
28种植物种子形态学性状及其萌发对绵羊瘤胃消化的反应
2015-02-25 08:39杨洁晶万娟娟娜丽克斯任爱天鲁为华
草业学报 2015年2期
杨洁晶,万娟娟,娜丽克斯,任爱天,鲁为华
(石河子大学动物科技学院,新疆 石河子832003)
28种植物种子形态学性状及其萌发对绵羊瘤胃消化的反应
杨洁晶,万娟娟,娜丽克斯,任爱天,鲁为华*
(石河子大学动物科技学院,新疆 石河子832003)
摘要:以天山北坡低山带的主要植物种子作为研究对象,观测和分析了该区的28种(禾本科、藜科、豆科、菊科、蓼科、蔷薇科、唇形科、葱科、石竹科、蒺藜科和苋科)植物种子的形态特征及其生物学特性,并对种子进行绵羊瘤胃消化实验,探讨了种子形态特征和消化前后萌发行为之间的关系。结果表明,28种植物种子的形态特征表现出丰富的多样性,种子的重量变化范围大,最大值是最小值的22倍。种子的长、宽和高的变化范围也较大,最大值分别是最小值的14,24和22倍。形状指数的测定结果表明种子主要以扁平状和近似球状为主,大部分植物的种子具有芒和果翅等附属结构,个别种子具有粘液结构。禾本科、藜科和豆科植物种子形态学特征可作为表征亲缘关系的指标,分类结果具有很好的一致性,其他科属植物种类较少,形态学特征分类和亲缘关系分类之间的一致性未能得到很好地反映。种子吸水过程均分为3个阶段,即快速吸水、缓慢吸水和稳定阶段。豆科植物种子的发芽率很低,仅在 4.2%~15.0%之间,菊科植物的萌发率为97.0%,6种藜科植物种子的萌发率为41.0%~97.0%,10种禾本科植物种子的萌发率为 53%~99%。菊科、藜科和禾本科萌发率要明显高于豆科。经过瘤胃消化后的种子,除了豆科种子其萌发率提高之外,其他种子萌发都受到了明显抑制,但仍有部分种子存活并能成功萌发,这为种子消化道传播提供了可能。消化前原始种子萌发率与其形状指数和重量存在显著相关,消化后种子萌发率与种子形状指数和种子长度存在显著相关。并且亲缘关系接近的类群种子其消化前后的萌发行为具有趋同性。
关键词:种子;形态特征;吸水特性;瘤胃消化;萌发行为
种子生物学特性和种子传播之间的关系一直是植物生态学重点研究的内容。种子生物学特性包括种子形态学、生理学及萌发行为,而上述因素直接影响幼苗存活能力。而种子的传播方式则受到种子生物学特性的直接影响,尤其受到种子形态学特征(如传播附属结构)的影响则更为直接,根据种子形态学特征可将种子传播(扩散)类型分为风扩散型、风扩散兼动物体外扩散型、水流扩散型、动物消化道扩散型等。而动物消化道传播被认为是传播效率较高的类型,因为它一方面有利于种子的远距离传播开拓新生境避免同胞竞争,一方面可以通过消化道作用使种子休眠性被打破而提高萌发率。目前有关动物对植物种子的消化道传播已经得到了广泛的关注[4-6]。
从理论上讲,种子性状也是植物分类中主要的分类指标,在进化水平上,近缘物种的种子性状会表现出明显的趋同性,进而会对环境选择压力产生趋同适应,比如荒漠地区的藜科(Chenopodiaceae)植物会在种子形状上表现出明显的二(多)型性[8-9],豆科(Leguminosae)植物种子的普遍硬实特征[10],菊科(Asteraceae)、十字花科(Brassicaceae)植物的黏液繁殖体等[11]。这种在进化水平上的趋同适应表明具有亲缘关系的植物群体在一定的同质选择压力作用下会表现出明显一致的适应性对策,进而能够保证物种在同质环境条件下的延续以及多样性特征。因此,从植物种子的群体特征去挖掘物种对环境条件的反应是目前研究的热点。
天山北坡低山带广泛分布于天山北坡低中山带及低山丘陵区,是放牧家畜冷季牧场,在整个草原畜牧业生产中居于非常重要的地位[2]。天山北坡低山带植物种类多,是家畜赖以生存的基础,尤其是一些优良牧草,如蒿类半灌木、豆科、禾本科、藜科牧草等。除了具有畜牧业生产功能外,在防止水土流失、防风固沙,维持草地生态平衡和平原绿洲区的生态安全等方面发挥着不可替代的作用,生态功能作用十分重要[13]。由于该区域植物种类丰富且是天然草地放牧畜牧业的主要放牧地,因此,植物和放牧家畜之间的相互作用非常剧烈,这就为阐明植物和家畜之间的关系创造了良好的条件。
本文以天山北坡中段低山带的28种常见牧草种子为研究对象,对其形态学以及萌发行为等基本性状进行初步分析,并利用Angiosperm Phylogeny Group III(APG III)分类系统阐明各物种在进化水平上的亲缘关系,进一步通过瘤胃瘘管消化试验来明确经家畜局部消化道作用对种子萌发行为的变化,试图回答以下3个问题,1)28种植物种子的形态学性状特征如何?2)自然状况下种子萌发行为在进化水平上是否存在趋同性?3)经局部消化道作用后,种子萌发行为会发生什么变化,这种变化在近缘种类群中表现趋势是否相同?
