
大肠埃希菌JM109表面抗原Ag43基因敲除与鉴定*
2015-03-04 07:55黄用豪赵焕阁周松森林映莹谭光宏黄风迎
重庆医学 2015年31期
黄用豪,赵焕阁,周松森,林映莹,谭光宏,黄风迎
(海南医学院海南省热带病重点实验室,海口571199)
表面抗原43(antigen 43,Ag43)是大肠埃希菌表达于细胞表面的重要抗原分子,该分子可以促使大肠埃希菌在特殊条件下形成生物膜,使细菌可以在极端的条件下生长并获得对多种抗生素的耐药性[1-3],是大肠埃希菌一个重要的功能分子。和细菌的菌毛和鞭毛不同,Ag43不需任何辅助分子,转运至细胞外膜的所有部件全部存在于该分子的组成结构中。Ag43分子由信号肽、α和β结构域3个功能部位组成[4-6],其中信号肽的结构能保证α和β结构域通过细胞的内质膜,β结构域则在细胞外膜上折叠成为一个桶状通道,α结构域穿过这一桶状通道后表达于细菌表面。Ag43的α与β结构域之间通过非共价键结合,加热到一定程度后α结构域就离开β结构域并溶解于水溶液中[7]。此外,由于α结构域含有大量的T 和B 淋巴细胞的抗原表位,因此Ag43还有可能是制备自身抗原分子疫苗,成为打破自身抗原分子免疫耐受的一个重要的抗原分子。
目前基因敲除是了解基因功能的重要手段,大肠埃希菌Ag43基因敲除后不但可以更进一步了解Ag43的功能,也有可能利用这一基因敲除菌作为一个Ag43嵌合蛋白的表达宿主细菌。因此,本研究利用基因工程技术将Ag43基因敲除并利用PCR 和基因测序技术对其进行鉴定。
1 材料与方法
1.1 材料 含有Ag43基因的大肠埃希菌JM109、细菌基因敲除试剂盒(TargeTron基因敲除系统,内含有pACD4K-C 质粒、连接酶、EBS通用引物等)购自Sigma公司。DNA Marker、PCR 试剂盒、凝胶回收纯化试剂盒、质粒提取试剂盒,内切酶NheⅠ和HindⅢ、电泳琼脂糖、异丙基硫代-B-D-半乳糖苷(IPTG)均购自大连宝生物公司。
1.2 方法
1.2.1 选择Ag43 基因敲除位点和突变RNA-蛋白复合体(RNP)的PCR 引物 从PubMed的基因库中查找到Ag43基因(CP000948.1)序列。登录Sigma公司提供的分析软件网站地址(www.sigma-aldrich.com/targetronaccess),将Ag43 基因序列的信息输入软件并按照提示进行操作,软件自动报告基因敲除(插入一个Group Ⅱintron)的最佳位置,并同时提供相应的3个PCR 引物(IBS、EBS1d、EBS2)的碱基序列信息。这3个PCR 引物决定了所敲除基因位点的特异性,由大连宝生物公司合成。
1.2.2 PCR 扩增突变Ⅱ组内含子RNP基因序列 将以上合成的3 个PCR 引物和试剂盒提供的EBS 通用引物混合成Four-primer mix,然后按照说明将试剂盒提供的Water、Intron PCR Template、Jump Start REDTaq Ready mix和Four-primer mix混合成50μL的PCR 反应体积。按94 ℃变性15s、55 ℃退火30s、72 ℃延伸30s进行30个循环反应,最后再72 ℃延伸2min。此后将PCR 产物在1% 琼脂糖上进行电泳分析,凝胶回收纯化试剂盒回收相应的PCR 产物。
1.2.3 Ⅱ组内含子RNP表达质粒pACD4K-C 的构建和鉴定 将以上回收的PCR 产物连同试剂盒提供的线性pACD4K-C质粒按T4连接酶要求的反应条件进行连接反应,获得本研究敲除Ag43基因的Ⅱ组内含子表达质粒pACD4K-Ag43。此后用上述同样的酶切方法对获得的重组质粒pACD4K-Ag43进行鉴定。
1.2.4 pACD4K-Ag43转化JM109菌株并诱导Ⅱ组内含子插入Ag43基因 按照TargeTron 基因敲除系统试剂盒说明书进行操作,将以上获得的pACD4K-Ag43重组质粒转化改造宿主细菌JM109,经冰浴和热休克后加入SOC培养基于37 ℃摇床培养1h,接着取100μL的转化培养物加入到3mL LB培养液中,37 ℃摇床培养过夜。第3天取40μL的过夜培养物加入到2mL 的LB培养液中,37 ℃摇床培养至OD600值为0.2后,加入IPTG(100mmol/L)诱导质粒表达,最后吸取100μL培养物至含有卡那霉素(25μg/mL)的LB培养基平板上30 ℃培养过夜,培养获得的菌株就可能是Ag43基因敲除的菌株。
1.2.5 Ag43基因敲除菌株基因测序鉴定 为了证明以上培养阳性菌株是否Ag43基因敲除菌,随机挑选以上培养卡那霉素阳性细菌并用大肠埃希菌基因组提取试剂盒提取相应的细菌基因组后,采用PCR 方法扩增相应区段基因序列进行琼脂糖凝胶电泳和基因测序鉴定。具体的PCR 扩增策略见图1所示。图示中For引物序列为:5′-CCA AAG CTT AAC GGT GA T ACC GG-3′,Rev引物序列为:5′-CCA CTC GAG TCA GAA GGT CAC AT-3′。A 方法采用For和Rev引物,PCR 产物约为4 500bp;B方法用For和EBS通用引物,PCR 产物为1 300~1 400bp;C方法用EBS2和Rev引物,PCR 产物约为3 500bp。最后将相应的PCR 产物交由大连宝生物工程有限公司进行测序。

图1 PCR 方法鉴定Ag43基因敲除细菌的引物位置及相应的产物
