
木质藤本植物黑老虎的居群结构与动态1)
2015-03-07 07:15:26付玉嫔司马永康祁荣频郝佳波毛云玲尹艾萍陈少瑜刘冬芬张荣贵
东北林业大学学报 2015年9期
付玉嫔 司马永康 祁荣频 郝佳波 毛云玲 尹艾萍 陈少瑜 刘冬芬 张荣贵
(国家林业局云南珍稀濒特森林植物保护和繁育重点实验室(云南省林业科学院),昆明,650201) (红河州林业科学研究所)
责任编辑:任 俐。
居群结构和动态变化是自然植物居群的重要统计特征,包括居群的年龄结构、分布格局、垂直结构、性比、居群数量及动态变化过程等[1]6,[2]。居群的年龄结构对分析过去居群的结构和受干扰状态、预测未来的居群动态具有重要作用,是分析居群动态的基础[2]。而植物居群分布格局的研究对确定居群特征、居群个体间相互关系及居群与环境间的关系具有非常重要的作用[3]。开展居群分布格局研究的目的不仅在于对植物居群水平结构进行定量描述,更重要的是能够揭示格局的成因,阐述居群的动态变化,并为该植物居群的生存现状、是否需要开展保护、开发利用等的探讨奠定科学基础[4]。目前,关于乔木类珍稀、濒危树种的居群结构及动态的研究较多[5-9],也提出了很多相应的研究方法[3]。但是关于木质藤本植物的研究主要集中在物种组成、多度、多样性、分布等方面[10-12],具体物种的居群结构及动态研究未见相关报道。
黑老虎(Kadsura coccinea (Lemaire)A.C.Smith)为五味子科(Schisandraceae)南五味子属(Kadsura Jussieu)木质藤本植物,具有多种用途。根、茎为传统民药,具有消肿、解毒、治疗跌打损伤、中枢抑制、安神、强壮等功效,在保肝、治疗慢性肝炎、抗氧化、抗HIV 病毒、抗衰老等方面也有作用[13]。此外,其果大、味酸甜,富含多种人体必需的氨基酸[14]和微量元素,无毒并具有调节血脂的作用[15],是一种具有潜在开发价值的珍稀野生水果[16-17]。在我国的四川、浙江、广东、广西、贵州、海南、湖南、江西和云南等地以及缅甸、老挝、泰国、越南和印度尼西亚等都有分布,生长于海拔200~2 000 m 半开阔的灌丛和林中。在云南主要产于屏边、金平、河口、景东、普洱、景洪、勐海、文山等县、市。近年来由于大量的取果、取藤、挖根等现象的普遍存在,加之生境的严重破坏,该物种的存活和繁殖已面临极大的挑战。目前,有关黑老虎分类学[18-19]、形态学[20-21]、药理活性[22]、药物化学[23-27]等方面的研究报道很多,但关于该物种的居群结构及动态研究尚无相关报道。文中以云南3 个天然群落保护较好的黑老虎居群为对象,旨在通过对黑老虎不同居群的空间分布格局和动态的研究,初步揭示其格局的成因和阐述居群的动态变化,并评估这一木质藤本植物居群的生存前景和保护价值。
1 研究区概况
1.1 分水岭自然概况
研究地点位于云南省分水岭国家级自然保护区内的分水岭林区。该自然保护区地处哀牢山山脉东南端,东经102°31'36″~103°31'50″,北纬22°26'36″~22°57'44″,为低纬度、高海拔区域,海拔1 300 m 区域年平均气温17.6 ℃,降水量2 448 mm,相对湿度83%,随海拔上升,气温下降、降水量增加,每上升100 m,年平均气温下降0.7 ℃,降水量增加100 mm,相对湿度增加1%[28]。黑老虎主要分布于保护区海拔1 700~2 500 m 的山地苔藓常绿阔叶林中。常见伴生植物有:基脉润楠(Machilus decursinervis.)、水青树(Tetracentron sinense)、木果石栎(Lithocarpus xylocarpus)、山地山龙眼(Helicia silvicola)、尖叶桂樱(Laurocerasus undulata)、五裂槭(Acer oliverianum)、小果冬青(Ⅰlex micrococca)、红椎(Castanopsis hystrix)、黧蒴栲(Castanopsis fissa)、异形南五味子(Kadsura heterroclita)、买麻藤(Gnetum montanum)等。在样地调查中,黑老虎常见于东北和东南坡向,凹形坡或直线坡的中上部,坡度较大,为25~35°,西南坡向少见。主要生长于山地黄壤或山地黄棕壤,母质为辉长岩、砂岩、青石等,表层腐殖质层明显,以下土体呈黄棕色、块状结构、质地黏重。样地内人为活动方式为放牧和采果,干扰强度较强。