1材料与方法
1.1 研究区概况
28种植物种子的采集区位于天山北坡中段石河子南山紫泥泉处的低山带,为天山北坡中段山地的典型代表区,整个低山由原始黄土覆盖,但在一些局部陡坡和断裂层处见有基岩裸露,海拔范围在1200~1800 m之间。该地区具有明显的温带大陆性气候特征,年降水量240~360 mm,蒸发量大于降水4~5倍或以上,降水多集中在6-8月,其占年降水量近50%,年平均气温5℃,无霜期120~140 d。整个草地植被发育好,群落结构复杂,生产力水平较高。
1.2 试验材料
本研究主要选择作为家畜主要饲草的植物种子或果实作为研究对象,共计11 科28种(表 1)。根据不同牧草种子的成熟时期,在2010年6-10月对28种植物种子进行采集。种子采集后于室内风干、筛选后置牛皮纸信封常温贮藏。各物种科种信息见表1。
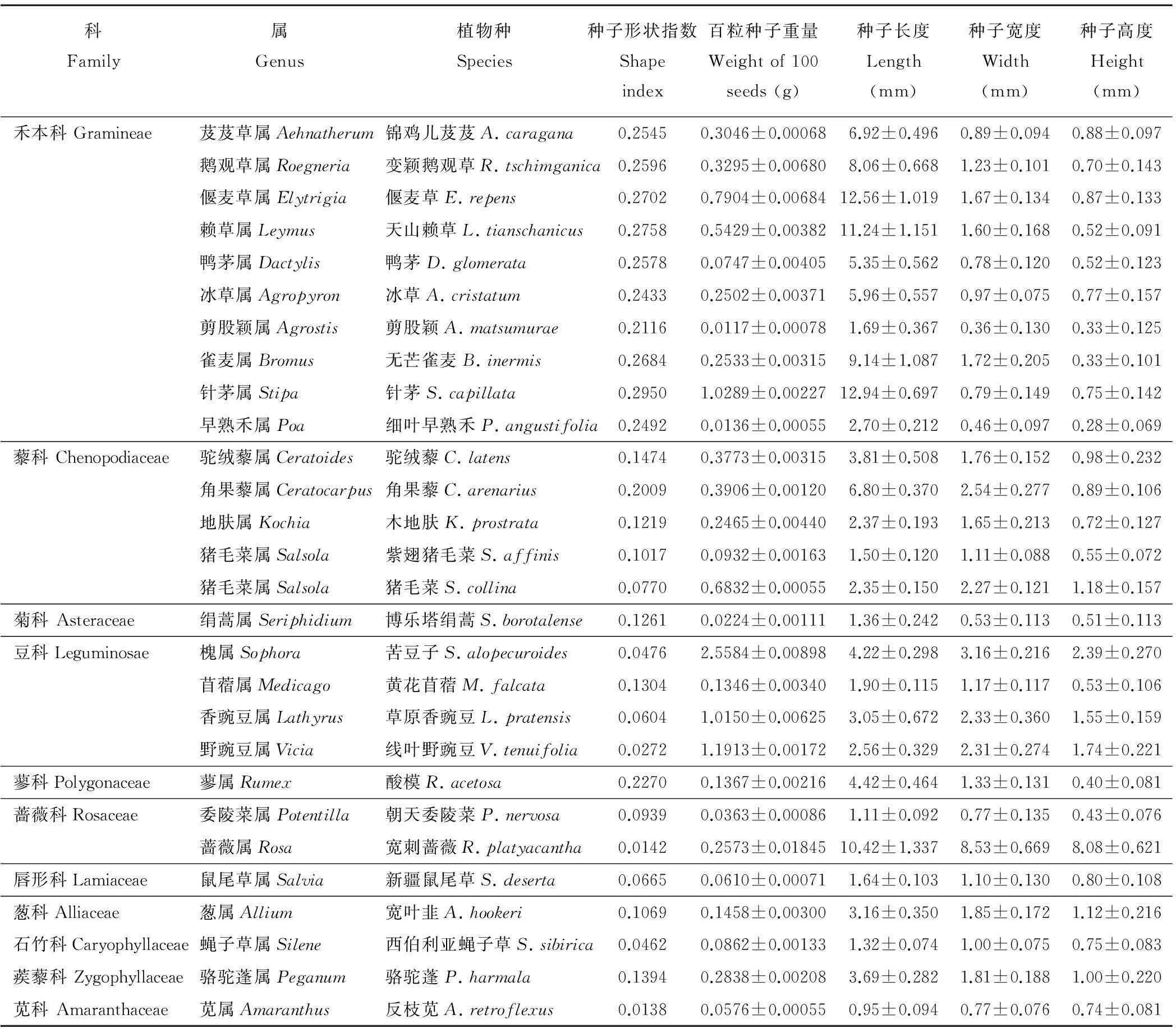
表1 28种植物的种子形态学特征
1.3 研究方法
1.3.1种子形态特征的测定用1/10000 电子天平和体视显微镜测量种子的百粒重,长(L)、宽(W)和高(H),并观察有无附属结构。然后参考Thompson等[14]的计算方法根据种子的长、宽和高计算种子形状指数,具体见公式(1)。
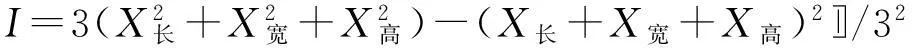
(1)
式中,I为种子形状指数,X为各种子的长、宽、高三维指标。
1.3.2种子吸水特性选取50粒种子,置于培养皿内,用蒸馏水浸润。每个试验重复3次。称量吸胀不同时间的种子湿重,根据公式(2)计算种子的吸水率:
吸水率(%)=(湿重-干量)/干重]×100
(2)
1.3.3种子的瘤胃消化瘤胃瘘管尼龙袋法[15],选用体重为(28.0±2.97) kg左右的3只瘤胃瘘管羊,代谢笼内单笼饲养。将植物种子放入尼龙袋内(尺寸为5 cm×8 cm,孔径在40~50 μm之间),用尼龙线扎口后绑在铁链子上。将尼龙袋在晨饲前(9:00)由瘤胃瘘管投入到瘤胃腹囊中,链条末端用尼龙绳连接一铁夹子,固定在瘤胃瘘管外部被毛上,以防止链条脱落入瘤胃内。然后正常饲喂。24 h后,于晨饲前分别从3只羊瘤胃内各取出1个尼龙袋(代表3个重复)。用自来水冲洗干净后,放在实验室内自然干燥后进行萌发实验。
1.3.4种子萌发特性采用纸上法(TP)[16],将自然状态下和经过瘤胃消化后的每种种子随机选取100 粒,放入双层滤纸培养皿中进行萌发,重复3次。根据采集地植被生长气候条件设置8 h 15℃/16 h 25℃变温和光照,每24 h进行萌发检测、统计萌发个数,胚根出现1~2 mm即可认为已经萌发,直到连续15 d不再有发芽种子出现时为止,最后以公式(3)计算萌发率。
萌发率(%) =发芽种子数/种子总数×100
(3)
1.4 数据分析
将植物种拉丁文输入Phylomatic(Version 3.0)中建立系统进化树,明确物种之间的亲缘关系。利用元分析软件RevMan 5.0中的固定效应模型进行消化前后种子萌发率效应值的计算和分析,将效应值划分为5个区间,即小于0.01,0.01~0.1,0.1~1,1~10,10~100,效应值大小在图中用黑色方块表示,若效应值大小位于0.01~1则表明对种子萌发率起抑制作用,而当其位于1之后,则表明对萌发率起促进作用,具体效应值计算方法见公式(4)。所有数据采用 Excel 软件录入并进行初步统计,用DPS 7.5进行形态学模糊聚类分析。