2 结 果
2.1 最佳Ⅱ组内含子插入位点及相应的RNP突变体PCR 引物 Sigma公司提供的分析软件分析结果发现,在Ag43基因上有多个位点适合插入Ⅱ组内含子,本研究选择E 值最高的1 812/1 913位点作为本研究的Ⅱ组内含子插入位点,同时选择这个位点的IBS、EBS1d和EBS2 3条特异引物供PCR 扩增特异RNP突变体,见图2。
2.2 多重PCR 扩增获得突变体RNP 的PCR 产物 按照Sigma公司的TargeTron基因敲除系统说明书要求,将以上3条PCR 引物连同试剂盒提供的EBS通用引物一起混成引物复合体,然后按照试剂盒推荐的PCR 反应条件进行多重PCR扩增,将扩增产物进行电泳分析。结果发现,PCR 扩增到的产物位于200~400bp之间,和预期350bp的相对分子质量大小相符,说明获得了Ag43特异RNP突变体的PCR 产物(图3,泳道1)。

图2 Ag43基因敲除位点和RNP突变体PCR 引物
2.3 表达RNP 的重组质粒pACD4K-Ag43的构建和鉴定 利用T4DNA 连接酶将RNP突变体的PCR 产物插入RNP表达质粒pACD4K-C 中,然后用酶切方法进行鉴定。经分析,Nhe I在RNP重组质粒上有两个酶切位点,酶切产物分别为3 646和4 029bp。Hind Ⅲ两个位点酶切产物为7 125和550 bp。图3第2泳道为Nhe Ⅰ酶切结果,两个片段大小和预测的3 646和4 029bp相一致;图3泳道3为HindⅢ酶切结果,切出约7 000和550bp两个片段。说明正确构建了含Ag43基因特异重组质粒,将这一重组质粒命名为pACD4K-Ag43。
2.4 Ag43基因敲除阳性菌株JM109(△Ag43)的鉴定 将重组质粒pACD4K-Ag43 转化含有Ag43 基因的大肠埃希菌JM109菌株,按照TargeTron基因敲除系统试剂盒所提供诱导和筛选方法,将Ⅱ组内含子基因序列插入Ag43 基因碱基1 812/1 813之间,造成Ag43基因功能丧失(基因敲除)。按照图1显示的A、B、C 3种鉴定方法对转化诱导细菌进行鉴定。3种PCR 扩增产物分别约为4 500、1 400和3 500bp,大小和预期相符(图3,泳道4~6)。另外,将这3 种PCR 产物的DNA 进行基因测序,测序结果和基因库中的Ag43 基因进行比对分析,发现Ag43基因的1 812/1 813位置插入了一段Ⅱ组内含子的DNA 序列。说明JM109菌株中的Ag43 基因被有效敲除,将这一株细菌命名为JM109(△Ag43)。
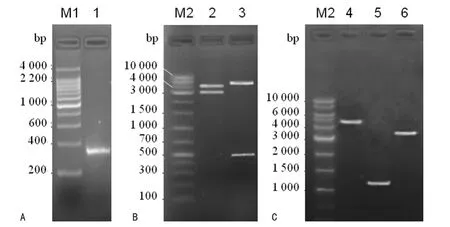
图3 重组质粒构建及鉴定
3 讨 论
本研究利用美国Sigma公司的专利细菌基因敲除试剂盒并按照试剂盒所提供的说明书的方法和步骤将一个带有卡那霉素耐药基因的Ⅱ组内含子插入大肠埃希菌JM109Ag43基因碱基1 812/1 913之间,造成Ag43基因功能失活,从而达到Ag43基因功能敲除的目。它是通过一个具有程序化限制性内切酶活性的RNP 将重组质粒pACD4K-C 表达的Ⅱ组内含子插入到Ag43基因中。由于本研究选用了1 812/1 913位点特异的3条PCR 引物(IBS、EBS1d和EBS2)和试剂盒EBS通用引物进行多重PCR 反应构建的pACD4K-Ag43重组质粒的特异性,使RNP能够定点将Ⅱ组内含子插入相应特异的Ag43基因碱基1812/1913之间。图3结果表明,用Nhe I 和Hind Ⅲ二种酶切方法均将重组质粒pACD4K-Ag43切出预想的片段,同时按照图1的PCR 扩增策略能够从诱导Ag4基因敲除菌中扩增到相应的PCR 反应产物,这3 种PCR 产物的DNA 序列经基因测序分析也发现Ag43 基因的1 812/1 813位点间确实插入了一段Ⅱ组内含子的DNA 序列,说明本研究获得了Ag43基因正确敲除的新的菌株JM109(△Ag43)。
Ag43是大肠埃希菌表面表达的一个独特分子,该分子α结构域受热至一定温度后就能从β结域分离,溶于加热的水溶液中,离心将菌沉淀后液体上清中就含有单一的α结构域分子。细菌鞭毛和菌毛是典型的细菌表面抗原分子,有人已经设想利用细菌鞭毛或菌毛作为细菌表面表达嵌合抗原分子的载体[8]。但是,利用细菌鞭毛或菌毛表达嵌合抗原分子存在较明显的缺点,主要表现为表达的分子受到表达分子大小的制约,插入外源分子也会影响载体蛋白的结构和功能,同时还降低载体的拷贝数[8]。另外,大多数细胞表面表达的抗原分子还受限于宿主细菌的背景,将这类细胞表面表达分子作为外源抗原分子的展示分子不具有普适性。显然,作为外源抗原分子的展示载体的细菌表面抗原分子必须能够适应较大的外源插入分子,同时还应具有较高的拷贝数及常见表达宿主的普适性。研究结果表明,一个大肠埃希菌细胞表面就有多达50 000个Ag43抗原分子,利用基因工程将Ag43构建成一个重组原核表达载体后,发现这一重组原核表达载体也能够在肺炎克雷伯杆菌和绿脓杆菌中大量表达,表达外还能够改变宿主细菌的形态结构[7]。此外,将Ag43作为外原抗原分子的表达载体比菌毛载体具有更好的表达能力,菌毛载体只能表达20~30个氨基酸的外源抗原分子,但Ag43载体表达的外源抗原分子则可高达500个氨基酸[7]。研究结果还表明,Ag43的免疫原性很强,同时还具有很多的T和B淋巴细胞抗原表位,可以促使免疫系统产生高强度的免疫应答[9]。本研究获得的Ag43基因敲除菌JM109(△Ag43)来自常用基因程表达宿主菌JM109,如果同时和一个可以表达Ag43嵌合分子的表达载体结合,二者就可以构成一个制备外源抗原分子的嵌合蛋白疫苗的表达系统,在制备疫苗和研究Ag43基因功能等方面都具有良好的应用前景。