1.2 大围山自然概况
研究地点设在云南省大围山国家级自然保护区。该保护区位于东经103°20'~104°03',北纬22°35'~23°07',雨量丰沛,年降水量达1 700~1 900 mm,每年1—5月份云南其他地区遭遇春旱时,仍然拥有420 mm 以上的降水量。年均温22.6 ℃,最冷月均温(1月份)15.2 ℃,最热月均温(7月份)27.7℃,大于10 ℃的积温8 246.2 ℃,偶有降雪和霜冻发生,区内海拔76.4~2 365 m[29]。黑老虎在该区域主要生长在海拔1 200~1 800 m 的季风常绿阔叶林中,常见伴生植物有木果石栎(Lithocarpus xylocarpus)、厚鳞石栎(Lithocarpus pachylepis.)、鹿角栲(Castanopsis lamontii.)、滇越猴欢喜(Sloanea mollis)、滇南杜鹃(Rhododendron hancockii)、青冈(Cyclobalanopsis sp.)、山地山龙眼(Helicia silvicola)、小花红花荷(Rhodoleia parvipetala)、平伐含笑(Michelia cavaleriei)、西南粗叶木(Lasianthus henryi)、半齿铁仔(Myrsine semiserrata)、黄丹木姜子(Litsea elongata)、扁担藤(Tetrastigma planicaule)等。黑老虎生长于东南坡、东坡及东北坡向,直线坡或凹形坡的中部、下部,坡度25°~45°,土壤为黄壤,母质为泥质页岩或青石。人为活动影响方式为频繁采果、取藤和放牧,干扰强度较强。
1.3 无量山自然概况
研究地点位于云南省无量山国家级自然保护区。该自然保护区地处滇西缅北、横断山脉、云贵高原、中南半岛4 个地理区域的结合部位,东经100°19'~100°45',北纬24°17'~24°55'。黑老虎主要分布在海拔1 800~2 200 m,陡峭、湿润的季风常绿阔叶林内,该地段属中亚热带至北亚热带气候,有降雪、霜等。黑老虎常生长于北坡、西北坡、西坡至南坡坡向,东南坡、东坡和东北坡向少见,生长于直线坡或凸坡的下部直至谷底或沟边,棕壤上,所生长的林分,群落高可达25~30 m,林内空气湿度大,黑老虎常攀援至群落的最上层,至树冠顶端,叶和花常生长在相当高之树冠层,并与其他植物混合生长而不易辨识。人为活动影响方式以放牧、取藤、采果为主,干扰强度较轻。常见半生植物有楠叶枫(Acer pinnatinervium)、八角枫(Alangium chinense)、截头石栎(Lithocarpus truncatus.)、山地山龙眼(Helicia silvicola)、梗花粗叶木(Lasianthus biermannii)、黄丹木姜子(Litsea elongata)、大果冬青(Ⅰlex macrocarpa)、光皮桦(Betula luminifera)、野核桃(Juglans cathayensis)、红花木莲(Manglietia insignis)、云南拟单性木兰(Parakmeria yunnanensis)、买麻藤(Gnetum montanum)、大花木荷(Schima forrestii)、云南黄杞(Engelhardtia spicata)等。