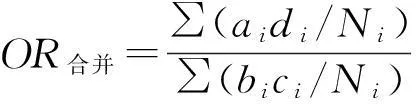
(4)
式中,OR为效应值;ai指消化后萌发种子数;bi指消化后未萌发的种子数;ci指消化前萌发种子数;di指消化前未萌发的种子数;Ni指萌发总数。
2结果与分析
2.1 种子形态学特征
28种植物种子的形态学特征见表1,通过分析结果表明,平均重量为0.403 mg,但物种间种子重量差别很大,单粒重量最小的为剪股颖(0.0117 mg),最大的为苦豆子(2.5584 mg),二者之间相差约22倍。
种子的长、宽、高的大小变化范围很大,种子的平均长度为4.76 mm,最长的为针茅(12.94 mm),最短的为反枝苋(0.95 mm),二者之间相差约为14倍。种子的平均宽度为1.73 mm,最宽的为宽刺蔷薇,为8.53 mm,最窄的为剪股颖,为0.36 mm,二者之间相差约为24倍;种子高度的平均值为1.08 mm,最高的为宽刺蔷薇,为8.08 mm,最低的为剪股颖,为0.33 mm,最大值与最小值之间相差约为22倍。
通过对28种植物种子的形状观测结果表明,当种子形状指数的值大于0.2时,种子的形状为扁平状,主要包括12种植物种子,分别是锦鸡儿芨芨、变颖鹅观草、偃麦草、天山赖草、鸭茅、冰草、剪股颖、无芒雀麦、针茅、细叶早熟禾、角果藜和酸模;当种子形状指数的值小于0.1时,种子的形状为近似球体状或椭圆形,主要包括9种植物种子,其分别为苦豆子、草原香豌豆、线叶野豌豆、猪毛菜、朝天委陵菜、新疆鼠尾草、西伯利亚蝇子草、反枝苋和宽刺蔷薇;当种子形状指数的值介于0.1~0.2之间时,种子的形状介于球状体和扁平状之间,主要包括7种植物种子,分别是黄花苜蓿、驼绒藜、木地肤、紫翅猪毛菜、博乐塔绢蒿、骆驼蓬和宽叶韭。其中,禾本科植物种子多为扁平状(种子形状的数值均大于0.2),豆科植物种子多为球体状(种子形状的数值均小于0.1),其他植物种子的形状介于两者之间。
通过对28种植物种子的附属结构观测结果表明,有芒植物种子主要包括锦鸡儿芨芨、变颖鹅观草、偃麦草、天山赖草、鸭茅、冰草、无芒雀麦、针茅;有果翅的植物种子主要包括木地肤、猪毛菜、紫翅猪毛菜;有刺的植物种子仅有一种,即角果藜;种子表面具毛的种子为驼绒藜。其中,有附属结构的种子占总数的46.43%。另外,对28种植物种子用蒸馏水湿润发现,2种植物种子具有分泌粘液的特性,分别是新疆鼠尾草和博乐塔绢蒿。
用长、宽、高、重量作为聚类变量,对28种植物种子进行聚类分析,结果表明,当距离小于8.92时可分为4类:锦鸡儿芨芨、细叶早熟禾、变颖鹅观草、鸭茅、剪股颖、偃麦草、赖草、冰草、针茅、无芒雀麦等10种禾本科牧草聚为一类;驼绒藜、朝天委陵菜、新疆鼠尾草、紫翅猪毛菜、黄花苜蓿、宽叶韭、西伯利亚蝇子草、反枝苋、角果藜、木地肤、骆驼蓬、博乐塔绢蒿、酸模聚为一类;猪毛菜、草原香豌豆、线叶野豌豆聚为一类;苦豆子、宽刺蔷薇为1类,具体结果见图1A。
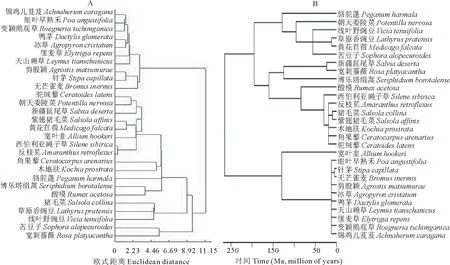
图1 28种植物种子形态学聚类(A)以及亲缘关系(B)图Fig.1 Morphological clustering(A) and phylogenetic tree (B) of 28 plants species
