游惠明,何东进,蔡昌棠,等.天宝岩长苞铁杉林倒木对土壤肥力质量的影响评价[J].应用与环境生物学报,2013,9(1):168-174.
[14] 段文标,杜珊,陈立新,等.阔叶红松混交林林隙大小和掘根微立地对小气候的影响[J].应用生态学报,2013,24(8):2097-2105.
[15] Strahm B D,Harrison R B,Terry t a,et al.Soil solution nitrogen concentrations and leaching rates as influenced by organic matter retention on a highly productive Douglas-fir site[J].Forest Ecology and Management,2005,218(1/3):74-88.
[16] Kappes H,Topp W,Zach P,et al.Coarse woody debris,soil properties and snails (Mollusca:Gastropoda)in European primeval forests of different environment conditions[J].European Journal of Soil Biology,2006,42(3):139-146.
[17] 何东进,何小娟,洪伟,等.森林生态系统粗死木质残体的研究进展[J].林业科学研究,2009,22(5):715-721.
[18] Dechene A D,Buddle A M.Decomposing logs increase oribatid mite assemblage diversity in mixed wood boreal forest[J].Biodiversity and Conservation,2010,19(1):237-256.
[19] GateS G M,Mohammed C,Wardlaw T,et al.The ecology and diversity of wood-inhabiting macrofungi in a native Eucalyptus obliqua forest of southern Tasmania,Australia[J].Fungal Ecology,2011,4(1):56-67.
[20] 贾杰,李静辉.林木风倒动态模型的建立与分析[J].森林工程,2013,29(4):63-68.
[21] 宋晓鹤.云杉风倒静力学模型的建立及其分析[D].哈尔滨:哈尔滨工业大学,2006.
[22] 王琳.云杉风倒动力学模型的建立与分析[D].哈尔滨:哈尔滨工业大学,2006.
[23] 赖秋明.云杉风倒动力学问题的研究[D].哈尔滨:哈尔滨工业大学,2008.
[24] Aguirre O,Hui G Y,Gadow K V,et al.An analysis of spatial forest structure using neighbourhood-based variables[J].Forest Ecology and Management,2003,183(1/3):137-145.
[25] Pommerening A.Evaluating structural indices by reversing forest structural analysis[J].Forest Ecology and Management,2006,224(3):266-277.
[26] Hanewinkel M,Pretzsch H.Modelling the conversion from evenaged to uneven-aged stands of Norway spruce (Picea abies L.Karst )with a distance-depentant growth simulator[J].Forest Ecology and Management,2000,134(1):55-70.
[27] Kint V.Structural development in ageing temperate Scots pine stands[J].Forest Ecology and Management,2005,214(1):237-250.
[28] Courbaud B,Goreaudb F,Dreyfusc P,et al.Evaluating thinning strategies using a tree distance dependent growth model:some examples based on the CAPSIS software“uneven-aged spruce forests”module[J].Forest Ecology and Management,2001,145(1):15-28.
[29] 安惠君.阔叶红松林空间结构研究[D].北京:北京林业大学,2003.
[30] 惠刚盈,胡艳波.混交林树种空间隔离程度表达方式的研究[J].林业科学研究,2001,14(1):23-27.
[31] 惠刚盈,Gadow K,胡艳波.林分空间结构参数角尺度的标准角选择[J].林业科学研究,2004,17(6):687-692.
[32] 汤孟平,唐守正,雷相东,等.林分择伐空间结构优化模型研究[J].林业科学,2004,40(5):25-31.
[33] 吴蒙蒙,王立海,侯红亚,等.采伐强度对阔叶红松林空间结构的影响[J].东北林业大学学报,2013,41(9):6-9.
[34] 韩维栋,黄剑坚,刘素青.强台风干扰后林分空间结构变化的研究[J].中南林业科技大学学报,2013,33(7):8-13.
[35] 惠刚盈,赵中华,胡艳波.结构化森林经营技术指南[M].北京:中国林业出版社,2010.
[36] 惠刚盈,胡艳波.角尺度在林分空间结构调整中的应用[J].林业资源管理,2006(2):31-35.
猜你喜欢
浙江农林大学学报(2022年4期)2022-08-18
卫星应用(2022年3期)2022-05-23
新农民(2021年21期)2021-09-17
阿来研究(2020年1期)2020-10-28
绿色科技(2017年15期)2017-09-01
新世纪水泥导报(2016年1期)2016-07-01
中央社会主义学院学报(2016年2期)2016-05-04
土木建筑工程信息技术(2013年1期)2013-10-17