2 研究方法
2.1 样方设置和调查方法
采用典型抽样法分别在云南分水岭国家级自然保护区、云南大围山国家级自然保护区和云南无量山自然保护区等3 个黑老虎居群内设置10 m×10 m的样方共计53 个,样地总面积达5 300 m2,其中分水岭设置11 个、大围山设置25 个、无量山设置17个。分别记录每个样方的海拔、坡向、坡度、坡位和土壤类型等生境因子以及人类活动影响方式和强度。在样方内,调查并记录其中的种类、株数、攀援高度、基径等指标。
2.2 数据统计与分析方法
相对年龄结构分级:用立木的大小级代替相对年龄结构的方法常用于具有明显主干树木的年龄结构分析中。文中的研究对象黑老虎为木质藤本植物,具有明显的主干,并与树木一样具有形成层和年轮,所以出于保护的目的,也采用以立木级代替相对年龄结构[9]的方法分析其年龄结构。文中参考我国学者曲仲湘[30]提出的5 级立木分级标准,并结合黑老虎始花相对年龄大小的具体情况,将黑老虎的立木分级标准划分为高度<33 cm 为I 级;高度≥33 cm,基径<0.5 cm 为II 级、0.5 cm≤基径<1.5 cm 为III 级、1.5 cm≤基径<4.5 cm 为IV 级、4.5 cm≤基径为V 级,由于藤本植物高生长极为旺盛,基径与胸径的差异很小,故文中以藤体的基径代替胸径。依据此分级标准将调查样地内所研究居群的个体进行分级统计,并绘制出居群立木级分布图。
高度结构分级:黑老虎为藤本攀援植物,在其生长过程中需要借助乔木树种作为支柱支持其生长和攀援。藤本植物的攀援特性决定了其垂直空间的结构与支柱木的高度密切相关,其藤体高度与支柱木的高度具有一致性,当藤攀援太高时测量藤体高度可操作性差,所以黑老虎幼苗和幼藤以自身的藤体高为分级高度,其他的以支柱木的高度代替黑老虎的高度进行高度结构分级,从而了解其垂直空间结构和动态特征。依据文献和实际调查结果,文中以5 m 为一个高度级统计各高度级内的株数(上限排外法)以确定高度结构,并绘制出居群高度级分布图[9-10]。
空间分布格局分析:文中选用方差均值比率法作居群水平空间分布格局的判定方法[9-10]。即在泊松分布中方差(S2)与均值(M)之比等于或接近于1,为随机分布;如果S2/M 的值大于1,则为集群分布;S2/M 的值小于1,则为均匀分布。同时,采用t检验来确定实测值与预期值的偏离程度[9-10]。
特定时间生命表编制:以立木的相对年龄大小级来代替绝对年龄级,根据特定时间生命表的编制原理和方法[31],计算生命表中各参数,编制黑老虎居群的特定时间生命表。其中:qx=dx/lx,Lx=(lx+lx+1)/2,Tx=∑Lx,ex=Tx/lx,式中:x 为单位时间年龄等级的值;ax为在x 龄级内出现的个体数;dx为从x到x+1 龄级间隔期内标准化死亡数;ex为进入x 龄级个体的生命期望寿命;lx为在x 龄级开始时标准化存活个体数;Lx为从x 到x+1 龄级间隔期内还存活的个体数;qx为从x 到x+1 龄级间隔期内的死亡率;Tx为从x 龄级到超过x 龄级的个体总数。
存活曲线绘制:依据特定时间生命表,以立木的相对年龄大小级为横坐标,以In(lx)为纵坐标,绘制黑老虎的存活曲线。
3 结果与分析
3.1 不同居群的相对年龄结构
根据调查统计结果,黑老虎在3 个居群样地内分别出现的植株数量为分水岭居群(A)38 株,大围山居群(B)61 株,无量山居群(C)49 株,各居群的居群数量都很少。从年龄结构(表1)可以看出,分水岭居群和大围山居群可分为4 个年龄级:Ⅱ、Ⅲ、Ⅳ和Ⅴ级;无量山居群仅可分为3 个年龄级:Ⅲ、Ⅳ和Ⅴ级,3 个居群均无Ⅰ级(幼苗级),而且都以成年藤到中、老年藤占据居群数量的绝大部分。其中,大围山居群为倒金字塔形,分水岭和无量山居群则接近倒金字塔形。3 个居群不同的年龄级所占比率不同,都以Ⅳ年龄级比率最大,由大到小的顺序为无量山(55.1%)、分水岭(52.6%)、大围山(42.6%),而且Ⅳ和Ⅴ级占据的比例都超过了50%,大围山居群Ⅳ和Ⅴ级占到了81.9%,无量山居群Ⅳ和Ⅴ径级占到73.5%,分水岭居群Ⅳ和Ⅴ径级占到60.5%;幼苗均无储备,幼藤的储备由大到小的顺序为分水岭居群(39.4%)、无量山(26.5%)、大围山(18.0%),3 个居群都属于衰退型居群结构,其中以大围山居群和无量山居群的衰退较严重,分水岭相对弱一些。