进一步利用Phylomatic(Version 3.0)软件对28个物种的亲缘关系进行划分并以系统进化树的形式反映出来,结果见图1B。其中进化枝的长度代表物种的进化时间,单位:Ma(million of years)。两种分类结果表明,28种植物中的10种禾本科植物种学指标聚为一类,表现出与禾本科相似的特征。但由于其他科属植物种类较少,因此形态学特征分类和亲缘关系形态学特征可作为表征亲缘关系的指标,分类结果具有很好的一致性。藜科和豆科的大部分也可通过种子形态学分类之间的一致性得到很好地反映。
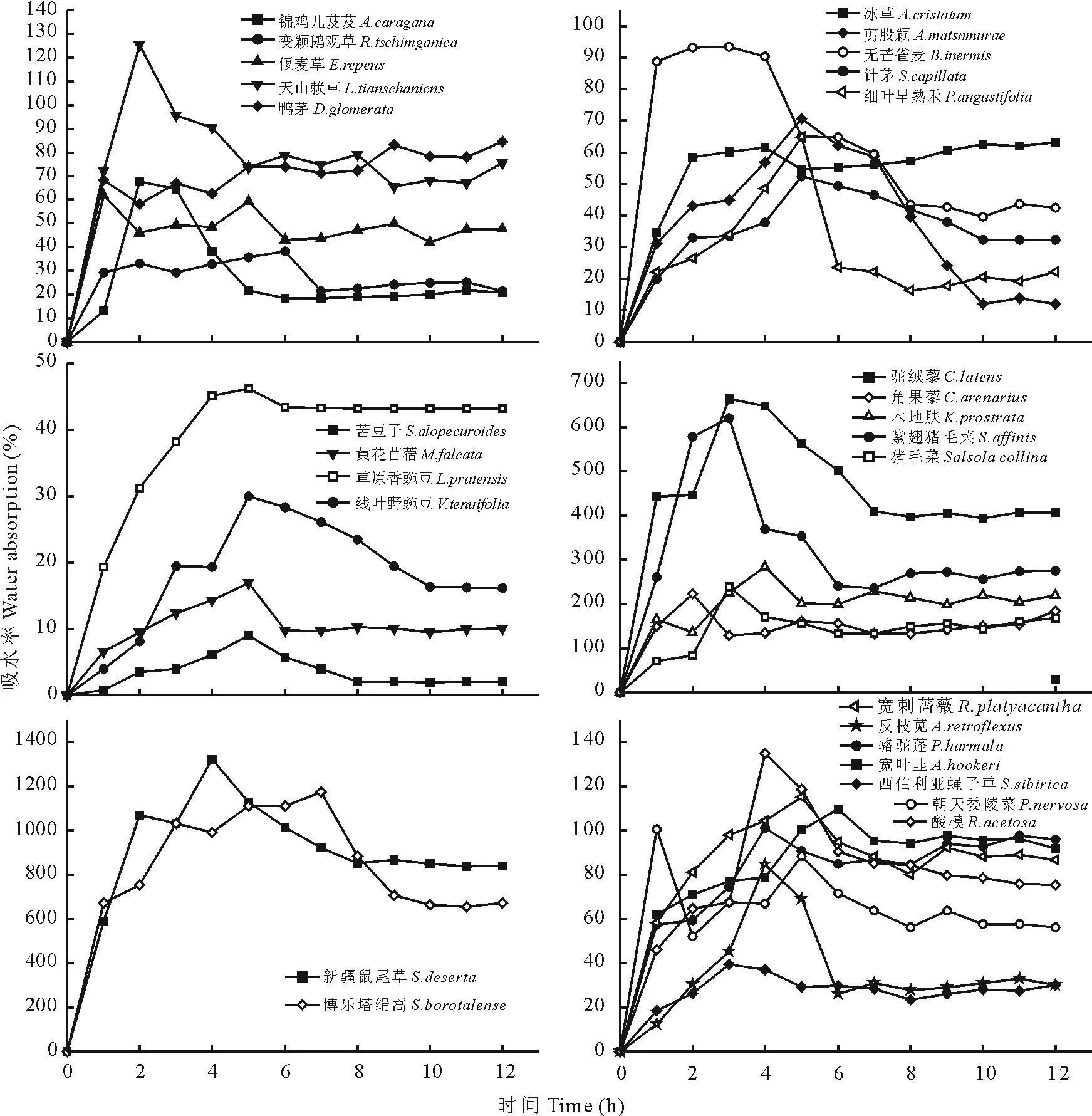
图2 28种植物种子的吸水特性Fig.2 Water-absorbing curve of 28 species
2.2 种子吸水特性
种子萌发成功与否与其萌发前期的吸水过程密切相关,通过对种子吸水曲线的测定结果表明,28种植物种子的吸水过程基本一致,都包含3个阶段,即快速吸水阶段、缓慢吸水阶段和稳定阶段。但是,在不同物种之间,种子各吸水的阶段所持续的时间是不同的,而在同一物种内,种子各吸水的阶段所持续的时间是基本相似的。禾本科植物的种子大致在4~5 h之前为种子的快速吸水阶段, 5~8 h之间为种子的缓慢吸水阶段, 这个阶段的吸水量占总吸水量8 h之后为种子的饱和阶段;藜科植物种子在3 h之前为种子的快速吸水阶段,3~10 h之间为种子的缓慢吸水阶段,10 h后为种子的饱和阶段;豆科植物种子在5~6 h之前为种子的快速吸水阶段,6~8 h之间为种子的缓慢吸水阶段,8 h之后为种子的饱和阶段;具有粘液结构的种子(绢蒿和鼠尾草)在2 h之前为种子的快速吸水阶段,2~10 h为种子的缓慢吸水阶段,10 h之后为种子的饱和阶段;剩余7种植物种子为不同的物种,所以种子各吸水阶段所持续的时间也是不相同(图2)。
通过对28种植物种子吸水率的测定可以看出,具有粘液结构的两种植物种子鼠尾草和绢蒿的吸水率最高,平均吸水率分别为943.94%和837.07%;而豆科植物中的苦豆子吸水率最低,平均吸水率为3.59%。方差分析显示,禾本科、藜科和豆科植物种子的吸水曲线基本相似,但其平均吸水率之间差异显著(P<0.05)。因此,即使是同一科不同植物种之间种子的吸水曲线变化相似,但在平均吸水率之间却不相同。
2.3 种子的萌发特性
28种植物种子发芽率的变化范围在4.00%~99.00%之间(表 2), 萌发率较高的植物种有剪股颖、 无芒雀麦、紫翅猪毛菜、猪毛菜、博乐塔绢蒿、酸模和反枝苋;萌发率较低的植物种是苦豆子、黄花苜蓿和线叶野豌豆。
将这28种植物种子萌发率进行分组,包括高萌型(萌发率>80%),中间型(55%~80%),次低萌型(20%~55%),低萌型(萌发百分率<20%)。高萌型的植物有14种,分别是变颖鹅观草、偃麦草、赖草、鸭茅、剪股颖、无芒雀麦、紫翅猪毛菜、猪毛菜、博乐塔绢蒿、朝天委陵菜、酸模、西伯利亚蝇子草、骆驼蓬和反枝苋;萌发率在55%~80%之间的为中间型,共5种,分别是锦鸡儿芨芨、冰草、细叶早熟禾、宽叶韭和新疆鼠尾草;萌发率在20%~55%之间的为次低萌型,有5种,分别是针茅、驼绒藜、角果藜、木地肤和宽刺蔷薇;萌发率小于20%的为低萌型,共4种,分别是苦豆子、黄花苜蓿、草原香豌豆和线叶野豌豆。