[1] Sherlock O,Dobrindt U,Jensen JB,et al.Glycosylation of the self-recognizing Escherichia coli Ag43autotransporter protein[J].J Bacteriol,2006,188(5):1798-1807.
[2] Wallecha A,Correnti J,Munster V,et al.Phase variation of Ag43is independent of the oxidation state of OxyR[J].J Bacteriol,2003,185(7):2203-2209.
[3] Ulett GC,Webb RI,Schembri MA.Antigen-43-mediated autoaggregation impairs motility in Escherichia coli[J].Microbiology,2006,152(Pt 7):2101-2110.
[4] Kjaergaard K,Schembri MA,Hasman H,et al.Antigen 43from Escherichia coli induces inter-and intraspecies cell aggregation and changes in colony morphology of Pseudomonas fluorescens[J].J Bacteriol,2000,182(17):4789-4796.
[5] Gabig M,Herman-Antosiewicz A,Kwiatkowska M,et al.The cell surface protein Ag43facilitates phage infection of Escherichia coli in the presence of bile salts and carbohydrates[J].Microbiology,2002,148(Pt 5):1533-1542.
[6] Roche A,McFadden J,Owen P.Antigen 43,the major phase-variable protein of the Escherichia coli outer membrane,can exist as a family of proteins encoded by multiple alleles[J].Microbiology,2001,147(Pt 1):161-169.
[7] Kjaergaard K,Hasman H,Schembri MA,et al.Antigen 43-mediated autotransporter display,a versatile bacterial cell surface presentation system[J].J Bacteriol,2002,184(15):4197-4204.
[8] Rani DB,Bayer ME,Schifferli DM.Polymeric display of immunogenic epitopes from herpes simplex virus and transmissible gastroenteritis virus surface proteins on an enteroadherent fimbria[J].Clin Diagn Lab Immunol,1999,6(1):30-40.
[9] Henderson IR,Owen P.The major phase-variable outer membrane protein of Escherichia coli structurally resembles the immunoglobulin A1protease class of exported protein and is regulated by a novel mechanism involving Dam and oxyR[J].J Bacteriol,1999,181(7):2132-2141.
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
昆明医科大学学报(2021年8期)2021-08-13
内蒙古师范大学学报(自然科学汉文版)(2021年3期)2021-06-01
生物信息学(2020年1期)2020-05-16
生物工程学报(2019年6期)2019-07-10
生物学通报(2019年1期)2019-02-15
中国棉花(2018年6期)2018-07-06
现代检验医学杂志(2016年2期)2016-11-14
天津科技大学学报(2016年1期)2016-02-28
中华灾害救援医学(2015年7期)2016-01-07