3.2 不同居群的高度结构
从表2可以看出,黑老虎3 个居群的高度结构不同,无量山居群的高度结构完整,可分为6 个高度级,大围山居群有5 个高度级,而分水岭居群仅分布在前3 个高度级。3 个居群中,分水岭居群的黑老虎植株数量随着高度级的增加而急剧递减,递减到15 m≤高度(H)<20 m 高度级则为0 株,当前幼年个体储备达71.05%,有利于群落中下一层向上一层的补充。大围山居群的黑老虎植株数量从0≤H<5 m 高度级到15 m≤H<20 m 高度级呈现递减趋势,而到20 m≤H<25 m 高度级又急剧增加,其中从10 m≤H<15 m 高度级向15 m≤H<20 m 高度级递减最快,甚至接近0 株,该居群在15 m≤H<20 m 高度级的储备非常低,不利于该层个体向上一层的补充,但其幼年个体储备丰富,达59.01%,整体上是有利于该群落个体下层向上一层的补充。无量山居群黑老虎植株数量从0≤H<5 m 高度级到5 m≤H<10 m 高度级稍有增加,然后从5 m≤H<10 m 高度级到15 m≤H<20 m 高度级为依次递减至最少,后又从15 m≤H<20 m 高度级依次增加至25 m≤H<30 m 高度级,与大围山居群一样在15 m≤H<20 m 高度级的株数储备最低,不利于该层向上一层的补充。
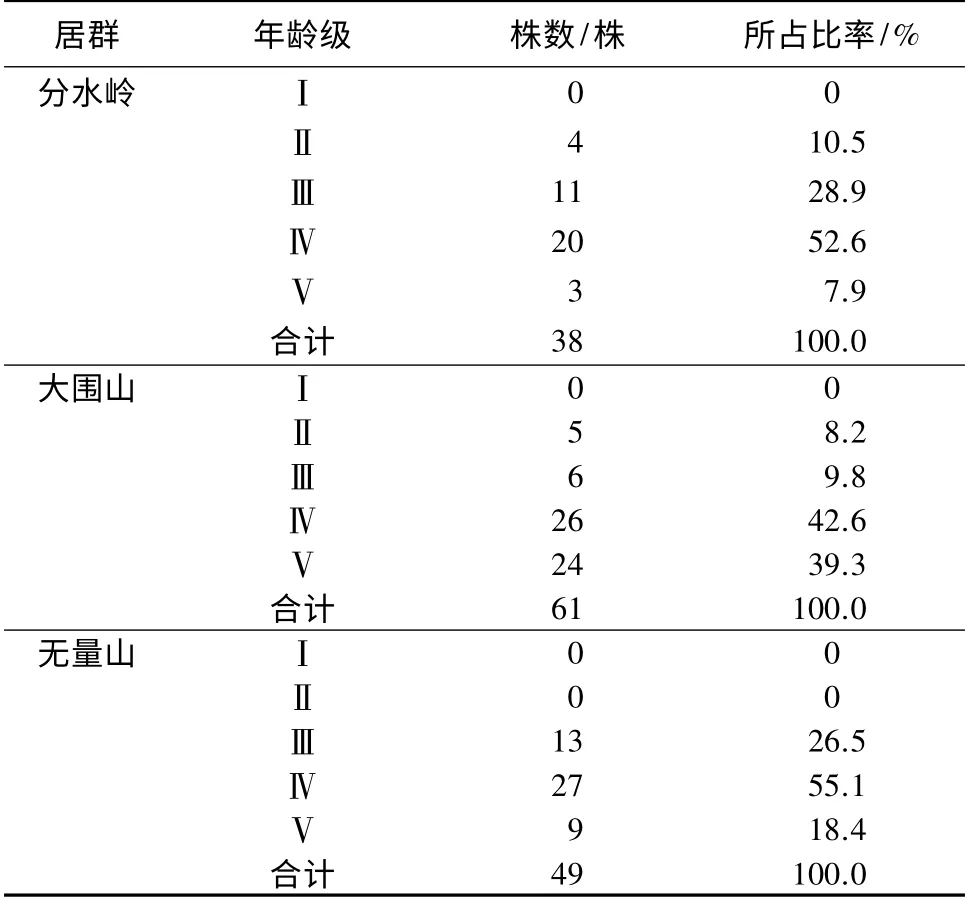
表1 黑老虎不同居群的年龄结构
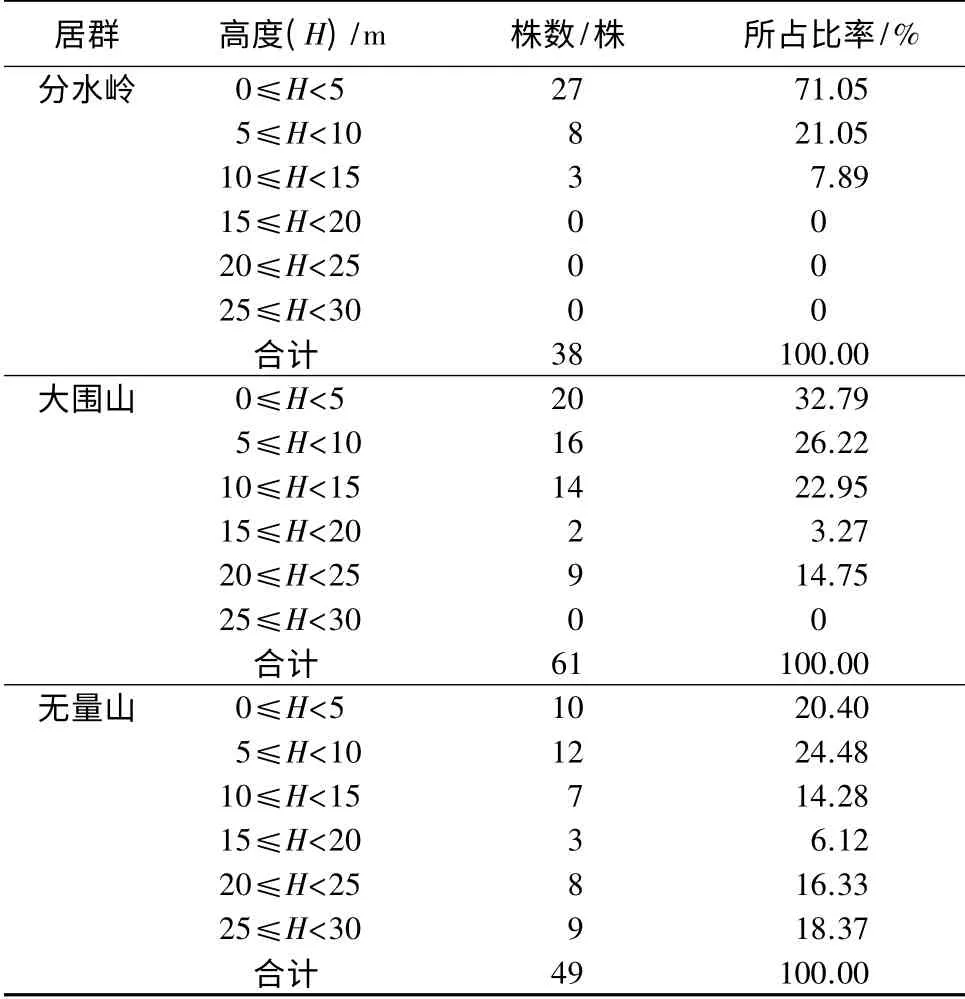
表2 黑老虎不同居群的高度结构
3.3 空间分布格局和动态
由表3可以看出,3 个居群的分布格局表现并不同,大围山和无量山居群都表现为随机分布,且在P=0.05 水平上检验不显著;而分水岭居群为集群分布,且在P=0.05 水平上检验显著,这可能与分水岭的幼藤级所占比例相对较大,而且多与萌生起源有关。进一步对3 个居群的Ⅱ、Ⅲ、Ⅳ和Ⅴ各年龄级的分布格局进行判定,其中分水岭居群Ⅲ、Ⅳ年龄级的S2/M 值大于1,为集群分布,但在P=0.05 水平上t检验不显著,Ⅱ和Ⅴ年龄级的S2/M 值接近于1,为随机分布,且在P=0.05 水平上t 检验不显著,所以,分水岭居群各年龄级均表现为随机分布;大围山居群和无量山居群的各年龄级的S2/M 值均接近或略大于1,且在P=0.05 水平上t 检验不显著,所以都表现为随机分布。表明3 个居群的各年龄级的分布格局的变化不大,仅在幼藤到中年藤阶段略有集群的现象,但到中年藤后期,受密度或环境因素的影响集群程度降低,表现为随机分布的格局。
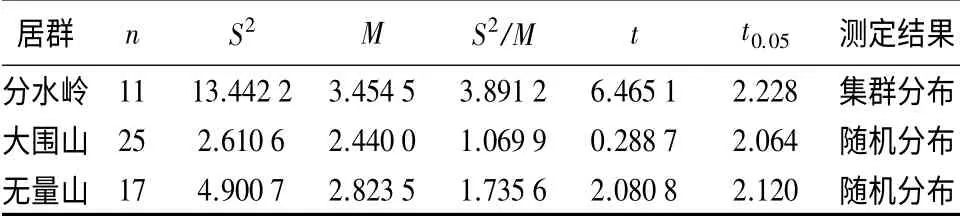
表3 黑老虎3 个居群的空间分布格局
3.4 特定时间生命表
由表4可以看出,3 个居群均无幼苗级,所以黑老虎3 个居群从幼苗长成Ⅴ级老藤的死亡(qx)趋势大致可以分为3 个阶段。
幼藤阶段(II~III 级):这一阶段3 个居群的死亡率都最低,其中分水岭居群为-1.750,大围山居群为-2.611,无量山居群为-1.077。但3 个居群在幼藤期的变动又有不同,分水岭从幼藤的前期到后期死亡率上升,大围山居群则幼藤前期阶段死亡率较高,生长稳定后死亡率下降。无量山幼藤期死亡率最低。黑老虎经过苗期的适应过程,到了幼藤期对环境的适应能力增强,此外,幼藤期各个个体占据的空间小,个体间的竞争能力小,加之处于林分冠层下受霜冻、雪灾天气的影响较小,能耐一定的庇荫,因此,黑老虎在这一时期保持了最低的死亡率。