瘤胃消化24 h后,除了豆科牧草种子萌发率有所上升外,其余各科牧草种子萌发率都有所下降,其中禾本科10种、藜科5种、菊科的绢蒿、蔷薇科的委陵菜、蓼科的酸模、唇形科的新疆鼠尾草、石竹科的西伯利亚蝇子草萌发率下降趋势明显。而宽刺蔷薇、宽叶韭、骆驼蓬、反枝苋经过消化后种子萌发率尽管有所下降但趋势不明显。
进一步对原始种子、瘤胃消化后的种子萌发率与种子形状指数、重量、长度、宽度和高度进行回归分析,结果表明,原始种子萌发率与种子形状指数、种子重量存在显著相关性,其中与种子形状指数符合形如y=0.191exp(-8176/x)的对数函数关系式,种子形状指数随着萌发率呈现对数增长。与种子重量符合形如y=7.770x-0.8773的幂函数关系式,种子重量随萌发率逐渐降低。瘤胃消化后的种子萌发率尽管大部分明显降低,但仍然和种子形状指数、长度存在明显相关,且分别符合形如y=0.5083x-0.4954和y=30.3592x-0.7960的幂函数关系式,种子形状指数、重量随萌发率降低(图3)。
2.4 基于效应值(OR)的萌发行为分析
将物种亲缘关系树型图与RevMan 5.0计算的效应值森林图结合,进行消化前后种子萌发率的变化分析。结果表明,合并后的总体效应值OR为0.09(图4),95%区间范围为0.07~0.10,且经过u检验和卡方检验,其结果均为P<0.01,说明总体效应值具有明显的统计学意义。总体上,经过瘤胃消化24 h后种子的萌发率均受到了明显的抑制作用。同时,根据效应值森林图统计学含义,有5种牧草种子的效应值横跨横轴尺度为1的无效竖线,因此,这5种牧草种子数据无统计学意义,即在此处不能明确得出消化处理对种子萌发率是否有促进或者抑制效应,这5种植物分别为宽刺蔷薇、苦豆子、黄花苜蓿、草原香豌豆、线叶野豌豆。另外,根据亲缘关系中各亲缘类群的表现来看,亲缘关系接近的牧草种子其效应值呈现出明显的趋同性,这也进一步说明,亲缘关系在某种程度上决定了物种某一性状对环境选择压力所采取适应对策的一致性。

表2 28种植物种子消化前后的萌发特性
注:表内数据为平均值±标准差,同行不同小写字母表示差异显著(P<0.05),大写字母表示差异极显著(P<0.01)。
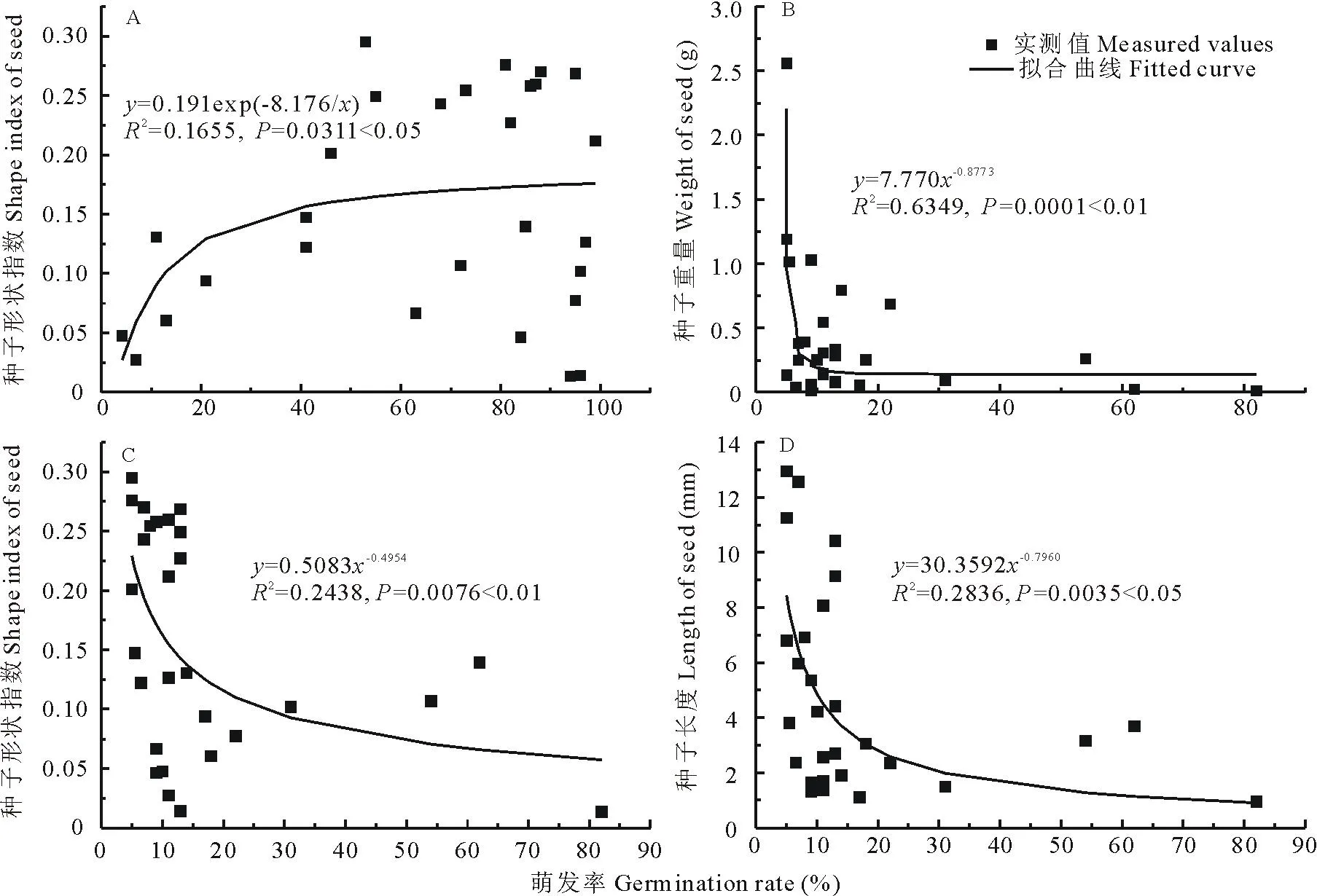
图3 28种牧草种子瘤胃消化前(A,B)和消化后(C,D)种子萌发特征与种子形态之间的关系Fig.3 Relationship of between germination rate and seed traits under original (A,B) and digested (C,D) seeds of 28 species
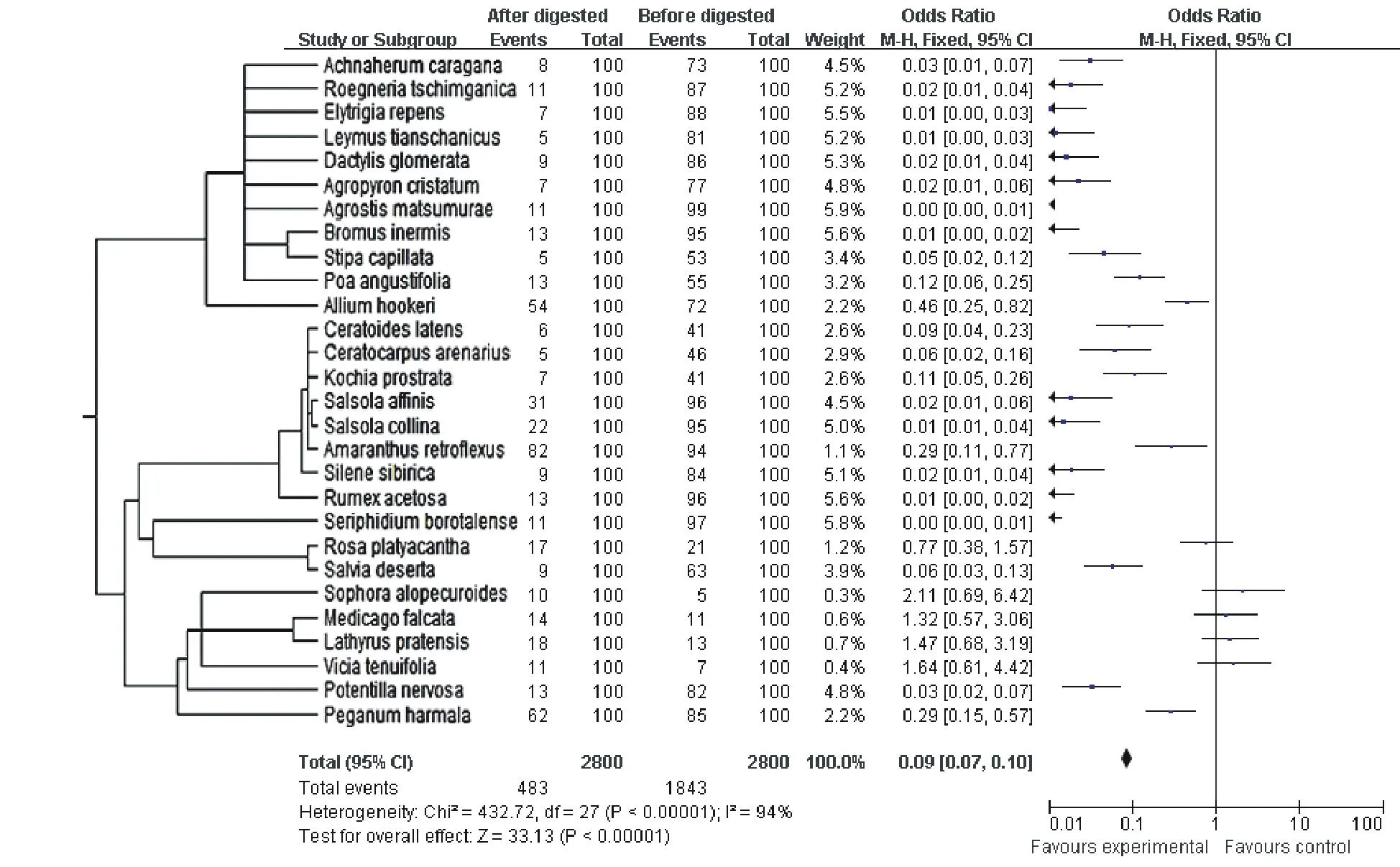
图4 28种植物种子经瘤胃消化前的后萌发率效应值Fig.4 Odds ratio of original and digested seeds of 28 species
Note:Values are given as mean±SE, the different lowercase letters indicate significant difference(P<0.05), different capital letters indicate significant difference(P<0.01).
3讨论与结论
3.1 讨论
种子的形态特征是种子与其环境长期相互作用的结果,不同植物种子的形态特征不仅会影响种子的萌发特性同时对种子的散布方式也有一定的影响。通过对28种植物种子形态特征进行比较发现,种子的重量,形状之间的差异较大,同时具有不同的附属结构,这些形态差异会影响种子萌发和幼苗建植。研究发现,种子的形状为球形或者近似球形时,种子流动性好,可实现较远距离的传播,从而使种子能在适宜的生境条件下萌发和存活[17-18]。芒的存在不仅能够降低种子降落的速度,而且能够增大种子与土壤充分接触面积,增加种子对水分的吸收,从而增加种子的萌发几率。而藜科植物猪毛菜、紫翅猪毛菜和木地肤等具有明显的果翅,这种结构更有利于种子在空中飞行,使种子能够实现远距离的传播,同时也可以增加种子与地面的摩擦,从而更有利于实现种子的散布与萌发。
种子在成熟以后,经过干燥其自身的含水量很少,组织之间会变得紧密坚实,细胞内部的内含物由于水分少,呈现出干燥的凝胶状态,从而使其自身代谢能力减弱。因此,当种子与水分直接接触时,种子就会快速地吸水并且膨胀。由于不同植物种子的形态特征不同导致其吸水过程的差异,从而导致种子萌发特性之间存在差别。通过对28种植物种子吸水特性和萌发特性进行比较发现,种子的吸水曲线变化是一致的,均包括快速吸水阶段、缓慢吸水阶段和稳定阶段。胡卉芳和李青丰[19]对5种禾本科牧草的吸水特性进行观测发现,5种禾本科牧草种子吸水曲线变化一致,但在种子吸水量之间存在显著差异。本研究中也得到了相似的结论,说明虽然植物种子的吸水曲线变化是相同的,但是由于种子的形态特征不同,种子的吸水率之间也是不相同的。同时,在本研究中具有粘液结构的两种种子吸水率最高,张桂红和贺学礼[20]对8种绢蒿属植物种子的吸水特性进行观测得到了类似的结论。对具有粘液结构的种子来说,粘液结构不仅能够使种子大粒化,增加种子的重量,使其不易被风吹走,确保种子在合适的条件下萌发,同时也能够增大种子与土壤水分的接触面积,有利于种子对周围土壤中水分的吸收和利用,从而更有利于种子的萌发,增加后代的生存几率。阿衣古力等[21]对伊犁绢蒿去粘液和有粘液种子的萌发率和萌发速度进行研究发现,种皮的粘液结构对种子萌发率和萌发速度有显著的影响,Huang 和 Gutterman[22]对油蒿种子及白沙蒿[23]粘液结构对种子萌发率影响的研究中得到了相似的结论,本研究结果与此一致。这主要是由于种皮的粘液结构中的物质,不仅能够促进种子对水分的吸收,同时,也能够增强种子的吸水和保水的能力,从而对种子的萌发及幼苗的发育更有利,而且粘液中的化学物质也有利于幼苗的顺利生长。本研究也发现植物种子表面有附属结构,例如具果翅、芒的种子吸水性要远远大于无附属结构的植物种子。对于豆科植物种子来说,硬实现象是普遍存在的,即豆科种子的物理性休眠,使得它对水分的吸收能力较弱进而影响萌发率,本研究中4种豆科植物种子吸水率和萌发率均显著低于其他植物种子,则再次充分证明了这一点。
种子形态学特征也会影响到种子萌发率,并且直接影响幼苗的建植。本研究通过对28种植物种子的萌发特性进行比较发现,未被消化种子的形状指数随着萌发率的增加而增加,即种子越圆,萌发率越低。这一结论在有关种子的形状与土壤种子库寿命关系的研究中已经得到了证实,圆形种子易形成持久种子库[24]。Thompson等[14]研究 97种英格兰植物也具有同样结论,并且认为这个持久性至少是5年。Grmie[25]也认为圆形种子与长形种子比较,在土壤中不萌发或很少萌发,存留时间长可形成持久土壤种子,这就意味着它们具有适应频繁干扰的能力,在土壤中等待适合的时机来萌发,属于“保守型”[26]。本研究中原始种子萌发率与种子重量之间存在对数函数关系,说明种子越小萌发率越高,且萌发速度较快。宗文杰等[27]对高寒草甸中51种菊科种子的萌发进行研究,发现小种子具有较高的萌发率和较快的萌发速度。而张蕾等[28]对青藏高原的31种杂草种子研究发现,随着种子大小的增加,萌发率和萌发速度减小,萌发开始时间和萌发高峰时间推迟。种子大小反映了其所能提供给幼苗存活的资源量的多少[29]。根据种子大小数目权衡模型,在有限的资源下,植物种子大小与其产生的种子数目呈负相关。小种子产生大量的小种子,其扩散能力较强,通过扩散等方式拓殖到一些大种子植物不能到达的领域。为了增加适合度,小种子必须尽可能多地占领适宜萌发的斑块,并在斑块中大量萌发来增加幼苗成活率进而增加整体适合度。因此,一般小种子的萌发率都较高,属于“冒险型”[30]。
研究种子萌发规律及对策,有助于认识和阐明物种进化及其生态适应特征[31]。植物在长期的进化过程中,除受到自身遗传因素的影响外,还受到外界环境的影响。一般来说,亲缘关系接近的物种,种子的形态特征较相似,并且对环境条件采取趋同性的适应对策。本研究中,通过种子形态学聚类和Phylomatic亲缘关系分类系统[32]分类结果,首先充分证实了亲缘关系接近形态学特征趋同这一观点。同时,经过瘤胃消化后亲缘关系接近的植物种子对消化的反应也有趋同特征,这就证明了种子性状及对环境条件的反应明显对亲缘关系具有依赖性。但同时我们也可看出,仅通过消化前后种子萌发率高低变化来分析植物种子对消化道传播的响应是不科学的[33]。因此,通过元分析技术中的固定效应模型来计算效应值,然后根据效应值的区间范围来判定瘤胃消化对种子萌发行为是抑制还是促进是比较可靠的统计方法,并进一步根据亲缘类群与消化前后种子萌发行为之间关系的结合,可进一步证明种子在相同的选择压力下在同一亲缘群体内的反应是否具有趋同特征,这也正是众多学者在研究有关种子消化道传播过程中所采取的研究方法。因此,元分析技术不仅可用在综述撰写的整合分析过程中,也可对实验性的多个数据进行严密的数理统计分析,并可取得良好的效果。
3.2 结论
28种植物种子形态学特征表现出丰富的多样性特征,种子(果实)的重量变化范围大,最大值是最小值的22倍。种子的长、宽和高的变化范围也较大,最大值分别是最小值的14,24和22倍。形状指数的计算结果表明种子主要以扁平状和近似球状为主,大部分植物的种子具有芒和果翅等附属结构,个别种子具有粘液结构。根据种子形态学特征聚类和亲缘关系分析的结果表明,相同科属的种子可聚为一类,种子形态特征可作为判定物种亲缘关系的重要依据。
种子萌发前期的吸水过程均分为3个阶段,即快速吸水、缓慢吸水和稳定阶段。同一亲缘类群的植物种子吸水过程相似,但是在吸水量上存在差别。豆科植物种子的吸水率及吸水速度最低。具粘液结构的种子吸水速率和吸水量均最高。
消化前的豆科植物种子的萌发率很低,仅在 4.2%~15.0%之间,属于典型的低萌型种子,菊科植物的萌发率为97.0%,6种藜科植物种子的萌发率为41.0%~97.0%,10种禾本科植物种子的萌发率为 53%~99%。菊科、藜科和禾本科萌发率要明显高于豆科。原始种子萌发率与其形状指数和重量存在显著相关,其萌发行为对形态学特征具有明显依赖,相同形状的种子萌发行为趋于一致。
经过瘤胃消化后的种子,其萌发率与种子形状指数和种子长度存在显著相关,且均符合幂函数模型。所有经瘤胃消化后的种子,除了豆科种子萌发率提高但通过效应值计算无统计学意义之外,其他种子萌发都受到了明显抑制,28种植物总体的OR值在95%区间范围内为0.09~0.10之间,表明瘤胃消化后对种子整体上具有明显抑制作用,但仍有部分种子存活并能成功萌发,这为种子消化道传播提供了可能,这些“逃逸”过动物消化的种子可被动物携带实现远距离传播,在拓殖新生境减少同胞竞争方面具有重要的生态学意义。另外,通过消化前后种子的萌发效应值结合亲缘关系树状图进一步发现,亲缘关系接近的物种不仅在形态学特征上具有明显的一致性,而且其消化前后的萌发行为具有趋同性。这就说明物种进化过程中仍然遵循一般的自然规则,即采取进化水平相同的物种总是采取一致的适应对策来应对环境条件造成的选择压力从而能增加自身适合度。
Reference:
[1]Thompson K. Seed and seed banks. New Phytologist, 1987, 106: 23-34.