成年至中年藤阶段(III 级后期~IV 级前期、中期阶段):这一阶段3 个居群的死亡率都大幅上升,达到了较高的比例,其中,分水岭居群上升到了0.850,大围山居群上升到了0.787,无量山居群上升到了0.840。黑老虎幼藤进入成年期以后,为了获取充足的光照,已经攀爬至林冠层上方,并尽可能的占据林冠层上方的水平空间,一株黑老虎可以同时占据周围空间隔离较小的4~5 株优势木的树冠,至中年时期各个个体形成了各据一方的局面,所以个体之间的竞争,尤其是冠层空间的竞争达到了最大,而且这一时期也是乔木树种竞争最强的时期,所以自梳和它梳的作用决定了其较高的死亡率。
老藤阶段(IV 级后期~Ⅴ级):这一阶段3 个居群的死亡率都迅速升到了最高,达到了1.000,黑老虎植株生理功能出现了明显的衰退,已很难应对分布地突然遭遇的干旱、霜冻、雪灾等特殊气候,此外,可能还受到攀援木死亡、倒塌等影响,都形成了其生命历程中的第一次死亡高峰。
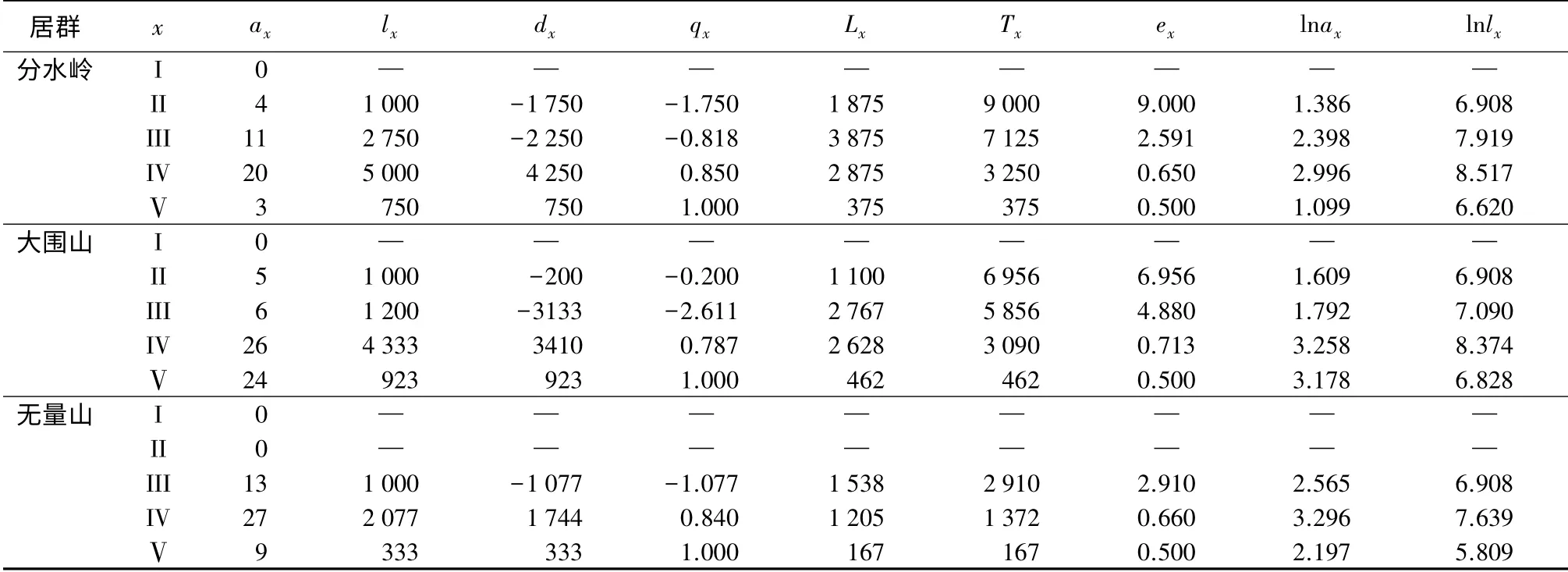
表4 黑老虎3 个居群的特定时间生命表
从死亡动态可以看出,3 个居群黑老虎的生理年龄大约处于IV 级阶段。均属于衰退居群。
平均生命期望(ex)反映的是x 龄级内个体的平均生存能力[7]。从表2可以看出,3 个居群黑老虎的ex值都以II 级幼藤阶段最高,并由大到小的顺序表现为分水岭居群、大围山居群、无量山居群。说明幼藤期分水岭居群的黑老虎平均生存能力最强,其次是大围山居群,无量山居群相对最弱。3 个居群的ex值从II 级幼藤到Ⅲ级幼藤,Ⅲ级幼藤到Ⅵ级中年藤都表现为陡降,从Ⅵ中年藤到Ⅴ级老藤表现为缓慢下降。表明黑老虎3 个居群幼藤期的个体生存能力并不稳定,受到的影响较大,具体造成幼藤期生存能力剧烈变动的原因尚需深入研究。黑老虎从幼年期向成年期、中年期转变时其ex值又陡降,分水岭居群陡降75%,大围山居群陡降85.3%,无量山居群陡降77.3%,这可能是从幼年期向成年、中年期转变时对生存空间要求的急剧增大,导致个体间剧烈竞争的结果。中年期向老年阶段过渡期,虽然生存空间需求也在逐渐增大,但导致的竞争减弱,所以其生存能力受到的影响较小,生存能力下降主要是自身生理功能缓慢衰退导致的。
3.5 存活曲线
根据表4编制的特定时间生命表的结果,绘制出黑老虎3 个居群的存活曲线如图1。
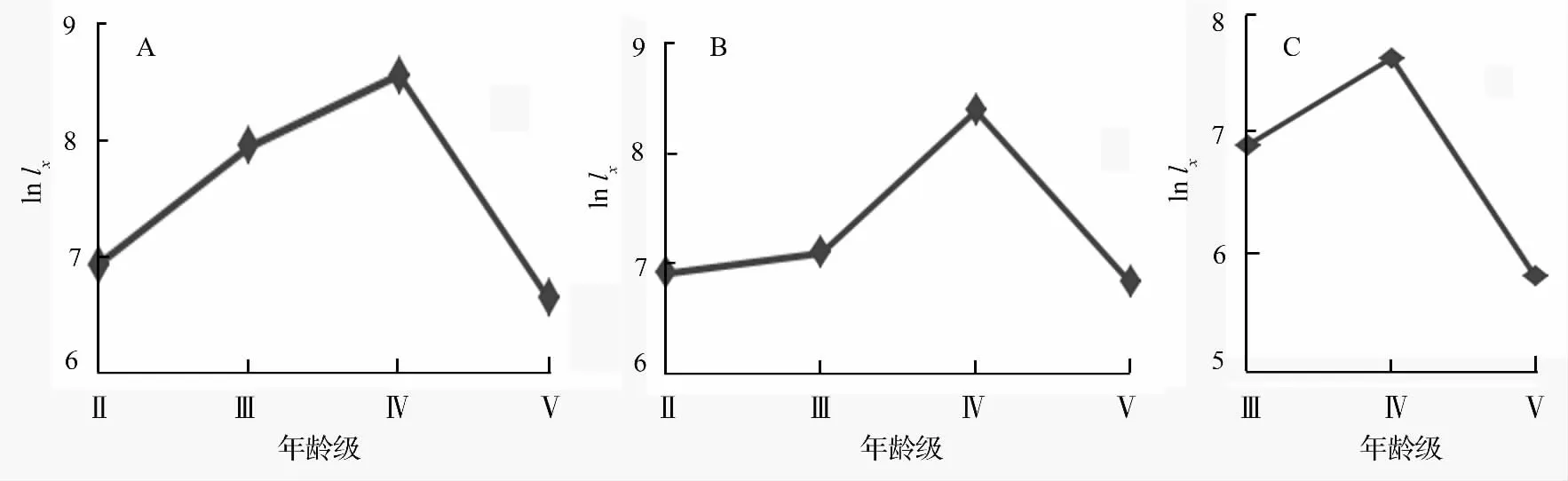
图1 黑老虎3 个居群的存活曲线
将黑老虎3 个居群的存活曲线与Deevey 存活曲线进行比较,3 个居群的存活曲线均介于Deevey I和II 型曲线之间,接近于Deevey I 型,呈明显的凸型,根据Leak[32]的划分标准,黑老虎3 个居群的年龄结构均属于衰退型,各居群正处于衰退状态。
4 结论和讨论
到目前为止,就藤本植物的居群特征方面的研究尚属空白。如何对藤本植物的居群特征及动态变化有一个清楚认识,对一些重要藤本药材的开发利用和保护具有重要的意义。居群的年龄结构是居群的重要特征之一,是居群内不同年龄个体数量的分布情况,它反映了居群当时的发育阶段及状况,同时也预示了居群的数量变化和未来的动态趋势[1]8。从文中研究结果可以看出,所调查的3 个黑老虎居群相对年龄结构都不完整,均缺少Ⅰ级幼苗,以成年、中年年龄级的比例最高,由大到小依次表现为无量山居群(55.1%)、分水岭(52.6%)、大围山(42.6%),仅中老年藤的比例就超过了50%,大围山居群占到81.9%,无量山居群占到73.5%,分水岭居群占到60.5%,各个居群的幼藤储备都非常低,均属于衰退型居群。就实地调查样地的情况而言,黑老虎为资源植物,根和藤茎都为中草药药材,其次果实为当地百姓喜欢采集食用的野生水果,目前该植物人工培育较少,主要以野生采集为主,所以大量而频繁的取藤、摘果、挖根、放牧等是目前3 个居群样地内人为活动的主要方式,取藤和挖根直接导致黑老虎结实产量下降,摘果直接造成了种子库种子的缺失,频繁的践踏则成为少量留存种子萌发更新的又一重要破坏活动,因此,人为活动是造成3 个居群幼苗缺失,幼藤储备少而为衰退居群的重要原因。植物居群自我更新能力从某一方面直接反映其居群生存和发展的潜力,自我更新能力的丧失预示着居群的衰老和消亡[9]。上述原因是黑老虎居群幼苗缺乏的一个方面。另一方面,木质藤本植物为喜光植物,其更新演替过程和生长主要受限于支柱木和光照获取能力,而光照的获取能力又与林分年龄和破坏程度有紧密的联系[33]。文中调查的3 个黑老虎居群均位于保护区范围内,一般到达下层林木的光照少和幼苗构建的微环境缺乏是这3 个居群所处林分的主要特征[33],如果黑老虎的种子传播不到林窗或林冠开放的地段,到达地表的光照较少,其更新、成长将变得困难。此外,由于林冠层对阳光的遮蔽以及由此而加剧的居群内和居群间个体的竞争(包括自感或者他感作用),使得幼苗或者幼藤的延续生长受到显著影响亦会导致缺失现象的发生[4]。再者,居群年龄结构的差异和波动,是居群发育过程中生物学特性,种内种间竞争以及环境因子综合作用的结果[34]。所以人为活动和林分自身的演替动态导致光照和支柱木获取能力降低都对黑老虎居群的更新和繁衍有影响,亦综合导致了较少的幼苗数量或幼苗缺失,使其处于衰退状态。因此,不仅要对黑老虎居群进行保护,及时促进人工更新也很重要。
此外,在实地调查中,3 个居群样地内出现的幼藤部分为萌生起源和克隆繁殖,而萌生起源往往是因为植物受到外界人为干扰和破坏造成其适应环境的另外一种繁殖方式,所以黑老虎萌生幼藤较多,可能是以前人们大量的取藤、摘果以及野生动物的取食导致的,后来建立了保护区以后人为破坏和干扰减少了,萌生繁殖的幼苗就少了。黑老虎种子的传播机制和幼苗构建过程尚不清楚,导致其更新极度困难,幼苗构建无法完成的原因和机理需要进一步研究。
居群的高度结构即居群个体的高度分布情况,反映了居群个体在垂直空间的结构状况和动态特征,也是居群的重要特征之一。从文中研究结果可以看出,黑老虎3 个居群的高度结构不同,分水岭居群仅有3 个高度级,大围山居群有5 个高度级,无量山居群有6 个高度级。依据Putz[35]对藤本植物的划分方法,黑老虎属于茎缠绕攀援藤本植物,其攀援高度直接受支柱木高度和胸径的影响,大径级支柱木被藤本植物攀援的比率高于小径级支柱木,茎缠绕攀援藤本的胸径与支柱木胸径极显著相关(P<0.001)[36]。所以3 个居群黑老虎的高度结构的大小级数的不一样不仅与林冠层的高度、支柱木的高度和胸径有直接关系,亦与能否获取到光照和支柱木有紧密关系[33]。为了获取充足的光照,黑老虎在攀爬至林冠层前以攀爬高度为主,当黑老虎攀援到林冠层后则迅速占领冠层的表平面,变为以增加攀援面积为主。分水岭的黑老虎居群由于所处林分多为次生林,林冠层高度为10~15 m,支柱木胸径小,其次部分样地属于林分破坏后形成的林窗,林窗内更新生长的支柱木高度仅6~8 m,由此,形成了较少的高度结构分布级数。大围山和无量山居群的林分保持较好,冠层高度和支柱木高度为20~28 m,支柱木胸径较大,因此,具有相对完整的高度结构。3 个居群黑老虎的高度结构呈连续发育,当前各级都有黑老虎植株分布,并以幼藤期的储备数量最高,有利于群落中下一层对上一层的补充,但从年龄结构组成知道,其无Ⅰ级幼苗的状况预示着黑老虎各居群的未来发展是不利的,这种高度级的逐层补充可能仅能持续一定时间后会发生停钝,如再无更新幼苗向上一层高度级的补充,那黑老虎居群的存在和繁衍将变得十分严峻。