[2]Gross K, Smith A. Seed mass and emergence time effects on performance ofPanicumdichotomiflorumMichx. across environments. Oecologia, 1991, 87(2): 270-278.
[3]Peco B, Lopez M, Alvir M. Survival and germination of Mediterranean grassland species after simulated sheep ingestion: ecological correlates with seed traits. Acta Oecologica, 2006, 30(2): 269-275.
[4]Yu X J, Xu C L, Wang F,etal. Recovery and germinability of seeds ingested by yaks and Tibetan sheep could have important effects on the population dynamics of alpine meadow plants on the Qinghai-Tibetan Plateau. The Rangeland Journal, 2012, 34(3): 249-255.
[5]Yu Y, Toshiya O, Bayarbaatar,etal. Spatial pattern of grazing affects influence of herbivores on spatial heterogeneity of plants and soils. Oecologia, 2009, 162(2): 427-434.
[6]Bram D, Maurice H. A reassessment of the role of simple seed traits in mortality following herbivore ingestion. Plant Biology, 2010, 13(1): 118-124.
[7]Zhang S T, Du G Z, Chen J K. The present situation and prospect of studies on evolutionary ecology of seed size variation. Acta Ecologica Sinica, 2003, 23(2): 353-364.
[8]Song Y G, Li L, Deng M,etal. Responses of the germination on dimorphic seeds ofSuaedaaralocaspicato salt stress. Acta Ecologica Sinica, 2014, 23(1): 192-198.
[9]Wang X Y, Gao R, Zhou X Q,etal. Fruit-set patterns and burial mechanism for subterranean dispersal units ofCeratocarpusarenarius. Acta Ecologica Sinica, 2014, 23(2): 352-357.
[10]Guo X M, Xu X Y, Meng X D,etal. Bourgeon characteristics of hardseed ofAlibiziajulibrissin durazz and its testa micro-morphology and structure. Journal of Inner Mongolia Agricultural University, 2006, 27(3): 13-18.
[11]Song M F, Liu H L, Zhang D Y,etal. Screening and comparison on myxospermy of 120 species occurring on the Junggar Desert. Journal of Desert Research, 2009, 29(6): 1134-1140.
[12]Xu P. The Grassland Resources and Its Utilization in Xinjiang. Urumqi: Xinjiang Science and Technology and Health Press, 1993.
[13]Zhao W Y. Deterioration, causes and control strategies of grassland resources in Xinjiang, China. Pratacultural Science, 2002, 19(2): 19-22.
[14]Thompson K B, Band S R, Hodgson J G. Seed size and shape predict persistence in soil. Functional Ecology, 1993, 7(2): 236-241.
[15]Han J G. Grassland Science. Beijing: China Agriculture Press, 2007.
[16]State Bureau of Technical Supervision. GB/T 3543.4-1995. Rules for the inspection of the national standard of the People’s Republic of China crop seed germination test. Beijing: China Standards Press, 1995.
[17]Zhang J N, Liu K. Mechanisms for plants detecting the optimum time and place to germinate. Acta Prataculturae Sinica, 2014, 23(1): 328-338.
[18]Yi Z D, Mou Z. Study of arid and semi arid area of main arbor and shrub seed morphological characters and physiological and ecological characteristics. Gansu Forestry Science and Technology, 1990, 2: 16-20.
[19]Hu H F, Li Q F. A study on germination characters with hydration-dehydration treatment in five grass species. Grassland of China, 2001, 23(3): 49-54.
[20]Zhang G H, He X L. Water-absorbing and germinating characteristics of eight species fromSeriphidium(Bess.) Poljak. seeds. Journal of Hebei University (Natural Science Edition), 2008, 28(5): 536-541.
[21]Aygul, Tursunay, Halida,etal. Studies on relationship between the natural seed coat and moisture ofSeriphidiumtransiliense. Arid Zone Research, 2000, 17(4): 54-56.
[22]Huang Z Y, Gutterman Y. Comparison of germination strategies ofArtemisiaordosicawith its two congenera from deserts of China and Israel. Acta Botanica Sinica, 2000, 42(1): 71-80.
[23]Huang Z Y, Gutterman Y, Hu Z H,etal. Seed germination inArtemisiasphaerocephalaI. the structure and function of the mucilaginous achene. Acta Phytoecologica Sinica, 2001, 25(1): 22-28.
[24]Leishman M R, Westoby M. Seed size and shape are not related to persistence in soil in Australia in the same way as in Britain. Functional Ecology, 1998, 12(3): 480-485.
[25]Grmie J P. Plant Strategies, Vegetation Processes, and Ecosystem Properties. UK: John Wiley and Sons, 2001.
[26]Zhao X F, Xu H L, Li Y,etal. Comparison of diaspore shape, size, and mass of main desert plants in the lower reaches of Tarim River of Xinjiang, China. Chinese Journal of Ecology, 2009, 28(3): 411-416.
[27]Zong W J, Liu K, Bu H Y,etal. The mode of seed size variation and the effects of seed size on fifty-one species of composite plants in a alpine meadow. Journal of Lanzhou University (Natural Sciences), 2006, 42(5): 52-55.
[28]Zhang L, Zhang C H, Lv J P,etal. Seed germination characteristics and their correlations with seed sizes of 31 common weeds in eastern Qinghai-Tibet Plateau. Chinese Journal of Ecology, 2011, 30(10): 2115-2121.
[29]Yan Q L, Liu Z M, Li X H,etal. Effects of burial on seed germination characteristics of 65 plant species on Horqin semi-arid steppe. Chinese Journal of Applied Ecology, 2007, 18(4): 777-782.
[30]Smith C C, Fretwell S D. The optimal balance between size and number of offspring. The American Naturalist, 1974, 108(962): 499-506.
[31]Zhang H X, Zhou D W. Research on seed germination ecology. Acta Agrestia Sinica, 2009, 17(1): 131-133.
[32]The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Botanical Journal of the Linnean Society, 2003, 141(4): 399-436.
[33]Traveset A, Verdú M. A meta-analysis of effect of gut treatment on seed germination. Seed Dispersal and Frugivory: Ecology, Evolution and Conservation. Wallingford: CABE, 2001: 339-350.