居群个体在水平空间的分布方式或者在水平空间上居群个体之间的相互关系称为分布格局[37]。居群的分布格局则由居群特性、居群关系和环境条件的综合影响所决定,是居群对环境长期适应和选择的结果[1]25。黑老虎3 个居群的分布格局不同,显然与它自身的生物学特性、生态习性、营养繁殖特性和所处的环境有关。具有高克隆繁殖特性的木质藤本植物,克隆繁殖的藤茎在空间分布上有更高的集群分布,这种集群分布亦是对林冠受干扰后或林窗的补充[38]。文中研究结果显示,分水岭的黑老虎居群呈集群分布,其所处林分为次生林分,之前受人为干扰较大,形成的林窗较多,加之对黑老虎挖根、取藤等现象时有发生,其次是偶尔发生的低温现象致使地上部分死亡,受干扰后其根茎部位的隐芽启动萌发长成新的藤茎,幼藤多为萌生起源,所以形成集群分布的格局主要与其萌生起源和克隆繁殖有关。黑老虎和许多木质藤本植物一样具有进行远距离克隆繁殖的特点[39],藤茎从林冠垂落到地面产生根系或根系沿林下地面不断伸长再产生根系的方式克隆出新的藤茎。这从另外一个层面也反映出,黑老虎克隆繁殖的特性可能是其种子传播和更新困难条件下适应环境的一种生殖策略,尚需研究证明。通常情况下,随着居群的不断发育,居群内个体对环境条件的要求加强,由密度制约因素引起的种内和种间的竞争加剧[40],增加了居群个体的死亡率,导致居群个体的集群强度随着年龄的增加而迅速下降,直至集群解体成为随机分布[41]。而且居群在幼年阶段较高的集群强度有利于存活和发挥群体效应,而成年时由于个体增大,自疏作用增强[42],集群强度降低则利于获得足够的环境资源[41]。在大围山和无量山2 个黑老虎居群中,中、老年龄级的个体分别占到了81.9%和73.5%,个体明显增大,自梳作用和支柱木的它梳作用都明显增强,其次,这2 个居群所处林分林龄较大,林分受干扰相对较少,以远距离克隆繁殖为主,克隆繁殖的藤茎少,所以这2 个居群的分布格局表现为随机分布。
对3 个居群各年龄级的分布格局的判定结果表明,仅分水岭居群Ⅲ、Ⅳ年龄级有集群现象,但t 检验结果表明差异不显著,所以都表现为随机分布的格局,各时期分布格局的动态变化不明显。
从居群的特定时间生命表可知,3 个居群黑老虎的成长包括幼藤、中成年藤和老藤3 个阶段,存活曲线都表现为明显的凸形,年龄结构均属于衰退型,都正处于衰退状态。
从上述分析讨论可知,黑老虎这一植物居群目前更新的方式主要为克隆繁殖,种子自我更新能力极弱,居群数量少,正处于衰退状态,且目前还在不断地、大量的利用野生资源的现状都预示着该居群的生存和发展十分堪忧。应该积极倡导对黑老虎野生资源的保护、收集保存和尽快开展人工引种驯化。
致谢:特别感谢云南金平分水岭国家级自然保护区管理局喻智勇先生和梁宗利先生,云南哀牢山、无量山国家级自然保护区景东管理局谢有能先生、刘长铭先生和刘业勇先生,屏边苗族自治县林业科学研究所王绍昌先生和河口瑶族自治县林业局张天有先生和陈长丽女士在野外调查中的尽心帮助。
[1] 王伯荪,李鸣光,彭少麟.植物种群学[M].广州:广东高等教育出版社,1995.
[2] 陈小勇,张庆费,吴化前,等.黄山西坡青冈种群结构与分布格局研究[J].生态学报,1996,16(3):325-327.
[3] 兰国玉,雷瑞德.植物种群空间分布格局研究方法概述[J].西北林学院学报,2003,18(2):17-21.
[4] 刘建锋,肖文发,郭志华,等.极度濒危植物:崖柏种群结构与动态初步研究[J].江西农业大学学报,2004,26(3):377-380.
[5] 郝佳波,司马永康,陈少瑜,等.麻栗坡大果木莲的居群结构及动态研究[J].西部林业科学,2012,41(3):84-88.
[6] 刘建锋,江泽平,肖文发.极度濒危植物:崖柏种群空间格局与动态的初步研究[J].江西农业大学学报,2005,27(5):708-712.
[7] 王剑伟,张光富,陈会艳.特有珍稀植物宝华玉兰种群分布格局和群落特征[J].广西植物,2008,28(4):489-494.