参考文献:
[1]张世挺, 杜国祯, 陈家宽. 种子大小变异的进化生态学研究现状与展望. 生态学报, 2003, 23(2): 353-364.
[2]宋以刚, 李利, 邓敏, 等. 异子蓬二型性种子萌发对盐胁迫的响应. 草业学报, 2014, 23(1): 192-198.
[3]王喜勇, 高蕊, 周晓青, 等. 角果藜的结实格局及地下散布单位的入土机制. 草业学报, 2014, 23(2): 352-357.
[10]郭学民, 徐兴友, 孟宪东, 等. 合欢种子硬实与萌发特性及种皮微形态与结构特征的研究. 内蒙古农业大学学报, 2006, 27(3): 13-18.
[11]宋明方, 刘会良, 张道远, 等. 准噶尔荒漠120种植物粘液繁殖体的甄别与比较. 中国沙漠, 2009, 29(6): 1134-1140.
[12]许鹏. 新疆草地资源及其利用. 乌鲁木齐: 新疆科技卫生出版社, 1993.
[13]赵万羽. 新疆草地资源的劣化、原因及治理对策. 草业科学, 2002, 19(2): 19-22.
[15]韩建国. 草地学. 北京: 中国农业出版社, 2007.
[16]国家技术监督局. GB/T 3543.4-1995. 中华人民共和国国家标准农作物种子检验规程发芽试验. 北京: 中国标准出版社, 1995.
[17]张佳宁, 刘坤. 植物调节萌发时间和萌发地点的机制. 草业学报, 2014, 23(1): 328-338.
[18]伊祚东, 牟真. 干旱半干旱地区主要乔灌木种子形态特征及生理生态特性的研究. 甘肃林业科技, 1990, 2: 16-20.
[19]胡卉芳, 李青丰. 5种禾本科牧草种子的萌发特性及耐旱性的研究. 中国草地, 2001, 23(3): 49-54.
[20]张桂红, 贺学礼. 8种绢蒿属植物种子吸水和萌发特性. 河北大学学报: 自然科学版, 2008, 28(5): 536-541.
[21]阿衣古力, 吐尔逊娜依, 哈丽旦, 等. 伊犁绢蒿种子天然种衣与水分关系初步研究. 干旱区研究, 2000, 17(4): 54-56.
[23]黄振英, Gutterman Y, 胡正海, 等. 白沙蒿种子萌发特性的研究Ⅰ. 粘液瘦果的结构和功能. 植物生态学报, 2001, 25(1): 22-28.
[26]赵新风, 徐海量, 李媛. 塔里木河下游主要荒漠植物繁殖体的形状、大小与质量比较. 生态学杂志, 2009, 28(3): 411-416.
[27]宗文杰, 刘坤, 卜海燕, 等. 高寒草甸51种菊科植物种子大小变异及其对种子萌发的影响研究. 兰州大学学报: 自然科学版, 2006, 42(5): 52-55.
[28]张蕾, 张春辉, 吕俊平, 等. 青藏高原东缘31种常见杂草种子萌发特性及其与种子大小的关系. 生态学杂志, 2011, 30(10): 2115-2121.
[29]闫巧玲, 刘志民, 李雪华, 等. 埋藏对65种半干旱草地植物种子萌发特性的影响. 应用生态学报, 2007, 18(4): 777-782.
[31]张红香, 周道玮. 种子发芽生态研究. 草地学报, 2009, 17(1): 131-133.
Seed morphology and effect of sheep rumen digestion on germinability of 28 grass of Tianshan
YANG Jiejing, WAN Juanjuan, Narkes Wali, REN Aitian, LU Weihua*
CollegeofAnimalScienceandTechnology,ShiheziUniversity,Shihezi832003,China
Abstract:Seeds from 28 plant species belonging to the Poaceae, Chenopodiaceae, Fabaceae, Asteraceae, Polygonaceae, Rosaceae, Lamiaceae, Alliaceae, Caryophyllaceae, Zygophyllaceae and Amaranthaceae, which are the main plant families present in the four grassland types on the north slope of Tianshan (regional indication would be good, eg. Northwest China), were collected, and their morphological characters recorded and germinability before and after sheep-rumen-passage determined. The seeds of the 28 plant species showed rich diversity in morphological characteristics, and their weights spanned a wide range, with the heaviest being 22 times the weight of the lightest. Similarly for dimensions of length, width and height, and the maxima and minima of the various species differed by a factor of 14, 24, and 22, respectively. A shape index indicated that in most species seeds were either markedly flattened or approximately sphericalin shape. For most species seed morphology was modified by adjacent structures, such as an awnon an adhering lemma in the Poacea, or fruiting wings in some herbs. Some seeds were borne in mucilaginous structures. Morphological characteristics of Poaceae, Chenopodiaceae and Fabacea were taxonomically diagnostic. Seeds of the remaining families did not show any phylogentic consistency in their morphological characteristics. The water absorption of plant seeds during germination can be divided into three stages: a fast water absorption phase, a slow water absorption phase and constant water content phase. The germination rate of legume seeds was very low, ranging from 4.2%-15.0%, while germination rate of Asteraceae species was up to 97%; the germination rate of 6 Chenopodiaceae species ranged from 41%-97%, and of 10 Poaceae species ranged from 53%-99%. After sheep-rumen-passage, the germination rate of Fabaceae seeds was increased, whereas the germinability of seeds of other families was decreased. However, despite reduced germinability after rumen passage, because of the survival of some seeds sheep browsing provides a possible pathway for seed dispersal. The germination rate of seed before rumen digestion was significantly associated with shape index and seed weight, while it was significantly correlated with seed shape index and seed length after rumen digestion. Additionally, there was convergence in germination behavior before and after digestion among seeds that have a close genetic relationship.
Key words:seeds; morphological characteristics; water absorption characteristics; rumen digestion; germination behavior
*通讯作者
Corresponding author. E-mail:winnerlwh@sina.com
作者简介:杨洁晶(1988-),女,河北石家庄人,在读硕士。E-mail:juncai19881004@163.com
基金项目:国家自然科学基金(31360568)和教育部博士点新教师基金(20126518120004)资助。
*收稿日期:2014-08-20;改回日期:2014-09-16
DOI:10.11686/cyxb20150213
http://cyxb.lzu.edu.cn
杨洁晶, 万娟娟, 娜丽克斯, 任爱天, 鲁为华. 28种植物种子形态学性状及其萌发对绵羊瘤胃消化的反应. 草业学报, 2015, 24(2): 104-115.
Yang J J, Wan J J, Narkes Wali, Ren A T, Lu W H. Seed morphology and effect of sheep rumen digestion on germinability of 28 grass of Tianshan. Acta Prataculturae Sinica, 2015, 24(2): 104-115.
猜你喜欢
小天使·一年级语数英综合(2019年10期)2019-11-10
儿童时代·幸福宝宝(2019年9期)2019-10-28
莫愁·家教与成才(2017年7期)2017-07-11
东方教育(2016年9期)2017-01-17
法制与社会(2016年33期)2016-12-15
新闻世界(2016年11期)2016-12-10
小学阅读指南·低年级版(2016年9期)2016-05-14
戏剧之家(2016年4期)2016-03-25
现代园艺(2016年2期)2016-03-15