[8] 杨汉远,冯邦贤,袁茂琴,等.珍稀濒危植物小叶红豆种群格局及濒危原因分析[J].种子,2013,32(6):52-54.
[9] 陈少瑜,付玉嫔,吴涛,等.濒危植物大果木莲种群格局及濒危原因分析[J].植物资源与环境学报,2012,21(2):102-106.
[10] 袁春明,刘文耀,杨国平.哀牢山湿性常绿阔叶林林窗木质藤本植物的物种组成与多样性[J].山地学报,2008,26(1):29-35.
[11] Balfour D A,Bond W J.Factors limiting climber distribution and abundance in a southern African forest[J].Journal of Ecology,1993,81:93-99.
[12] Padaki A,Parthasathy N.Abundance and distribution of lianas in tropical lowland evergreen forest of Agumbe,central Western Ghats,India[J].Tropical Ecology,2000,41(2):143-154.
[13] 艾菁,李于善.冷饭团化学成分及其活性研究进展[J].化学与生物工程,2005(2):7-9.
[14] 毛云玲,付玉嫔,祁荣频,等.云南黑老虎不同种源氨基酸和其他指标的分析与评价[J].氨基酸和生物资源,2015,37(2):14-19.
[15] 李志春,孙健,封毅,等.黑老虎果毒理实验及其对血脂的调节作用[J].食品科学,2011,32(1):203-205.
[16] 田定科.野生果树资源:冷饭团的利用初探[J].中国果业信息,2006,23(2):28.
[17] 邵则夏.五味子的食用药用价值与开发利用[J].中国野生植物资源,1995(4):33-35.
[18] 林祁,段林东,姚炳矾.南五味子属(五味子科)三种植物之补记[J].植物分类学报,2005,43(6):567-570.
[19] 毕海燕,林祁,刘长江,等.南五味子属(五味子科)的种子形态及其分类学意义[J].植物分类学报,2002,40(6):501-510.
[20] 段林东,林祁,袁琼.南五味子属花的形态及其系统学意义[J].植物研究,2004,24(1):87-92.
[21] 杨志荣,林祁,文香,等.英南五味子属(五味子科)植物叶表皮形态特征[J].植物研究,2009,29(2):147-163.
[22] 石焱芳,陈海玲.黑老虎的药理活性成份研究进展[J].海峡药学,2013,25(7):67-69.
[23] 王楠,李占林,华会明,等.黑老虎根化学成分[J].中草药,2010,41(2):195-197.
[24] Yan Song,Qing Jiezhao,Yong Shengjin,et al.A new triterpenoid from Kadsura coccinea[J].Chinese Chemical Letters,2010,21(11):1352-1354.
[25] 舒永志,成亮,杨培明,等.黑老虎的化学成分及药理作用研究进展[J].中草药,2011,42(4):805-813.
[26] 舒永志,成亮,曺濬喆,等.黑老虎的化学成分研究[J].中草药,2012,43(3):428-431.
[27] 王楠,李占林,刘晓秋,等.黑老虎根化学成分的研究(Ⅱ)[J].中国药物化学杂志,2012,22(4):305-309.
[28] 许建初.云南金平分水岭自然保护区综合科学考察报告集[M].昆明:云南科学技术出版社,2002:23-41.
[29] 王娟,杜凡,马钦彦,等.大围山国家级自然保护区药用植物资源及其多样性研究[J].北京林业大学学报,2002,24(4):6-11.
[30] 曲仲湘.南京灵谷寺森林现状的分析[J].植物学报,1952,1(1):l8-49.
[31] 李先琨,向悟生,唐润琴.濒危植物元宝山冷杉种群生命表分析[J].热带亚热带植物学报,2002,10(1):9-14.
[32] Leak W B.Age distribution in virgin red spruce and northen hardwoods[J].Ecology,1975,56:1451-1454.
[33] Dewalt S J,Schnitzer S A,Denslow J S.Density and diversity of lianas along a chronosequence in a central Panamanian lowland forest[J].Journal of Tropical Ecology,2000,16(1):1-19.
[34] Bonan G B.The size structure of theoretical plant population:spatial patterns and neighborhood effects[J].Ecology,1988,69:1721-1730.
[35] Putz F E.The natural history of lianas on Barro Colorado Island Panama[J].Ecology,1984,65:1713-1724.
[36] 袁春明,刘文耀,杨国平,等.哀牢山湿性常绿阔叶林木质藤本植物的物种多样性及其与支柱木的关系[J].林业科学,2010,46(1):15-22.
[37] 彭少麟.南亚热带森林群落动态学[M].北京:科学出版社,1996.
[38] Schnitzer S A,Mangan S A,Dalling J W,et al.Liana abundance,diversity,and distribution on Barro Colorado Island,Panama[J].PloS One,2012,7(12):52114.
[39] Yorke S R,Schnitzer S A,Mascaro J,et al.Increasing liana abundance and basal area in a tropical forest:the contribution of long-distance clonal colonization[J].Biotropica,2013,45(3):317-324.
[40] 刘智慧.四川省晋云山栲树种群结构和动态的初步研究[J].植物生态学与地植物学学报,1990,14(2):120-127.
[41] 金则新.浙江天台山甜槠种群结构与动态[J].生态学杂志,1999,18(6):10-15.
[42] Molles M C.Ecology:Concepts and applications[M].Beijing:China Science Press& McGraw-Hill Companies,Inc,2000:176-177.
猜你喜欢
中国果业信息(2023年9期)2023-12-17 07:43:49
河北科技师范学院学报(2022年2期)2022-08-26 08:55:24
浙江中医药大学学报(2021年6期)2021-07-12 03:05:14
兽医导刊(2019年1期)2019-02-21 01:13:44
民族音乐(2019年6期)2019-01-09 17:52:15
创作(2018年1期)2018-11-12 21:10:04
草地学报(2018年5期)2018-11-07 02:25:00
戏剧之家(2015年24期)2016-01-12 00:36:26
城市道桥与防洪(2014年1期)2014-02-27 07:24:36
河南水利与南水北调(2013年4期)2013-08-15 00:47:53
